Article Text

Abstract
Background: Aberrant epithelial repair is a key event in the airway remodelling which characterises obliterative bronchiolitis (OB) in the transplanted lung. The potential for airway epithelium from lung transplant recipients to undergo epithelial to mesenchymal cell transition (EMT) was assessed in culture and in vivo in lung allograft tissue.
Methods: Change in epithelial and mesenchymal marker expression was assessed after stimulation with transforming growth factor β1 (TGF-β1) alone or in combination with tumour necrosis factor α (TNFα) and compared with untreated controls. The ability of cells to deposit extracellular matrix, secrete matrix metalloproteinases (MMPs) and invade collagen was investigated. Immunolocalisation of epithelial and mesenchymal markers was compared in airway tissue from stable recipients and those with OB.
Results: Untreated cells maintained epithelial morphology and phenotype. TGF-β1 reduced expression of epithelial markers, increased expression of vimentin and fibronectin, promoted collagen I and fibronectin deposition and increased MMP-9 production. Co-treatment with TNFα dramatically accentuated phenotypic and some functional features of EMT. Airway epithelial biopsies from recipients with OB demonstrated significantly increased staining for mesenchymal markers and significantly reduced E-cadherin staining compared with stable recipients.
Conclusions: These observations demonstrate the ability of human airway epithelium to undergo EMT and suggest this phenomenon may be a potential link between inflammatory injury and TGF-β1-driven airway remodelling in the development of OB.
Statistics from Altmetric.com
Lung transplantation provides a valuable therapeutic option for selected patients with end-stage lung disease.1 However, long-term survival remains limited to a median of 5 years by the development of bronchiolitis obliterans syndrome (BOS).2 The pathological lesion of BOS is obliterative bronchiolitis (OB), which is characterised by aberrant epithelial repair and airway remodelling leading to obstruction due to deposition of extracellular matrix (ECM) as a result of an excessive fibroblastic response.
The origin of the fibroblasts responsible for the deposition of ECM remains unclear, with in situ proliferation of resident fibroblasts and recruitment of circulating progenitor cells proposed as potential sources.3 Evidence from animal models of airway obliteration suggests epithelial injury and, importantly, a failure to re-establish an intact epithelium may be critical to the pathogenesis.4
Epithelial cells can respond to injury in a number of ways including repair, necrosis and apoptosis. However, epithelial to mesenchymal transition (EMT) is increasingly recognised as an alternative response to injury. During EMT, cells lose epithelial properties and gain properties of mesenchymal cells including production of matrix metalloproteinases (MMPs) and deposition of ECM.5 Recent data from animal models and epithelial cell lines suggest that EMT may be an important response in lung epithelium at the alveolar level,6 7 8 9 however data using primary human lung epithelium has been limited to one previous pilot study.10
Transforming growth factor β1 (TGF-β1) is implicated in driving fibrosis in the lung, liver and kidney, but a mechanism by which TGF-β1 may contribute to the pathogenesis of OB remains to be elucidated. Previous studies have shown that TGF-β1 expression is increased in recipients with OB11 12 13 14 and, in an animal model of airway obliteration, interrupting TGF-β1 binding to its receptor reduced intraluminal airway matrix deposition.15
TGF-β1 signals predominantly via the SMAD signalling pathway,16 17 18 but can also activate the mitogen-activated protein kinase (MAPK) pathway.19 20 21 However, activation of the MAPK signalling pathway by pro-inflammatory stimuli such as tumour necrosis factor α (TNFα)22 raises the possibility of cross-talk between TGF-β1 signalling and inflammatory signalling in epithelium. EMT may therefore provide a plausible link between excessive inflammation, TGF-β1 activity and the development of OB. We hypothesise that airway epithelial cells in the transplanted lung may undergo TGF-β1-driven EMT and that this may be exaggerated by pro-inflammatory stimuli such as TNFα.
We have previously demonstrated preliminary evidence of EMT in the airways of lung transplant recipients by showing increased expression of the single mesenchymal marker S100A4 in airway epithelium from stable lung allografts.10 This study aims to extend these initial observations by comprehensively assessing the ability of airway epithelial cells from lung transplant recipients to undergo EMT and evaluating the evidence for EMT in tissue sections from stable lung transplant recipients and those with OB.
Methods
Antibodies and reagents
See online supplement for details.
Cell culture
Primary bronchial epithelial cells (PBECs) from stable lung transplant recipients were isolated as previously described (see online supplement for details).23
Cell treatments
Cells were cultured in media alone or treated with TGF-β1 (0.1–20 ng/ml) and/or TNFα (1–20 ng/ml) for 72 h. Protein concentrations were determined using the BCA protein assay kit (Perbio Science, Northumberland UK).
Western blotting
Total cell lysates (5–60 μg) were separated on 4–12% bis-Tris gels (Invitrogen, Paisley, UK). Membranes were incubated with primary antibodies and detected with horseradish peroxidase (HRP)-labelled IgG conjugates (Abcam, Cambridge, UK). Antibody complexes were visualised using the SuperSignal West Pico chemiluminescent kit (Perbio Science). Results are normalised to β-tubulin.
Immunofluorescence
Cells fixed in 4% (w/v) paraformaldehyde or explanted lung tissue were incubated with primary antibodies and detected using an appropriate fluorochrome-linked secondary antibody. DAPI was used as a nuclear counterstain. Images acquired using a LSM 510 laser scanning confocal microscope.
Gelatin zymography
MMP-9 activity in conditioned media from cells were analysed as previously described (see online supplement for details).10
Invasion assay
The invasive potential of cells was evaluated by assessing penetration of Matrigel coated filters. Briefly, cells were treated with TGF-β1 (10 ng/ml) and/or TNFα (20 ng/ml) for 72 h, 10 000 cells loaded in the invasion chamber and incubated for 24 h at 37°C. The number of invasive cells was determined following Diff-Quik (Dade Behring, Milton Keynes, UK) staining and scored at bright field microscopy.
Trichloroacetic protein precipitation
Proteins secreted from the cells in to the culture media were assessed by trichloroacetic (TCA) protein precipitation. Briefly, 250 μl TCA (100% w/v) was added to 1 ml of culture media and incubated at 4°C for 10 min. The protein precipitate was then pelleted by centrifuging at 14 000 rpm for 5 min. The protein pellet was washed twice in 200 μl cold acetone and dried by heating to 95°C for 10 min. The pellet was resuspended and separated under denaturing conditions by SDS-PAGE. Membranes were incubated with primary antibodies and detected as previously described.
Human lung tissue sampling
Sequential sections from stable recipients, OB recipients and normal control lungs were stained with antibodies against E-cadherin, vimentin and α-smooth muscle actin (α-SMA) using a modified immunoperoxidase method (Envision; Dako, Ely, UK). The extent of the area of positive bronchial epithelial staining was assessed in five non-overlapping high power fields as previously reported (see online supplement for details).24
Statistical analysis
The response of cultured cells from each subject to TGF-β1, TNFα and TGF-β1 with TNFα were compared with their untreated control cells. Changes in protein expression and protein secretion (relative band density) and numbers of invasive cells were quantified relative to untreated controls and expressed as mean (standard error of the mean (SEM)).
The differences in EMT marker expression in control, stable and BOS affected individuals were quantified for each marker by averaging the percentage positive staining area in five high power fields of intact epithelium for each individual. Results are presented as mean (SEM).
The significance of differences between groups was assessed by a one-way analysis of variance (ANOVA) using SPSS 14.0. Differences with a p value of <0.05 were considered statistically significant.
Results
Characteristics of patient groups
Cultures of PBECs were obtained from five male lung transplant recipients with stable allograft function between 6 and 30 months after transplantation. All subjects showed no evidence of acute rejection (grade A2 or above by ISHLT classification) or infection and were BOS stage 0 (>90% baseline forced expiratory volume in 1 s, FEV1).
Transbronchial biopsies were obtained from six lung transplant recipients (five men, one woman) with stable allograft function between 6 and 12 months after transplantation. All subjects showed no evidence of acute rejection or infection and were BOS stage 0. Biopsies of OB affected lung were obtained from eight lung transplant recipients (three men, five women) undergoing retransplantation for advanced BOS stage 3 (<40% baseline FEV1) between 32 and 116 months after their first transplant. Normal control lung tissue was obtained from six unused donor lungs.
Effect of TGF-β1 and TNFα on epithelial cell morphology and protein expression
Untreated or TNFα treated cells maintained a uniform “cobblestone” appearance characteristic of epithelial cells (fig 1A(i) and (iii), respectively). The cells demonstrated high expression of the epithelial marker cytokeratin 19 (fig 1A(v) and (vii)) and little or no expression of the mesenchymal markers fibronectin (fig 1A(ix) and (xi)) or vimentin (fig 1A(xiii) and (xv)). TGF-β1 treatment markedly changed the cell phenotype to a more elongated bipolar phenotype characteristic of mesenchymal cells (fig 1A(ii)). This morphological change was associated with a downregulation in cytokeratin-19 expression (fig 1A(vi)) and an upregulation in fibronectin (fig 1A(x)) and vimentin (fig 1A(xiv)) expression. Co-treatment of the cells with TGF-β1 and TNFα dramatically accentuated the phenotype change (fig 1A(iv)) and further increased the expression of fibronectin (fig 1A(xii)) and vimentin (fig 1A(xvi)) compared with TGF-β1 alone.
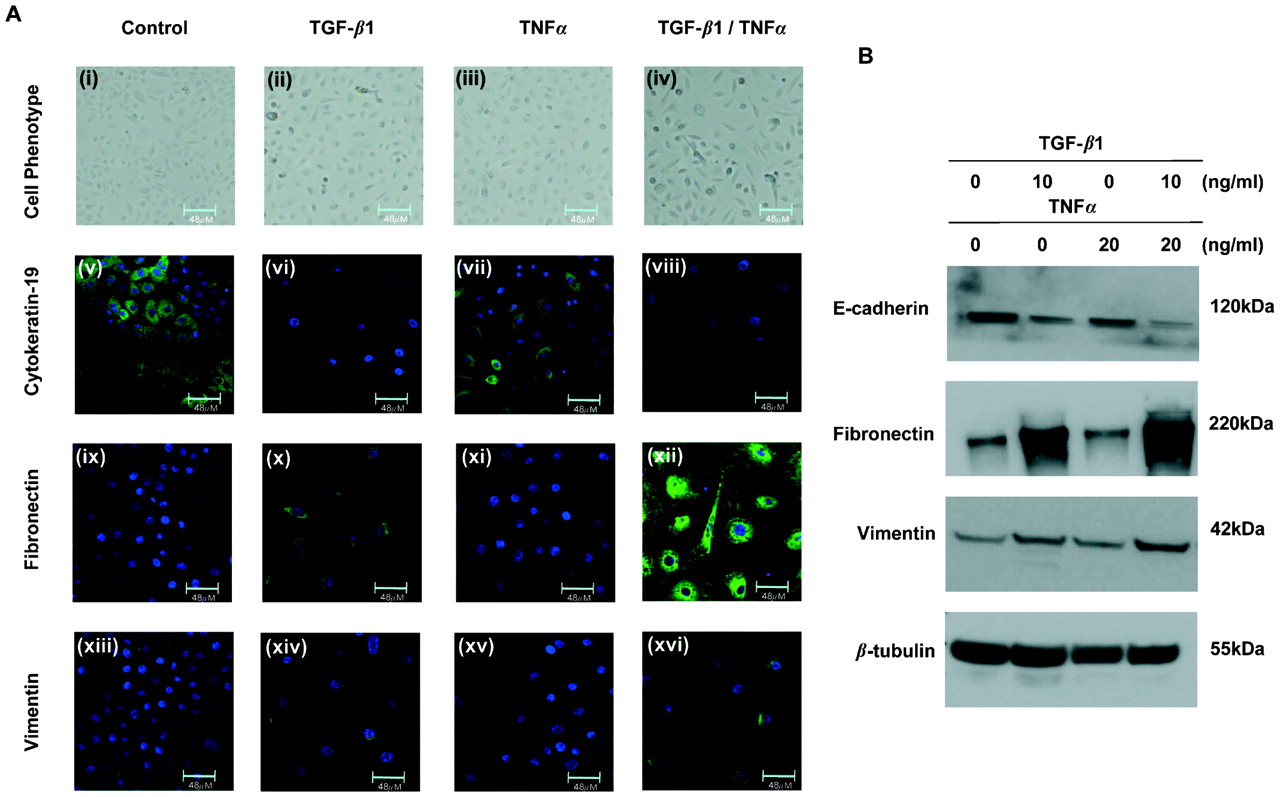
Change in protein expression induced by transforming growth factor β1 (TGF-β1) and tumour necrosis factor α (TNFα) in human primary bronchial epithelial cells (PBECs) from the transplanted lung. (A) PBECs left untreated (i) or treated with TNFα (iii) maintain the characteristic “cobblestone” appearance of epithelial cells, express cytokeratin-19 (v and vii) but express little to no fibronectin (ix and xi) or vimentin (xiii and xv). Cells treated with TGF-β1 begin to lose cell-cell contact and adopt an elongated phenotype (ii), downregulate cytokeratin-19 expression (vi) and increase fibronectin (x) and vimentin (xiv) expression. Co-treatment of the cells with TGF-β1 and TNFα induces a more dramatic change in phenotype (iv) and further downregulated the expression of cytokeratin 19 (viii) and upregulated fibronectin (xii) and vimentin (xvi) expression compared with TGF-β1 alone. All images shown at ×63 magnification. (B) Untreated cells and cells treated with TNFα express high levels of E-cadherin but little to no fibronectin or vimentin (lanes 1 and 3, respectively). Cells treated with TGF-β1 downregulate E-cadherin expression and upregulate fibronectin and vimentin expression (lane 2). Cells treated with TGF-β1 and TNFα further downregulate E-cadherin expression and upregulate fibronectin and vimentin expression compared with TGF-β1 alone (lane 4). All experiments repeated in five individual cell donors.
Western blotting was used to quantify the changes in protein expression (fig 1B). Untreated cells expressed high levels of E-cadherin but little vimentin or fibronectin. Treatment with TGF-β1 induced a statistically significant downregulation in E-cadherin expression (p<0.001, n = 5) and a marked increase in fibronectin (p<0.001, n = 5) and vimentin (p = 0.01, n = 5) expression compared with untreated cells. Co-treatment of the cells with TGF-β1 and TNFα induced a statistically significant change in protein expression compared with TGF-β1 alone (E-cadherin p = 0.006, n = 5; fibronectin p<0.001, n = 5; vimentin p = 0.005, n = 5). Treatment of the epithelial cells with TNFα alone had no effect on protein expression compared with untreated cells (E-cadherin p = 0.168, n = 5; vimentin p = 0.977, n = 5; fibronectin p = 0.624, n = 5).
Effect of TGF-β1 and TNFα concentrations on EMT marker expression
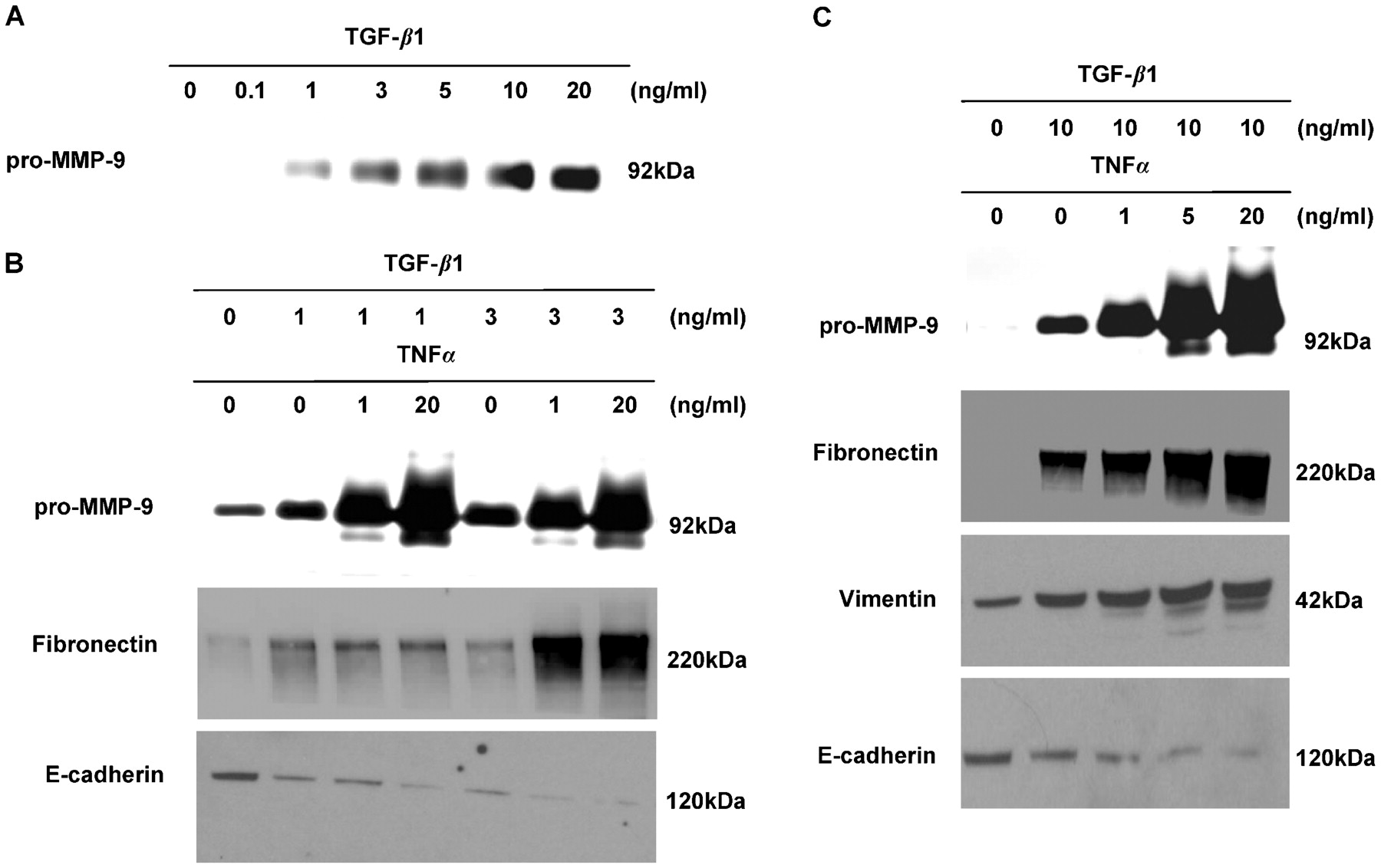
The effect of increasing concentrations of TGF-β1 (0.1–20 ng/ml) on the production of pro-MMP-9 was assessed using gelatin zymography. Control cells and cells treated with 0.1 ng/ml TGF-β1 secrete little or no pro-MMP-9 depending on the cell donor. However, cells treated with >1 ng/ml TGF-β1 secrete pro-MMP-9 in a dose-dependent manner (fig 2A). The accentuating effects of TNFα on EMT were further evaluated using lower doses of TGF-β1 (1 ng/ml and 3 ng/ml). A higher dose of TNFα (20 ng/ml) is required to accentuate EMT in the presence of a low dose of TGF-β1 (1 ng/ml) but, if the dose of TGF-β1 is increased to 3 ng/ml (fig 2B) or 10 ng/ml (fig 2C), a dramatic accentuating effect is achieved with as little as 1 ng/ml TNFα.

Accentuation of transforming growth factor β1 (TGF-β1)-driven epithelial mesenchymal transition (EMT) in human primary bronchial epithelial cells (PBECs) from the transplanted lung with low doses of tumour necrosis factor α (TNFα). (A) PBECs treated with increasing concentrations of TGF-β1 secrete increased levels of pro-matrix metalloproteinase-9 (pro-MMP-9) into the cell culture media compared with untreated cells. (B) Untreated cells express high levels of E-cadherin but little to no fibronectin and or pro-MMP-9 (lane 1). Cells treated with TGF-β1 at 1 ng/ml downregulate E-cadherin expression and upregulate fibronectin and pro-MMP-9 expression (lane 2). Addition of TNFα at 1 ng/ml upregulates pro-MMP-9 expression compared with TGF-β1 alone (lane 3). Addition of TNFα at 20 ng/ml further upregulates pro-MMP-9 expression and decreases E-cadherin expression (lane 4). Cells treated with 3 ng/ml of TGF-β1 downregulate E-cadherin expression and upregulate fibronectin and pro-MMP-9 expression (lane 5). Addition of TNFα at 1 ng/ml downregulates E-cadherin expression and upregulates fibronectin and pro-MMP-9 expression (lane 6). Addition of 20 ng/ml TNFα further upregulates pro-MMP-9 expression but has no effect on E-cadherin and fibronectin expression (lane 7). (C) Untreated cells express high levels of E-cadherin but little to no fibronectin, vimentin or pro-MMP-9 (lane 1). Cells treated with TGF-β1 at 10 ng/ml downregulate E-cadherin expression and upregulate fibronectin, vimentin and pro-MMP-9 expression (lane 2). Addition of TNFα induces a dose-dependent decrease in E-cadherin expression and a dose-dependent increase in fibronectin, vimentin and pro-MMP-9 expression (lanes 3–5).
Effect of TGF-β1 and TNFα on the invasive potential of epithelial cells
To determine whether airway epithelial cells are undergoing true functional EMT, their ability to penetrate a Matrigel-coated filter was investigated (fig 3A). Very few untreated cells penetrated the Matrigel filter (mean (SEM) 61 (28.4)), but the number of invasive cells increased following treatment with TGF-β1 compared with untreated cells (284 (38.7), p = 0.012, n = 5). TNFα alone had no effect on the number of invasive cells compared with untreated cells (84 (38.7), p = 0.672, n = 5). Co-treatment of the cells with TGF-β1 and TNFα significantly increased the number of invasive cells compared with TGF-β1 alone (514 (51.6), p = 0.037, n = 5).

Transforming growth factor β1 (TGF-β1) and tumour necrosis factor α (TNFα) induce increased invasive potential and deposition of the extracellular membrane (ECM) in human primary bronchial epithelial cells (PBECs) from the transplanted lung. (A) Quantitation of mean (SEM) number of invasive cells per treatment. Untreated or TNFα-treated PBECs show little to no invasion. TGF-β1-treated cells show an increase in invasion that is further accentuated by co-treatment of the cells with TGF-β1 and TNFα (*p = 0.037, n = 5). (B) PBECs treated with TGF-β1 (lane 2) secrete an increased level of pro-matrix metalloproteinase 9 (pro-MMP-9) into the cell culture media compared with untreated cells (lane 1). Treatment of the cells with TGF-β1 and TNFα accentuated the release of pro-MMP-9 compared with TGF-β1 alone (lane 3). (C) PBECs left untreated (lane 1) or treated with TNFα (lane 3) do not deposit collagen I or fibronectin. PBECs treated with TGF-β1 deposit collagen I and fibronectin (lane 2). Co-treatment of the cells with TGF-β1 and TNFα inhibits the TGF-β1-dependent deposition of collagen I while having no effect on fibronectin deposition (lane 4). All experiments performed in five individual cell donors.
The effect of TGF-β1, in the presence or absence of TNFα, on the production of pro-MMP-9 was assessed using gelatin zymography (fig 3B). Control cells secrete a low level of pro-MMP-9 that is increased by treatment with TGF-β1 (p = 0.007, n = 5). Co-treatment with TNFα further increased the secretion of pro-MMP-9 compared with cells treated with TGF-β1 alone (p<0.001, n = 5).
Effect of TGF-β1 and TNFα on deposition of ECM by epithelial cells
Cells treated with TNFα do not significantly increase secretion of collagen type I (p = 0.446, n = 5) or fibronectin (p = 0.990, n = 5) compared with untreated cells as assessed by TCA precipitation (fig 3C). However, TGF-β1 treatment induces a statistically significant increase in the secretion of collagen type I (p<0.001, n = 5) and fibronectin (p<0.001, n = 5) compared with untreated cells. Interestingly, co-treatment of the cells with TGF-β1 and TNFα had no statistically significant effect on the secretion of fibronectin (p = 0.156, n = 5) but significantly decreased the secretion of collagen type I (p = 0.004, n = 5) compared with TGF-β1 alone.
Expression of markers of EMT in vivo in airway epithelium
Biopsies were subjected to immunolocalisation studies for E-cadherin (fig 4A(i)–(iii)), α-SMA (fig 4A(iv)–(vi)) and vimentin (fig 4A(vii)–(ix)). The percentage of airway epithelium staining positive for each marker was quantified and compared with non-transplanted control lungs (fig 4B). Epithelium from normal control tissue was highly positive for E-cadherin (mean (SEM) 68.53 (2.54)%, n = 6) but expressed little α-SMA (6.69 (2.28)%, n = 6) or vimentin (2.05 (0.54)%, n = 6). Epithelium from stable transplant recipients showed a statistically significant decrease in E-cadherin expression (51.81 (2.28)%, p = 0.004, n = 6) and increase in vimentin expression (6.42 (0.77)%, p = 0.009, n = 6) compared with control tissue. α-SMA expression was not significantly different in the epithelium of stable transplant recipients compared with normal epithelium (4.89 (2.43)%, p = 0.657, n = 6). Epithelium in explanted lung tissue from patients with BOS showed a statistically significant decrease in E-cadherin expression (40.06 (3.98)%, p = 0.017, n = 8) and a statistically significant increased expression of α-SMA (21.21 (4.34)%, p = 0.008, n = 8) and vimentin (12.61 (2.32)%, p = 0.035, n = 8) compared with epithelium in both normal tissue and stable recipients.

Expression of markers of epithelial to mesenchymal transition (EMT) in vivo in human lung. (A) Lung sections from normal control lung ((i), (iv) and (vii)), from stable transplants ((ii), (v) and (viii)) and obliterative bronchiolitis (OB) affected tissue ((iii), (vi) and (ix)) were stained for E-cadherin ((i)–(iii)), α-smooth muscle actin (α-SMA) ((iv)–(vi)) and vimentin ((vii)–(ix)). An IgG control is shown (x). All sections shown at magnification ×100. (B) Mean (SEM) percentage of epithelial area per high power field (n = 5) stained positive for E-cadherin (i), α-SMA (ii) and vimentin (iii) in normal control lung (n = 6), stable transplant lung (n = 6) and OB affected lung (n = 8). *p<0.05.
To further evaluate EMT in vivo, lung tissue from patients with BOS was analysed for co-expression of E-cadherin and α-SMA or S100A4. Figure 5A shows an area of intact airway epithelium with clear E-cadherin expression at the junction between cells. In a number of the cells expressing E-cadherin we also see the expression of the early marker of EMT, S100A4 (see arrows), showing that these cells co-express both epithelial and mesenchymal markers. By comparison, in a more damaged area of epithelium (fig 5B) we see that E-cadherin, although still present, is reduced in intensity and very differently distributed throughout the cell. Furthermore, certain cells in the damaged epithelium also express α-SMA, a late marker of EMT, co-localising in E-cadherin positive cells (see arrows).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Co-expression of E-cadherin and S100A4/α-smooth muscle actin (α-SMA) in epithelial cells in the airway from recipients with obliterative bronchiolitis (OB). Sections from OB lung were stained for E-cadherin and S100A4 (A) or E-cadherin and α-SMA (B). (A) In an area of intact airway epithelium, E-cadherin (green) expression at the interface between cells is clearly visible. In some cells the expression of S100A4 (red) (arrows) is co-localised with E-cadherin. (B) In a more damaged area of epithelium, reduced E-cadherin (green) expression associated with an increase in the expression of α-SMA (red) is seen (arrows). Image acquired on a Leica confocal microscope (×63 magnification).
Discussion
In this study we have adopted a multifunctional approach to identification of EMT ex vivo using cell morphology, upregulation of mesenchymal proteins, downregulation of epithelial proteins, secretion of ECM and a functional assessment of invasive and migratory capacity. This multifunctional approach is likely to be much more robust than reliance on any single marker of EMT. In addition, our quantitative immunohistochemistry has demonstrated reduced epithelial E-cadherin expression and increases in vimentin and α-SMA expression in the airway epithelium of patients with BOS compared with stable recipients and control lung samples.
The results of this study add significantly to the understanding gained from our previously reported pilot observations in the lung transplant airway.10 The present study provides a comprehensive assessment of the ability of airway epithelial cells from the transplanted lung to undergo EMT. Our observations offer significant insight into what might be an important mechanism in the development of OB. The hypothesis that EMT represents a common final pathway in the development of airway remodelling after lung transplantation is attractive. It could explain why a broad range of insults, both alloimmune and non-alloimmune, could produce a similar pathological end point.
TGF-β1 has been widely implicated in driving fibrosis in a variety of organs such as the liver and kidney, but a mechanism by which this may occur in OB has not been fully elucidated. It has been previously observed that TGF-β1 expression is increased in patients with OB.11 12 13 14 In addition, studies in the rodent heterotrophic tracheal transplant models of OB have shown that interrupting TGF-β1 binding to its receptor or inhibiting its downstream signalling via the SMAD pathway reduces intraluminal airway matrix deposition.15
It is well recognised in clinical practice that BOS is often first diagnosed or progresses more rapidly following a significant episode of airway injury such as a viral or bacterial infection, and these sources of acute inflammation in the transplant airway may be very important in accentuating the process already driving OB. It has been previously suggested that repeated injury and persistent inflammation in the epithelium coinciding with defective regeneration is likely to lead to excessive fibroproliferation and obliteration of small airways in OB.25 The role played by pro-inflammatory cytokines in driving excessive fibroproliferation is unclear, and our data may highlight a possible novel mechanism in the lung.
Alveolar macrophages play a crucial role in initiating an acute inflammatory response in the lung and, on activation, produce a multitude of pro-inflammatory cytokines and growth factors.26 A prominent product produced by macrophages is TNFα which is believed to play a role in fibrotic disorders.27 Also, an increase in the level of intracellular TNFα found in CD4+ and CD8+ T cells during acute lung transplant rejection has been reported,28 and TNFα can increase metastatic potential in human colonic epithelial organoid models of colon cancer by accentuating EMT.29 The addition of TNFα to cultures containing TGF-β1 significantly increased the expression of vimentin and fibronectin while further downregulating E-cadherin and cytokeratin-19 expression compared with TGF-β1 alone. Our observations, together with those of previous studies, suggest a possible link between acute inflammation and accentuated TGF-β1-driven epithelial remodelling.
The demonstration of EMT in cultured cells always causes debate as to whether the observations represent true EMT or simply a “stress response” of cells in culture. It is our opinion that demonstrating true transition from an epithelial to a mesenchymal phenotype requires analysis of functional properties as well as protein expression and morphology. The secretion of MMP is one functional characteristic of mesenchymal cells. The ability of our ex vivo cultures to acquire an invasive phenotype was analysed. TGF-β1 induces a threefold increase in the number of invasive cells which is associated with an increased secretion of pro-MMP-9. As with protein expression, co-treatment of the cells with TGF-β1 and TNFα further accentuated the effects with an increase in both pro-MMP-9 production and the number of invasive cells observed. These observations support our hypothesis that epithelial cells in the airway may undergo EMT, penetrate the basement membrane and migrate into the subepithelial layer to lay down excessive connective tissue.
The cells that drive the obstruction of the bronchioles in OB are capable of depositing ECM proteins such as fibronectin and collagen I. We confirmed that TGF-β1 induced a marked increase in collagen I secretion. However, in the presence of TNFα this was inhibited. This result is in agreement with previous studies showing that TNFα can inhibit collagen production both in vivo and in vitro.30 31 32 Interestingly, although TGF-β1 induced an increase in fibronectin deposition, the addition of TNFα had no significant effect on this which suggests that different pathways may control collagen I and fibronectin deposition. This raises the intriguing possibility that episodic release of TNFα in the presence of TGF-β1—such as could occur in response to repeated inflammatory insults to the lung—could drive migration and invasion of transformed epithelial cells but not deposition of collagen until the levels of TNFα fall again, in which case the transformed cells could then drive fibrosis.
The changes seen in expression levels of E-cadherin, vimentin and α-SMA in the epithelium of OB lung compared with stable transplant recipients as well as our finding of co-localisation of epithelial and mesenchymal markers in individual cells in the epithelium of OB lung support our hypothesis. In this study, areas of intact epithelium were analysed in OB, stable and control lung. We did not analyse severely remodelled airways in the OB lung as these areas represent advanced lesions where the identification of the epithelium is impossible. Instead, our approach allowed a direct comparison between marker expression in the study groups.
Our data do not prove a definitive causative link between the remodelling seen in BOS and EMT in the airway. However, we believe these observations represent a significant increase in the understanding of the potential for airway epithelium to undergo EMT and, in addition, highlight a possible synergistic action between acute inflammation and the ability of TGF-β1 to drive EMT. The mechanism driving this synergy needs more investigation to identify possible therapeutic targets that may interfere with this process in the lung.
REFERENCES
Supplementary materials
Web only appendix 64;9:770
Files in this Data Supplement:
Web only appendix 64;9:770
Files in this Data Supplement:
Footnotes
See Editorial, p 742
▸ Additional Method details are published online only at http://thorax.bmj.com/content/vol64/issue9
Funding This work was supported by a research grant from the Medical Research Council UK. AJF is supported by a GlaxoSmithKline clinical fellowship award.
Competing interests None.
Provenance and Peer review Not commissioned; externally peer reviewed.