Article Text

Abstract
BACKGROUND Aspergillus fumigatus can colonise the airways and the lungs with localised underlying conditions and occasionally invade the surrounding lung tissues even in subjects without systemic predisposing factors, presumably by escaping the local host defences. The aim of this study was to investigate the effects of A fumigatus culture filtrate (ACF) on the activities of human phagocytes—inhibition of germination of A fumigatus spores by alveolar macrophages (AMs) and hyphal damage by polymorphonuclear leucocytes (PMNs)—which are the critical host defences against A fumigatus.
METHODS Spores were incubated with AMs at a ratio of 1:1 in a medium containing different concentrations of ACF for 10 hours at 37°C. Spore germination was visualised with light microscopy and the inhibition rate was calculated. The percentage of hyphal damage caused by PMNs pretreated with various concentrations of ACF was measured by a colorimetric tetrazolium metabolic assay.
RESULTS The inhibition rate of spore germination by AMs cultured with medium alone (control) was 90 (0.8)% whereas that by AMs cultured with the medium containing 10% ACF was significantly (p<0.05) reduced to 41.7 (4.6)%. ACF suppressed the inhibition of spore germination in a dose dependent manner without altering the phagocytosing activity against the spores. The percentage of hyphal damage caused by PMNs pretreated with medium-199 (control) was 78.1 (2.3)% compared with 65.3 (2.8)% when PMNs were pretreated with 50% ACF (p<0.05).
CONCLUSIONS A fumigatusreleases biologically active substance(s) which suppress the inhibition of spore germination by AMs and also suppress PMN mediated hyphal damage, and thus may contribute to the pathogenicity of this fungus.
- Aspergillus fumigatus
- human phagocytes
- host defence
Statistics from Altmetric.com
Aspergillus species, most commonly Aspergillus fumigatus, are responsible for a wide range of diseases including saprophytic colonisation, chronic necrotising aspergillosis, and rapidly invasive diseases.1 ,2 Although acute progressive and invasive diseases occur most commonly in systemic immunocompromised hosts, A fumigatus frequently colonises damaged lungs and airways including healed tuberculosis, bronchiectasis and cystic fibrosis, and occasionally invades the surrounding lung tissues even in patients without apparently compromised systemic host defences.1 ,2 However, the pathogenic mechanisms and substances are still poorly understood.
It has been proposed that host defences against A fumigatusconsist of mucociliary clearance in the airways, alveolar macrophages (AMs), and polymorphonuclear leucocytes (PMNs). AMs can ingest the inhaled spores of A fumigatus, inhibit them from germination, and kill them. The germinated spores and hyphae that escape the mucociliary clearance and AMs can be killed by PMNs, mainly by an oxidative mechanism.3 ,4 These lines of defence cooperate in elimination of the fungus.
We have previously reported that A fumigatus culture filtrate (ACF) causes damage to respiratory ciliated epithelium by reducing ciliary beat frequency, and suppresses human phagocyte function including PMN migration towards formyl-methionyl-leucyl-phenylalanine and O2 – release from PMNs in response to phorbol myristate acetate, and concluded that A fumigatus produces several substances which suppress phagocyte function and mucociliary clearance.5-7
In the present study we have investigated the effects of ACF on inhibition of spore germination by human AMs and hyphal damage by human PMNs, in addition to the effects of ACF on ingestion of spores by AMs and the release of O2 – from PMNs in response to A fumigatus hyphae.
Methods
FUNGUS
A fumigatus, YN strain, clinically isolated from the sputum of a patient with invasive pulmonary aspergillosis, was used in the present study. The strain was stored on potato dextrose agar (PDA) (Difco Laboratories, Detroit, Michigan, USA) slants at –80°C. Spores were obtained by scraping the surface of the slants, suspended in phosphate buffered saline (PBS), and kept at 4°C. To induce spore germination a suspension of 1.0 × 106 spores in RPMI-1640 (Gibco BRL, Paisley, UK) was incubated in polypropylene tubes (Falcon, Becton Dickinson, Lincoln, New Jersey, USA) for 16 hours at 30°C and for an additional two hours at 37°C.8 After incubation for 18 hours almost all spores developed into hyphae with a length of 10–30 μm, but not branched. The hyphae were washed with PBS, vigorously vortexed, and used in the PMN experiments.
PREPARATION OF A FUMIGATUS CULTURE FILTRATE
A fumigatus culture filtrate (ACF) was prepared as described previously.5 ,7 Spores (2 × 107) of YN strain were cultured in flasks containing 300 ml of medium-199 (Gibco BRL) at 37°C. After five days of culture the fungi were removed by passing the medium through gauze, and ACF was sterilised by passage through Millex-GV 0.22 μm millipore filters (Millipore Products, Bedford, Massachusetts, USA) and stored at –80°C.
PHAGOCYTES
Human AMs were isolated from bronchoalveolar lavage (BAL) fluid of six healthy adult volunteers. BAL was performed with 4 × 50 ml sequential volumes of saline at 37°C. The recovery rate of BAL fluid was 60–65%, and Giemsa staining of cytospin preparations of BAL suspensions showed that more than 90% comprised AMs.
PMNs were prepared from the heparinised venous blood obtained from eight healthy adult volunteers by dextran sedimentation followed by centrifugation over Ficoll gradient and hypotonic lysis of erythrocytes. After treatment with ACF, the viability of cells was more than 95% as assessed by the trypan blue exclusion test. Cell damage was checked by release of lactate dehydrogenase (LDH) and no difference was found in LDH concentration whether or not they had been treated with ACF.
INGESTION OF SPORES BY AM
Ingestion of spores was assessed as described previously.7 Spores of YN strain were added to the AM suspensions (2 × 106/ml) in medium-199 containing ACF (0%, 5%, and 10%) at an AM:spore ratio of 1:5. Each mixture was incubated in a water bath with shaking at 37°C for 120 minutes. Immediately after incubation, all tubes were washed with cold 0.1% EDTA-PBS. Cells were attached to a glass slide by the cytospin procedure and stained with May-Grunwald-Giemsa. Both percentage phagocytosis and phagocytic index were calculated by counting 500 AMs per glass slide under a light microscope. Percentage phagocytosis was defined as the percentage of AMs with one or more spores ingested or attached. The phagocytic index was the mean number of spores that had been ingested by or attached to100 AMs.
INHIBITION OF SPORE GERMINATION BY AM
The ability of AMs to inhibit spore germination was determined by a modification of the procedure of Waldorf et al.9 One millilitre aliquots of cell suspension in culture medium (RPMI-1640 with 5% heat inactivated AB serum) were seeded into multiwell tissue culture plates (Falcon) and incubated at 37°C in a 5% CO2 incubator for two hours. Non-adherent cells were discarded by washing with RPMI-1640 and the culture medium containing various concentrations of ACF (0%, 1%, 3%, 5%, and 10%) was added to each well to form monolayers of AMs at a concentration of 5 × 105/well. AMs were challenged with spores in a ratio of 1:1 and centrifuged for five minutes at 100g to facilitate contact between the spores and AMs. Incubation was continued for a total of 10 hours at 37°C in the CO2 incubator, after which the plates were centrifuged, the supernatant discarded, and 0.5 ml of 2.5% deoxycholate (Sigma, St Louis, Missouri, USA) added to each well to lyse AMs. The plates were stained with lactophenol cotton blue and an inverted microscope (Olympus Optical Co, Tokyo, Japan) was used to visualise spore germination. The inhibition rate was calculated from the percentage of spores which did not germinate out of a total of 500 examined per well.
RELEASE OF O2– FROM PMNS IN RESPONSE TO HYPHAE
Release of O2 – from PMNs in response toA fumigatus hyphae was assessed by superoxide dismutase (SOD) inhibitable reduction of cytochrome C.10 Briefly, PMNs were preincubated with medium-199 containing various concentrations of ACF (0%, 10%, 30%, and 50%) for one hour at 37°C and then washed with PBS. PMNs were incubated with hyphae at a ratio of 1:1 in 1 ml of Hank’s balanced salt solution (HBSS) containing cytochrome C (120 μmol) (Sigma), with or without 50 μg SOD (Sigma), in a shaking water bath at 37°C for one hour. These reaction tubes were centrifuged and O2 –production was assessed in the supernatants as the difference in absorption from the control at 550 nm measured on a spectrophotometer (Hitachi U-3210, Hitachi Ltd., Tokyo, Japan), and amount of O2 – in nanomoles released per 106PMNs was calculated. All experiments were performed in quadruplicate.
PMN-INDUCED HYPHAL DAMAGE
Damage of the hyphae caused by PMNs was assessed by a colorimetric assay.8 PMNs were pretreated with various concentrations of ACF (0%, 10%, 30%, and 50%) for one hour and washed with PBS. PMNs (6 × 106) were resuspended in HBSS and added to hyphae in a final volume of 1 ml containing 5% heat inactivated AB serum at a PMN:hyphae ratio of 4:1. After one hour of incubation at 37°C in a CO2 incubator the PMNs were lysed with distilled water. The hyphae were then washed with distilled water and resuspended in 0.4 ml of 0.5 mg/ml 2,3-bis-(2-methoxy-4-nitro-5-sulphenyl)-(2H)-tetrazolium-5-carboxanilide (XTT; Sigma) supplemented with 40 μg/ml coenzyme Q (Sigma). Assay tubes were further incubated for one hour at 37°C in a water bath and 100 μl from each supernatant was transferred to the wells of microtitre plates. Absorption at 450 nm was determined using an ELISA plate reader (Flow Laboratories). The percentage of hyphal damage was defined as [1—(A450 of fungi incubated with PMN – A450 of PMN alone)/A450 of fungi incubated without PMN] × 100%.
ANALYSIS OF DATA
All data are presented as mean (SE) values. AM experiments were carried out in duplicate and PMN experiments in quadruplicate on four or five separate occasions. The raw data were subjected to repeated measures analysis of variance (ANOVA) and the Dunnett’s pairwise multiple comparison t test. A p value of less than 0.05 was considered to be significant between control and ACF treatment.
Results
EFFECT OF ACF ON INHIBITION OF SPORE GERMINATION BY AM
AM cultured with medium alone (control) inhibited germination ofA fumigatus spores effectively with an inhibition rate of 90 (0.8)%. On the other hand, treatment with 5% and 10% ACF led to reductions of 29% and 54%, respectively, in the inhibition rate compared with the control (p<0.05; fig 1). The number of AMs recovered from culture supernatant in each well after 10 hours of incubation was always less than 4 × 104 which indicated that the suppressive effect of ACF was not a consequence of loss of AMs. To determine whether ACF has a direct effect on spore germination, spores were incubated in AM-free medium containing ACF at concentrations similar to those used in the AM experiments. ACF did not show any direct effects on spore germination, and germination rates incubated with or without ACF were always >99%.
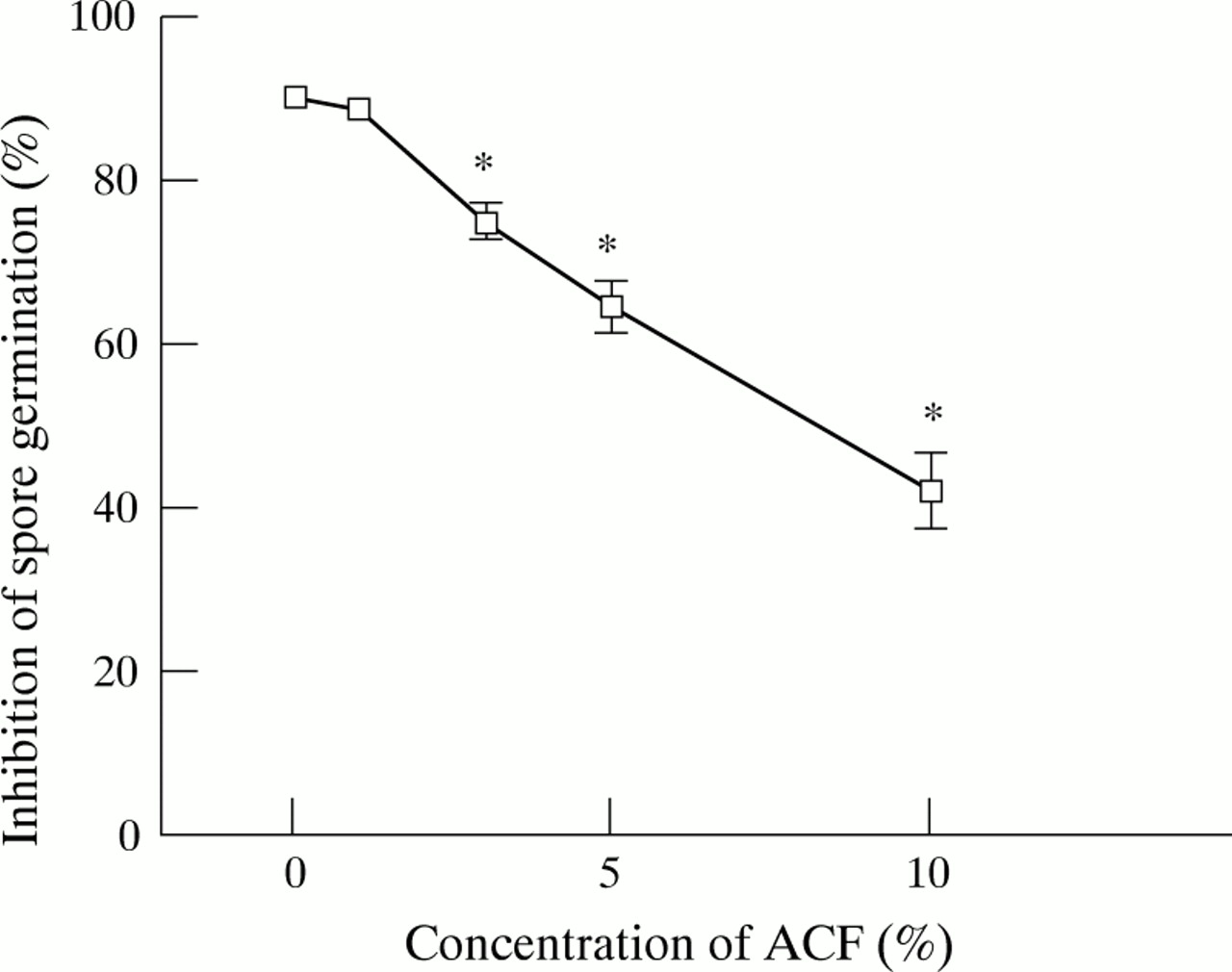
Effect of A fumigatus culture filtrate (ACF) on inhibition of spore germination by human alveolar macrophages (AMs). AMs co-cultured with different concentrations of ACF were challenged with spores at 37°C for 10 hours. The AM:spore ratio was 1:1. The percentage of inhibition was determined by calculating the number of spores which did not germinate. The results represent the mean (SE) of four separate experiments, each performed in duplicate. *p<0.05 compared with control (medium alone).
The effect of ACF on the phagocytic activity of AMs against spores was also evaluated. Percentage phagocytosis and the phagocytic index were 69.1 (7.0)% and 155.6 (45.8) in medium alone, 73.6 (5.0)% and 179.5 (54.1) when pretreated with 5% ACF, and 72.0 (4.4)% and 172.8 (52.1) when pretreated with 10% ACF. Thus, 5–10% ACF had no significant effect on either AM mediated percentage phagocytosis or percentage phagocytic index (p>0.05). It is therefore very likely that the suppressive effect of ACF on the inhibition of spore germination described above was not associated with its effect on the ingestion of spores.
EFFECT OF ACF ON O2– RELEASE FROM PMN STIMULATED WITH HYPHAE
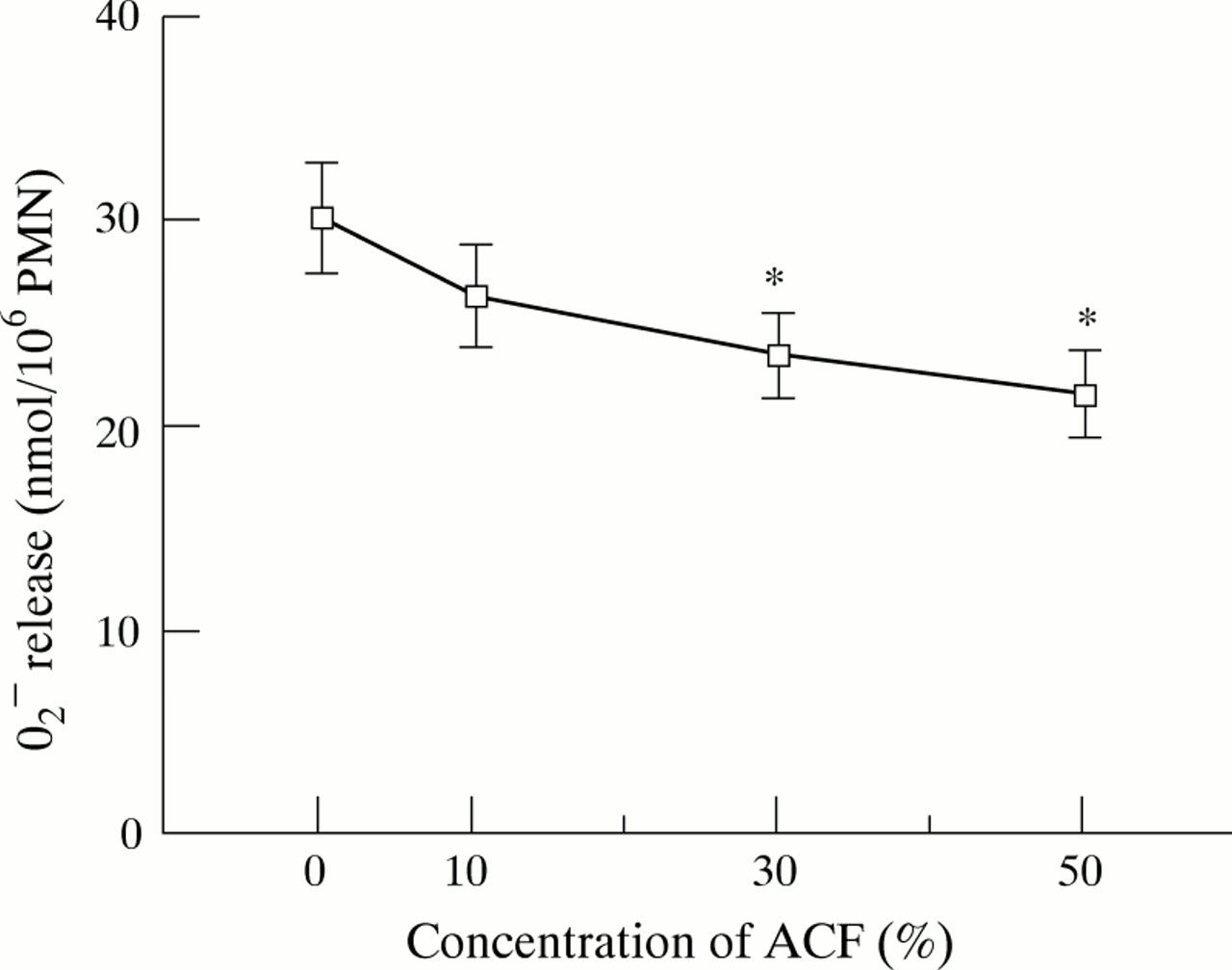
The effect of ACF on the oxidative metabolic burst of PMN, measured as the release of O2 – in response to non-opsonised A fumigatus hyphae, was examined. As shown in fig 2, pretreatment with 30% and 50% ACF for one hour significantly suppressed O2 – release in response to hyphae by 22% and 29%, respectively (p<0.05). Since spontaneous O2 – release was not affected by ACF, and ACF itself did not stimulate O2 –release of PMNs,7 this suppressive effect of ACF was not due to exhaustion of PMNs by ACF.
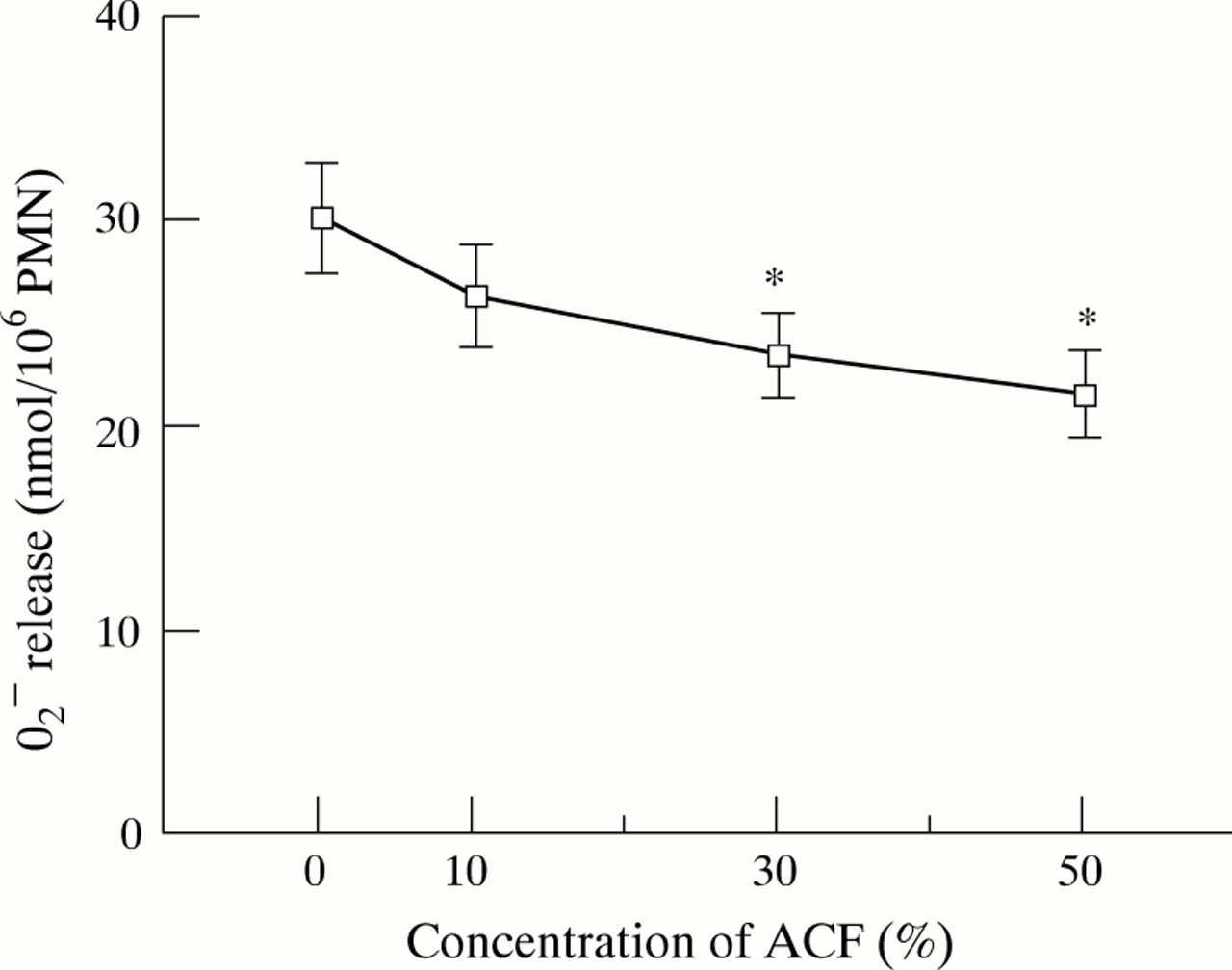
Effect of A fumigatus culture filtrate (ACF) on O2 – release by human polymorphonuclear leucocytes (PMNs) in response to A fumigatus hyphae. PMNs were pretreated with various concentrations of ACF at 37°C for one hour. O2 – release was assessed by cytochrome C reduction assay at a PMN:hyphae ratio of 1:1 and expressed as nmol/106 PMNs/h. The results represent the mean (SE) of five separate experiments, each performed in quadruplicate. *p<0.05 compared with control (medium-199).
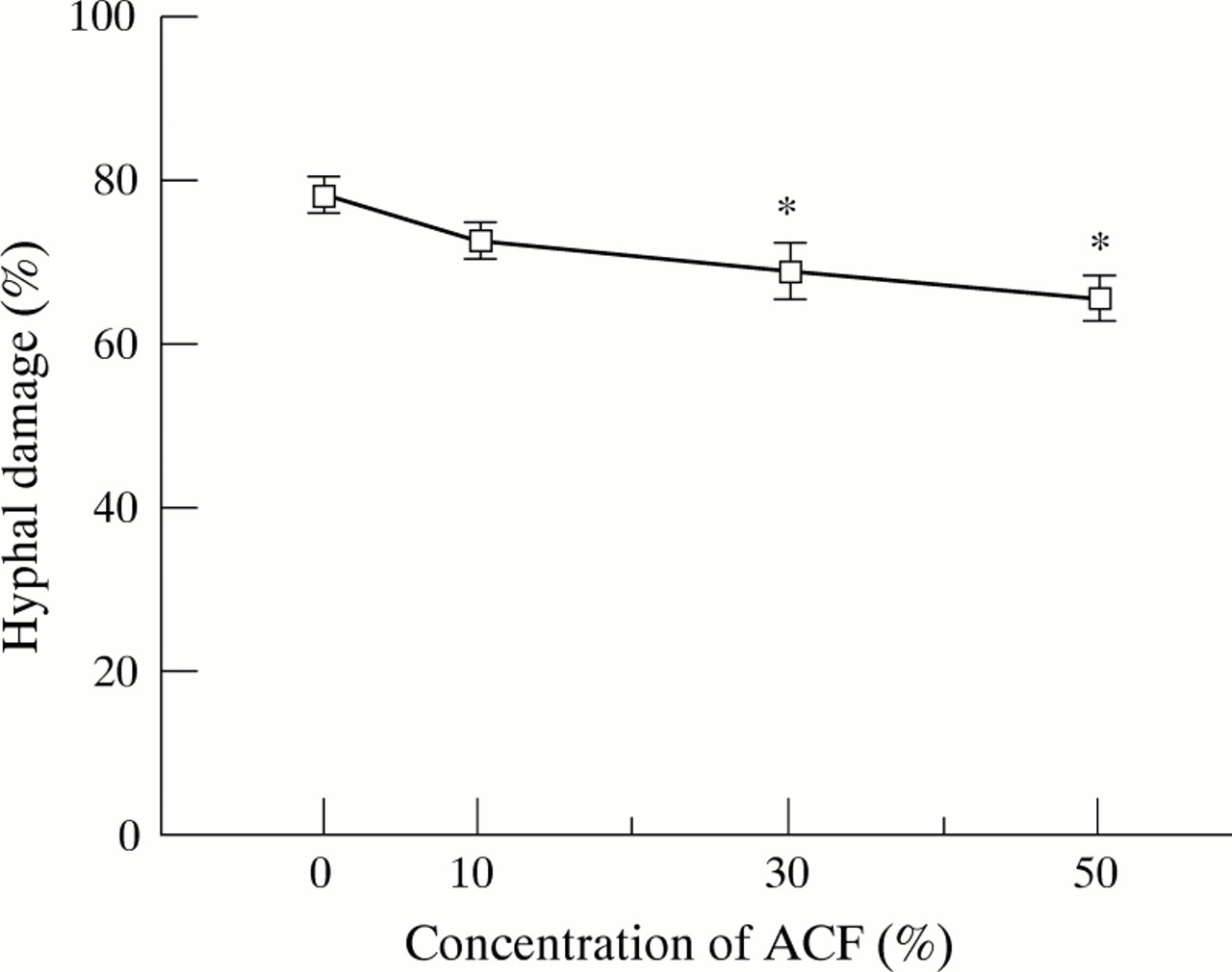
EFFECT OF ACF ON PMN INDUCED HYPHAL DAMAGE
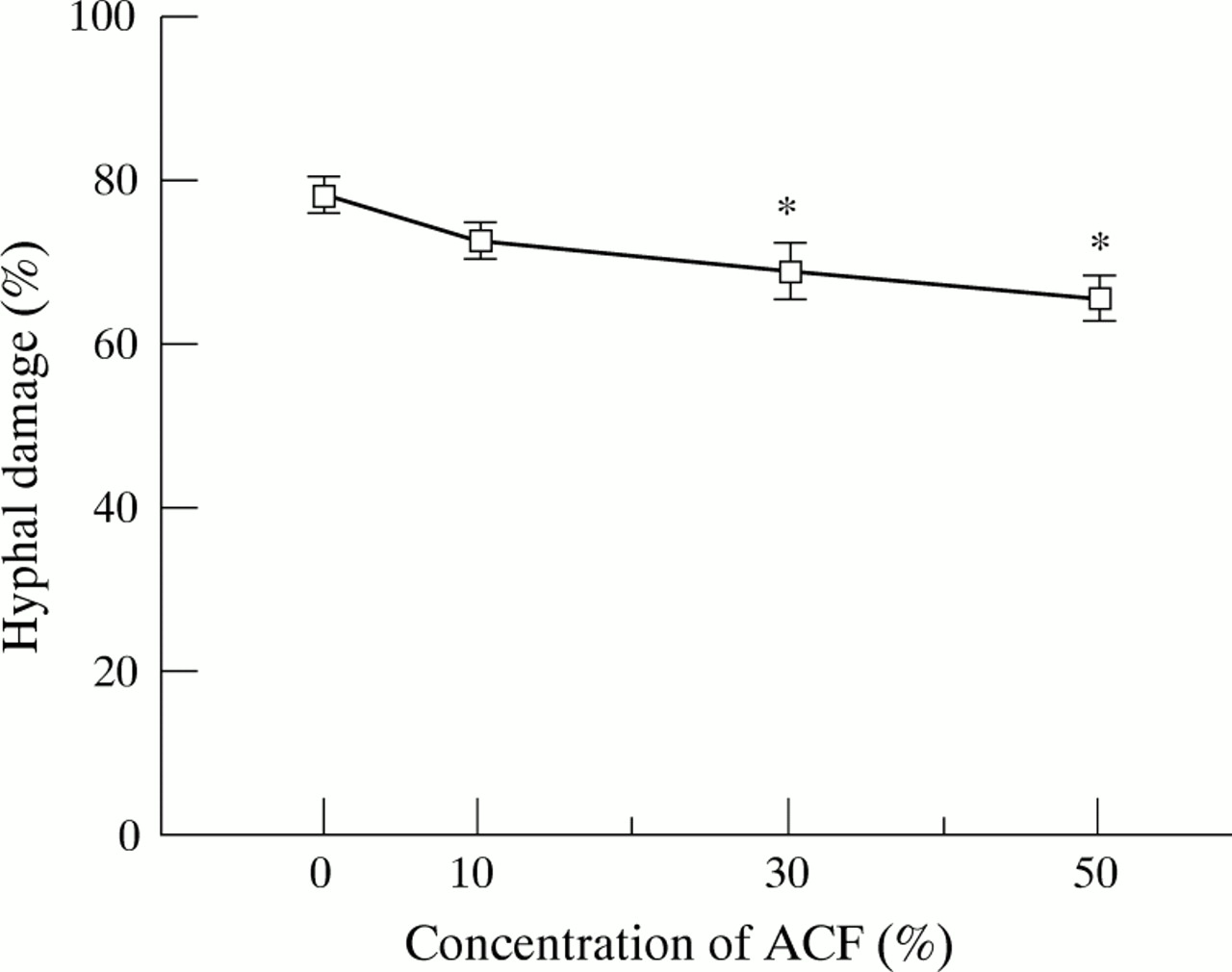
We assessed the hyphal damage using the XTT colorimetric metabolic assay, a rapid and quantitative method for assessment of fungal cell damage.8 The effects of ACF on PMN induced damage ofA fumigatus hyphae are shown in fig 3. Pretreatment with 30% and 50% ACF for one hour significantly suppressed the percentage of hyphal damage by 12% and 16%, respectively (p<0.05). When PMNs and hyphae were incubated with ACF throughout the one hour experimental period without pretreatment, ACF showed the same suppressive effect on the PMN induced hyphal damage (data not shown). When hyphae were incubated in PMN-free medium containing ACF at the same concentrations as used in the PMN experiments, ACF did not affect the metabolic activity of the hyphae as determined by XTT assays (data not shown).
{kind=link}
{kind=link}
{kind=link}
Effects of A fumigatus culture filtrate (ACF) on hyphal damage by human PMNs. PMN suspensions pretreated with various concentrations of ACF were co-cultured with A fumigatus hyphae at 37°C for one hour. The PMN:hyphae ratio used in this experiment was 4:1. The percentage of hyphal damage was examined by a colorimetric assay using (2,3)-bis-(2-methoxy-4- nitro-5-sulphenyl)-(2H)-tetrazolium-5-carboxanilide (XTT). The results represent the mean (SE) of five separate experiments, each performed in quadruplicate. *p<0.05 compared with control (medium-199).
Discussion
The results of this study show that ACF suppresses inhibition of spore germination of A fumigatus by human AMs without altering their phagocytosing activity against the spores, and that ACF also suppresses human PMN induced A fumigatus hyphal damage apparently parallel with O2 – release in response to hyphae. Although the present study was performed using a single strain of A fumigatus, limited experiments carried out with other clinical isolates of A fumigatus have shown similar results (data not shown).
Roilides et al reported that PMNs can respond to and damage Aspergillus hyphae irrespective of opsonisation, and that serum independent receptors of PMNs may contribute to attachment to Aspergillus hyphae and stimulation of the oxidative burst in the alveoli where serum opsonisation is poor.10 Warren et al found that PMNs can damage hyphae by releasing reactive oxygen intermediates and that the degree of the damage correlates with the duration and the levels of oxidant production.1 In this study we found that ACF suppresses O2 – release from PMNs in response to non-opsonised hyphae and also suppresses PMN mediated hyphal damage at similar concentrations. We assume that A fumigatusderived products circumvent the host defence network, thus enhancing the chance of colonisation, proliferation, and invasion of A fumigatus in human tissues.
It has been suggested that secondary metabolites, especially gliotoxin and extracellular proteases, may contribute to the pathogenic potential of A fumigatus. 6 ,11-13 Müllbacher et al reported that a three day culture of A fumigatusgenerated metabolites with antiphagocytic activity (inhibition of phagocytosis by mouse peritoneal exudate cells, inhibition of adherence to plastic surfaces by rodent peritoneal macrophages and AMs, reduction of the basal rate of H2O2 production by human PMNs), and they identified one of the biologically active components as gliotoxin.11 ,12 Slight et al reported thatA fumigatus spore diffusates obtained from three hour culture supernatants reduced phagocytic activity (inhibition of oxidative burst by rat AMs and inhibition of production of tumour necrosis factor α by rat AMs). They suggested that the active component of spore diffusate was of small molecular weight (<10 kD) and might have a carbohydrate component.14 ,15 However, to our knowledge there have been very few studies to date that have reported the suppressive effects of Aspergillus products on the antifungal activities of human phagocytes. This study may be the first to demonstrate the suppressive effects of Aspergillusproducts on antifungal host defences by both human AMs and PMNs.
Amitani et al isolated the factors responsible for causing damage to the human respiratory ciliated epithelium from ACF and identified one of the most potent substances as gliotoxin.6 We have previously suggested that gliotoxin should be a candidate for the antiphagocytic substances in ACF.7 Richard et al detected gliotoxin in the lung tissue of turkey poults infected with A fumigatus and suggested the involvement of gliotoxin in the pathogenesis of aspergillosis.16 These findings suggest that gliotoxin should be one of the most important substances contributing to fungal growth in human tissues.
Additional studies are in progress to clarify which of various A fumigatus derived substances including gliotoxin are the most important in suppressing human phagocytic function.
Acknowledgments
We thank Professor R D Diamond, Section of Infectious Diseases, University Hospital, Boston University for his helpful advice regarding the assay of hyphal damage and Dr S Ishimura, Tsurumi University for his critical advice regarding the statistical analysis.