Article Text

Abstract
BACKGROUND Activatedc-fos binds tojun proteins to form the activation protein 1 (AP-1) transcription factor that regulates cytokine and other proinflammatory genes.c-Fos may play a key role in nasal polyp formation. Glucocorticoids may exert their anti-inflammatory effects through an interaction of glucocorticoid receptors withAP-1 that leads to mutual inactivation of both factors, and a “default” termination ofAP-1 mediated gene activation. This may explain the beneficial effects of glucocorticoids in the treatment of nasal polyps.
METHODS To test this hypothesis in humans in vivo the immunohistochemical expression ofc-fos-immunoreactive material (c-fos-irm) was assessed in nasal polyps from eight steroid naive subjects, polyps from eight subjects treated with topical beclomethasone dipropionate (BDP), and normal inferior turbinate nasal mucosa (n = 6).
RESULTS mRNA forc-fos was detected in all nasal polyps and normal mucosa. In contrast, c-fos-irm was present in all steroid naive subjects but in only two of the eight subjects treated with BDP (p = 0.007, two-tailed Fisher’s exact test).c-Fos-irm was expressed solely in epithelial cells and glandular structures; it was expressed in normal epithelium and glands, but the staining intensity was low.
CONCLUSION Glucocorticoids appear to modulate expression of c-fos-irm and possibly AP-1 in human airway epithelial cells in vivo.
- glucocorticoids
- c-fos
- AP-1
- epithelial cells
- nasal polyps
Statistics from Altmetric.com
Nasal polyps are known to express high levels of proinflammatory transcription factors such asc-fos.1 Thec-fos oncogene participates in transduction cascades that are initiated by many proinflammatory mediators.2 c-Fos combines with members of the c-jun family to form the transcription factor activation protein 1 (AP-1). AP-1moves from the cytoplasm into the nucleus where it binds toAP-1 promoter sites on a wide variety of genes. AP-1 helps initiate transcription of many proinflammatory genes, including those for cytokines.
One mechanism of glucocorticoid action is postulated to be an interaction with c-fos orAP-1.3-6 Glucocorticoids enter cells and bind to glucocorticoid receptors. These activated receptors are thought to bind to activated AP-1, leading to mutual inactivation of both protein factors. This reduction in the amount of free activated AP-1 may lead to a “default” reduction in the expression of genes that require AP-1 for activation.3-5 ,7 ,8 Hence, synthesis of mRNAs for many proinflammatory products may become reduced.9 ,10
A human model would be useful for studying this postulated interaction between glucocorticoid and c-fos and to learn its relevance to the actions of glucocorticoids in vivo. Since Emery et al 1 noted the presence of c-fos, c-jun, and c-myc mRNA in human nasal polyps it has been of interest to determine the distribution ofc-fos immunoreactive material (c-fos-irm) in nasal polyps and the effects of glucocorticoids on c-fos-irm and mRNA expression in vivo. It was hypothesised thatc-fos-irm would be expressed as a marker of cellular activation in polyps, but that glucocorticoid treatment would reduce the levels of expression. Nasal polyps from steroid treated and steroid naive subjects were examined by immunohistochemistry and reverse transcriptase-polymerase chain reaction (RT-PCR).
Methods
NASAL TISSUES
Subjects with refractory nasal obstruction due to nasal polyposis underwent polypectomy. Patient diagnoses, allergy status (determined by clinical and allergy skin test parameters) and medications were noted (table 1). Inferior turbinates were obtained from six subjects who underwent surgery for nasal septal deviation and jaw deformity. This tissue served as a source of normal nasal mucosa. Cases were coded to provide confidentiality. Tissues were immediately frozen in liquid nitrogen and stored at –70°C. A portion was frozen in OCT for cryostat sectioning.
Subject characterisation and results of c-fos immunohistochemical analysis
Immunohistochemical analysis and RT-PCR were performed to detectc-fos immunoreactive material (c-fos-irm) and mRNA, respectively. Clinical histories were reviewed and individual cases were sorted into two groups: those who had never received glucocorticoids or had received none in the past three months (steroid naive) and those who had received any topical nasal or oral steroid in the past three months (steroid treated).
IMMUNOHISTOCHEMISTRY
Cryostat sections (4 μm) were cut onto silanated glass slides and stored at –20°C. Slides were heated to 4°C for five minutes, then to room temperature before washing in phosphate buffered saline (PBS) twice for five minutes each. Endogenous peroxidase activity was destroyed in 0.03% hydrogen peroxide in methanol for 30 minutes at room temperature. The slides were rinsed gently with distilled water, then washed with PBS twice for five minutes each. Endogenous biotin was blocked (Dako Biotin Blocking System X590) by: (1) adding avidin solution (#1) for 10 minutes, (2) washing with PBS twice for five minutes each, (3) adding “Biotin Solution #2” for 10 minutes, and (4) washing with PBS twice for five minutes each. Non-specific binding sites were blocked with Dako blocking buffer for five minutes. Polyclonal rabbit antiserum specific for humanfos (Caltag, San Francisco, CA, USA) diluted 1:300 in 2% BSA was added, and slides incubated overnight at 4°C in a humidified chamber. Non-immune rabbit serum was used as a negative control.
Slides were washed with PBS three times for 10 minutes each, then biotinylated goat anti-rabbit IgG (Dako LSAB Kit, Peroxidase, K684) was added for 10 minutes. After washing in PBS twice for five minutes each, streptavidin-peroxidase was applied for 10 minutes. After washing twice with PBS for five minutes each, AEC chromogen (Dako) was applied for exactly 40 seconds, then immediately rinsed with distilled water. Slides were mounted with Crystal/Mount (Biomeda), air dried, and baked at 80°C for 10 minutes. After cooling, slides were coverslipped with Permount (Fisher).
Slides from steroid naive polyps known to have positivec-fos-irm staining and from steroid treated polyps known to have negative c-fos-irm staining were always included as positive and negative controls when new polyp sections were stained. The distribution ofc-fos-irm staining was observed by light microscopy, but was not scored for relative intensity because of concerns about adequate quantitative standards. Therefore,c-fos-irm staining was scored as either present or absent.
The proportions of c-fos-irm positive polyps in the two subject groups were compared by Fisher’s exact test with probability quoted for two tailed comparisons.
POLYP INCUBATION IN VITRO
Nasal polyps from steroid naive subjects were cut into 4 × 4 mm fragments and cultured in 2 ml CMRL-1066 (Biofluids, Rockville, MD, USA) with or without 1 μM dexamethasone in CMRL-1066 in 5% CO2, room air11 for 24 hours before being frozen in OCT in liquid nitrogen. Expression ofc-fos-irm was determined in fresh frozen, media-treated, and dexamethasone-treated polyps.
RNA EXTRACTION
Fresh or frozen nasal polyp tissues (4 × 4 mm) were placed in sterile Eppendorf tubes, 1 ml of Tri-Reagent (Molecular Research Corporation, Cincinnati, OH, USA) was added, and homogenised at room temperature.12 Chloroform (0.2 ml) was added and the two phases were separated by centrifugation at 12000g for 10 minutes. The top RNA-containing layer was removed to a fresh tube, 0.5 ml isopropanol was added, and the contents were gently mixed for 15 minutes. After centrifugation at 12000g for 30 minutes the pellet was washed with 70% ethanol and again spun at 12000g for 30 minutes. The RNA pellet was resuspended in 10 ml diethylpyrocarbonate treated water and used immediately for RT-PCR.
PRIMERS
β-Actin primers were purchased from Clontech (Palo Alto, CA, USA). The primers13 ,14 generated an RT-PCR product 661 bases long from mRNA, but 867 bases long from genomic DNA because of the presence of a short intron. β-actin mRNA was used as a positive control in all RT-PCR reactions to ensure that intact mRNA was present and that there was no DNA contamination.
c-Fos primers were synthesised by the Lombardi Cancer Center Core Laboratory.15 The sense primer (5′-TGC CAA CTT CAT TCC CAC GGT) coded for nucleotides 1212–1232 in exon 2. The antisense primer (5′-TAG TTG GTC TGT CTC CGC TTG) was complementary to the mRNA sequence which spanned the exon 3–4 splice site (Cntd 1968AA GCG GAG ACA GAC Cntd 2102). The RT-PCR product from mRNA was 347 bases long, and no product could be synthesized from genomic DNA.
ZO-1 mRNA, which codes for the protein that forms epithelial tight junctions, was used as a positive control to demonstrate the presence of epithelial cells in nasal polyp RNA preparations. This was necessary to ensure that mRNA from epithelial cells (the site of c-fos immunoreactive material) was present in polyp RNA preparations.ZO-1 sense (5′Cntd 10AT AGA ATA GAC TCC CCT GGntd 30) and antisense (5′Cntd 443TG CTG GCT TGT TTC TCT ACntd 423) primers16 ,17 were synthesised. Two alternatively splicedZO-1 mRNAs are present in epithelial cells and represent a full length mRNA coded by at least three exons and a shorter mRNA with the second putative exon (“motif a”, 240 nucleotides) deleted. The RT-PCR products from these mRNAs were 433 and 193 nucleotides long, respectively.
QUALITATIVE RT-PCR
RNA, reverse transcriptase/antisense primers, and Perkin Elmer RT-PCR reagents (Norwalk, CN, USA) were mixed according to manufacturer’s recommendations at 4°C in a PE thermocycler. Mineral oil (70 μl) was added and the temperature increased to 42°C for 60 minutes to anneal the RT/antisense primer to specific mRNA sequences. The solution was denatured at 99°C for five minutes and cooled to 70°C.
Perkin Elmer taq, buffer, Mg2+, and sense primers were preheated to 70°C and added to each reversely transcribed tube (“Hot-Start” conditions that reduce non-specific priming). To permit efficient annealing and extension for this first cycle of PCR, the temperature was adjusted to 57°C and maintained for five minutes. This was followed by extension for two minutes at 72°C. Annealing temperatures were calculated according to Meinkoth and Wahl.18 Tubes were cycled for a total of 45 cycles of 94°C for one minute, 57°C for one minute, and 72°C for two minutes. This number of cycles is required because of the low yield of RNA from polyp fragments.
PCR products and Clontech DNA standards were run in 0.5 × TBE on 2% agarose (FMC, Rockland, ME, USA) for 1.5 hours at 120 V. Gels were stained with ethidium bromide and the bands were visualised by UV fluorescence and photographed with Polaroid 667 film. Photographs of UV fluorescence patterns from representative gels were scanned (Aldus Photostyler 2.0 for Windows, Aldus, Seattle, WA, USA) and positive and negative images were compiled (Corel Draw 4.0, Corel Corp, Toronto, Ontario, Canada) and laser printed.
Results
IMMUNOHISTOCHEMISTRY
c-Fos-irm was present in nasal polyps of steroid naive patients (figs 1 and 2, table 1). Epithelial cells, particularly those cells with cytoplasm in the most superficial half of the epithelial thickness, were the predominant site of expression. There were very few glands in polyps; gland-like structures had the appearance of invaginated epithelial tubes lined with cuboidal epithelial cells. When present in steroid naive subjects, these “glandular” cells were invariably positive forc-fos-irm.

c-Fos-irm in nasal polyps. (A) c-Fos-irm in gland and duct cytoplasm in a nasal polyp from a subject never treated with steroids. (B) Non-immune serum control.

c-Fos-irm in nasal polyps. (A) Steroid naive subject with c-fos-irm in epithelial cell cytoplasm. (B) Non-immune serum control of a consecutive section. (C) BDP treated subject with no detectable c-fos-irm.
c-Fos-irm was absent in nine of 12 polyps from subjects who were currently taking oral or topical glucocorticoids. This ratio was significantly different from that for steroid naive subjects (p = 0.0014), suggesting that glucocorticoid use contributed to the difference in c-fos-irm expression (table 2). Treatment with topical beclomethasone dipropionate (BDP) for at least three months significantly reducedc-fos-irm expression (p = 0.007).
Comparisons of c-Fos-irm immunohistochemistry in nasal polyp epithelium from steroid naive and steroid treated subjects
Only 30% of the subjects on glucocorticoids hadc-fos-irm expression in nasal polyps (tables1 and 2). In general the staining was less intense than in the steroid naive subjects, but semiquantitative immunohistochemistry was not performed to assess differences. One subject (G) had three polypectomies (G1, G2, G3) with c-fos-irm detectable in G3 while on topical BDP alone. Subject I had severe chronic sinusitis complicating nasal polyposis but had received oral methylprednisolone for only seven days prior to surgery. More than seven days of oral treatment may be necessary to dampenc-fos-irm expression. Staining in this subject was weak. Weak staining was also found for subject M who was treated with topical BDP. c-Fos-irm was absent in 75% of the polyps from subjects treated with topical BDP.
In all of the inferior turbinate control tissuesc-fos-irm was present in the superficial cells of the epithelium and a fraction of the submucosal gland cells (fig 3). The pattern of staining of epithelial cells was similar to that in nasal polyps (fig 2) but the staining was less intense. Basal cells had no staining in some sections. Submucosal gland cells were the only cells with detectable staining in the lamina propria.
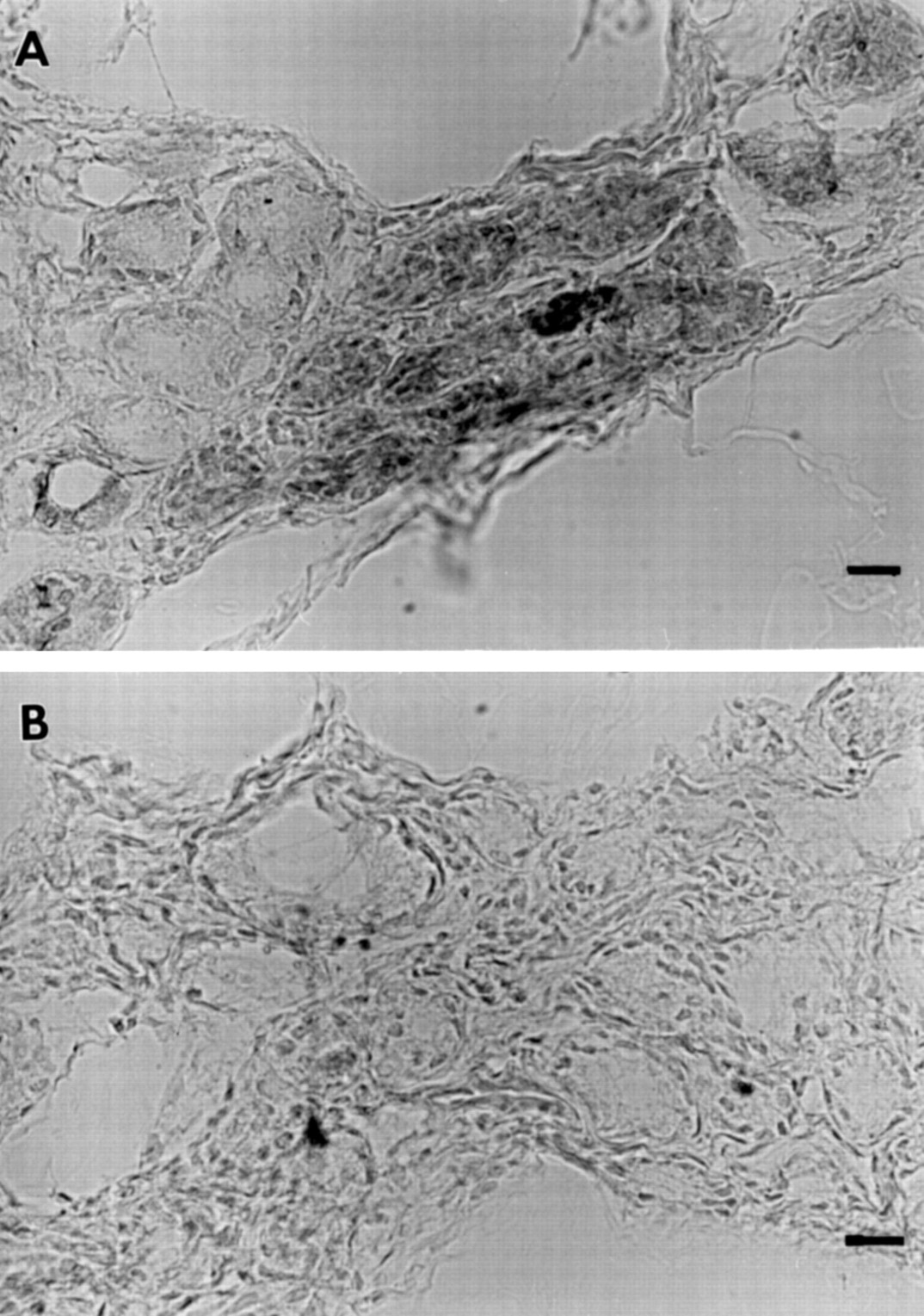
c-Fos-irm in human nasal submucosal glands. (A) Steroid naive subject with gland cells showing either intense or relatively weak staining. (B) Non-immune serum control with no staining. Bars represent 50 μm.
POLYP INCUBATION
Culture of polyps from steroid naive subjects for 24 hours in 1 μM dexamethasone did not reduce c-fos-irm expression. Longer periods of glucocorticoid treatment may be necessary to alter expression of c-fos-irm. However, culture conditions may activate cells, inducec-fos-irm expression, and confound these experiments.
QUALITATIVE RT-PCR
β-Actin mRNA was present, indicating extraction of intact RNA from nasal polyps of steroid naive (A, J, K, N, O) and steroid treated (B, C, G2, G3, L) subjects (fig 4). There was no DNA contamination since the single bands were at the size predicted for mRNA transcripts and not at the size predicted for genomic DNA.
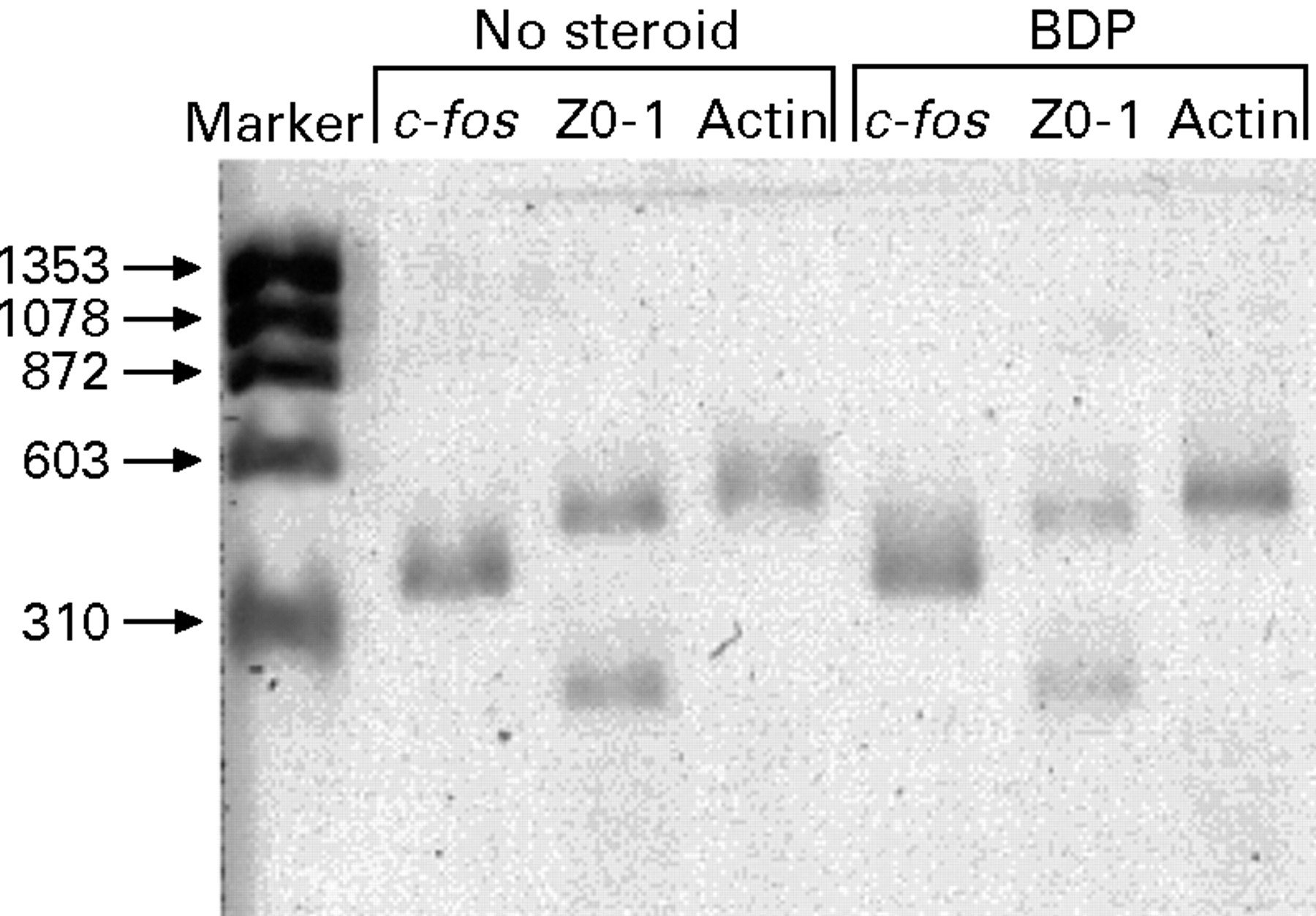
Qualitative RT-PCR of β-actin, ZO-1, and c-fos in human nasal polyp mRNAs from a steroid naive subject (subject J) and a subject treated with BDP (subject L).
ZO-1 mRNA was present in all the polyps, indicating the presence of epithelial cells. Therefore, it was possible for c-fos mRNA to be present since these were the cells that expressed c-fosimmunoreactive material. Both the 433 and 193 nucleotideZO-1 mRNAs were detected in all samples.
c-Fos mRNA was present in all the nasal polyps tested regardless of whether or not they had been treated with glucocorticoids. There did not appear to be a difference in relative expression when compared with β-actin orZO-1 band intensities.
mRNA for c-fos, ZO-1, and β-actin were all detected in inferior turbinate mRNA, indicating their expression in normal mucosal tissue (data not shown).
These data suggest that c-fos mRNA is actively and continuously transcribed or stored within epithelial cells in all nasal polyps. This is consistent with the high levels ofc-fos mRNA detected by RNase inhibition experiments by Emery et al.1Glucocorticoid treatment for 24 hours in vitro did not eliminate the steady state presence of c-fos mRNA in explants. In contrast, c-fos-irm was reduced by prolonged topical treatment in vivo.
Discussion
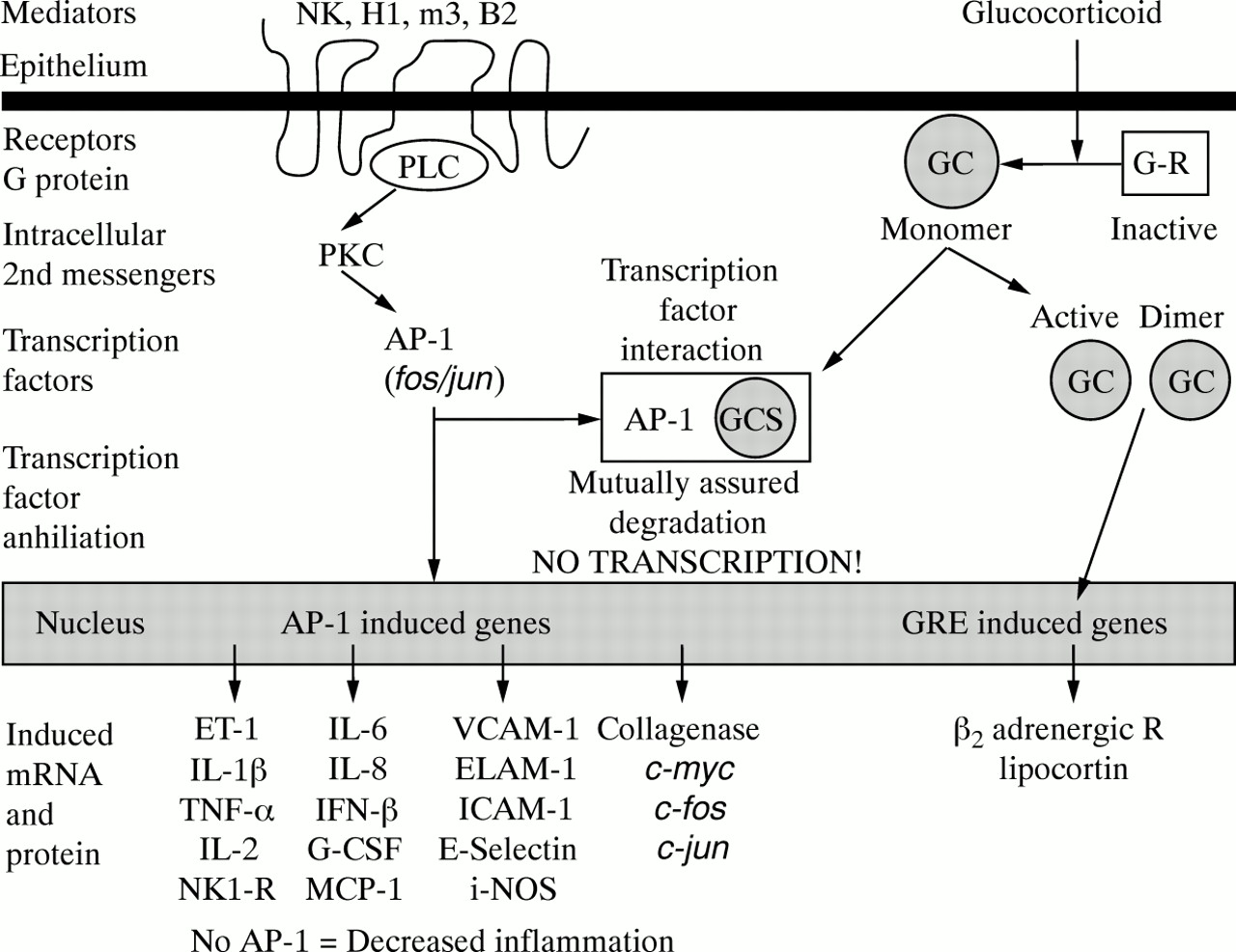
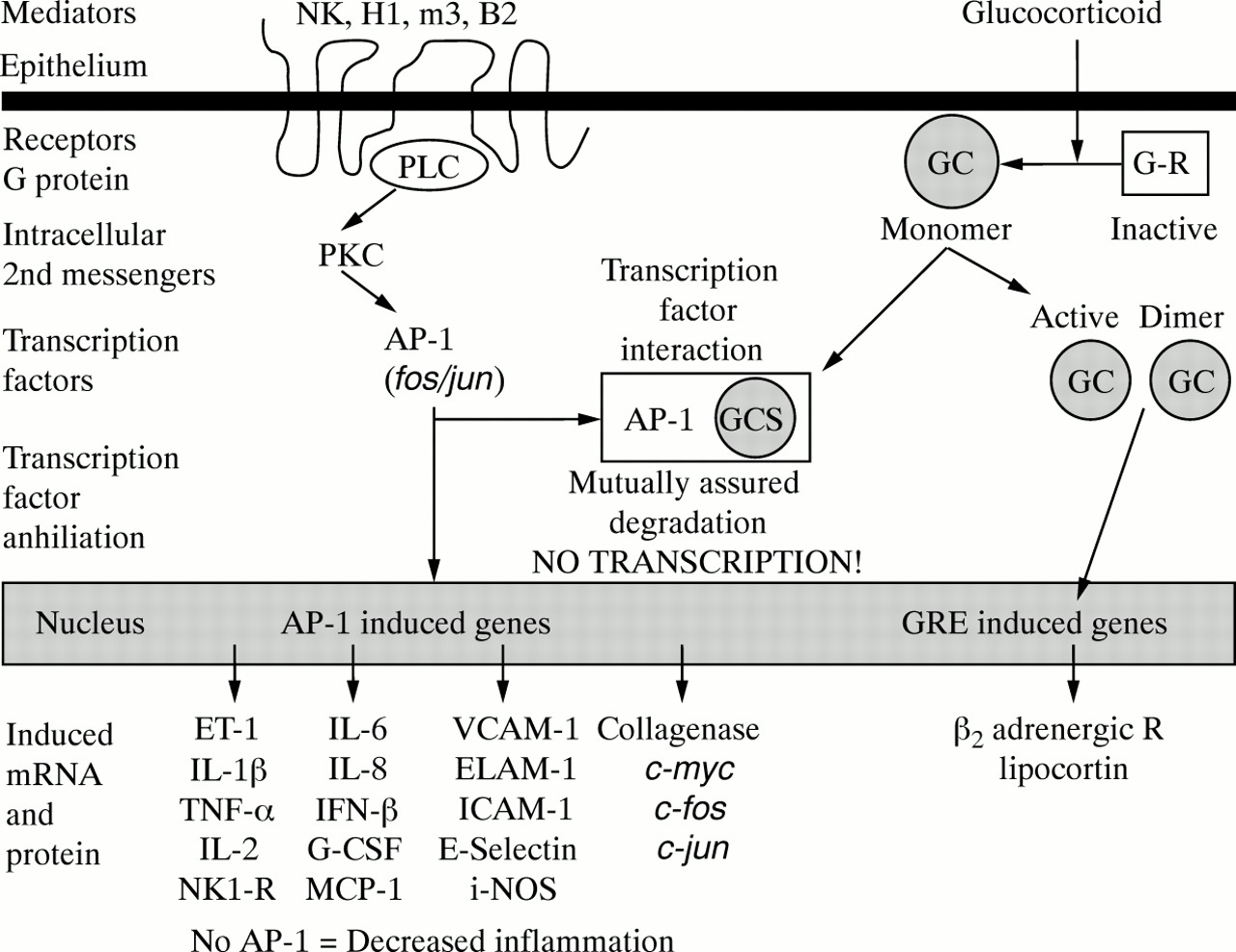
This experiment was undertaken to examine the hypothesis that glucocorticoids could modulate the expression ofc-fos/AP-1 in human tissues using nasal polyps as a clinically relevant human disease model. This hypothesis predicts that the activation of inflammatory mediators such as neurokinin (NK), histamine H1, muscarinic m3, bradykinin B2, and other 7-transmembrane receptors leads to activation of c-fos andc-jun, and the formation ofAP-1 (fig 5, left side).AP-1 participates in the induction of a wide variety of proinflammatory proteins, receptors, and mediators. Glucocorticoids bind to cytoplasmic receptors, which dimerise and translocate to the nucleus to induce genes such as the β 2 adrenergic receptor (fig 5, right side).AP-1 and activated glucocorticoid receptors can bind to each other, leading to their mutual proteolytic degradation and the loss of their nuclear transcription factor effects. The current study supports this hypothesis since nasal steroids reduced the expression of epithelialc-fos-irm.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hypothesis of c-Fos/AP-1 interactions with glucocorticoids. Activation of neurokinin (NK), histamine H1, muscarinic m3, bradykinin B2, and other 7-transmembrane receptors leads to the activation of c-fos and c-jun, and the formation of AP-1 (left side). AP-1 participates in the induction of a wide variety of proinflammatory proteins, receptors, and mediators. Glucocorticoids bind to cytoplasmic receptors which dimerise and translocate to the nucleus to induce genes such as the β2 adrenergic receptor (right side). AP-1 and activated glucocorticoid receptors can bind to each other, leading to their mutual proteolytic degradation and the loss of their nuclear transcription factor effects. The current study supports this hypothesis since nasal steroids reduced the expression of epithelial c-fos-irm.
The findings of this study indicate thatc-fos mRNA was transcribed in nasal polyps from all subjects and in normal nasal mucosa.c-fos mRNA was translated intoc-fos protein (c-fos-irm). In nasal polyps from steroid naive subjects epithelial and glandular cells stained intensely withc-fos-irm. In contrast, after steroid treatment in vivo staining was absent or was of low intensity. Higher intensity staining occurred in subjects with more extensive nasal and sinus disease, or in those who had been treated for shorter periods of time—for example, seven days of methylprednisolone in subject I. Twelve weeks of topical BDP was effective in reducingc-fos-irm expression, confirming the rationale for direct topical treatment of respiratory diseases.
The continuing presence of c-fos mRNA with reduced expression of c-fos-irm expression after steroid treatment could be due to (a) continued transcription ofc-fos mRNA but with reduced translation intoc-fos-irm protein, (b) increased turnover ofc-fos-irm protein such that levels were not detectable by immunohistochemistry, or (c) binding of activated glucocorticoid receptor monomers andc-fos-irm/AP-1leading to the mutual inactivation, proteolytic degradation, and the loss of both the c-fos-irm and glucocorticoid receptors. The latter is most consistent with current hypotheses of glucocorticoid actions (fig 5). However, the qualitative amplification protocol was not designed to detect differences between sets, so partial reductions in mRNA expression could have occurred and not been detected. Much more c-fos mRNA is detected by RNase protection assay in polyps than in other human nasal mucosa tissues.1
The epithelial localisation suggests that theAP-1 transcription factor plays an important role in epithelial cells and may significantly contribute to the pathology of nasal polyposis. AP-1 induces many mRNAs for proinflammatory proteins, including cytokines, that could initiate or promulgate mucosal inflammation.6 ,8-10 ,19 ,20 Alternatively, epithelial cells could be “innocent bystanders” that are activated by mast cell or eosinophil factors: c-fos expression would be a marker of cell activation. However, the production of eotaxin, RANTES, GM-CSF, IL-8 and other cytokines by epithelial cells suggests a more integral role.19 ,20
There was a striking absence of c-fos-irm expression in fibroblasts, endothelium, and inflammatory cells. This suggests thatc-fos/AP-1mediated events may be more important in the epithelium than these other cell populations, or that epithelial cells maintain a large “reservoir” of non-activated but immunoreactivec-fos that could be available for immediate cellular activation. Alternatively,c-fos-irm could be present and active, yet still be below the limits of immunohistochemical detection.
Low intensity c-fos-irm staining was also present in inferior turbinate mucosa epithelium and gland cells. Demolyet al have described similar low intensity staining in bronchial epithelium.21
Some of the subjects had recurrences of their nasal polyps despite treatment with topical glucocorticoids. This indicates that, although glucocorticoids may interfere with AP-1 and reduce polyp growth, there are still subjects who do not respond and require polypectomy. This implies the presence of steroid non-responsive mechanisms that contribute to this mucosal inflammatory state. This hypothesis is supported by Norlander et al who found that treatment with nasal budesonide spray (200 or 400 μg twice per day, Rhinocort Aqua, Astra, Lund, Sweden) for eight weeks did not alter the expression of mRNA for glucocorticoid receptor, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-10, TNF-α, or interferon-γ in polyps from patients with refractory nasal polyposis.22Other cytokines such as IL-13 may be responsible since they can induce steroid resistance.23
As already mentioned, there are several limitations to this study that can be rectified in future investigations. A placebo controlled comparison showing the expression of c-fosmRNA, c-fos-irm, and protein function after treatment with nasal steroids is warranted. However, the dose and duration of treatment remains to be established. Polyps may disappear with treatment, leaving no tissue for post-treatment analysis. Responses in aspirin sensitive and insensitive polyp patients should be compared. These in vivo studies suggest that the dramatic modulatory effects of glucocorticoids that can be readily demonstrated by reductionist in vitro experiments of purified or cultured lymphocytes, epithelial, or other cells may not predict mucosal effects in complex disease states. The qualitative RT-PCR confirmed the presence of epithelial cell mRNA (ZO-1) and the expression of c-fos mRNA in the epithelium, but this method should not be considered quantitative. Studies of the transcriptional and translational controlc-fos expression,c-fos-glucocorticoid receptor interactions, and c-fos turnover need to be established in model systems since there is insufficient protein and mRNA in polyps to develop these methods de novo. These experiments provide human in vivo evidence to support the hypothesis that activated glucocorticoid receptors can interact with c-fos and theAP-1 transcription factor and suggest that this interaction may contribute to the beneficial effects of glucocorticoids.
Acknowledgments
This study was supported by a research grant from Glaxo-Wellcome Research, Research Triangle Park, North Carolina, USA.The authors would like to give special thanks to Mark Osterman for his assistance.