Article Text

Abstract
Background: Subepithelial collagen and extracellular matrix protein deposition are important pathophysiological components of airway remodelling in chronic asthma. Animal models based on the local reaction to antigens show structural alterations in the airway submucosal region and provide important information regarding disease pathophysiology. We describe a murine model of peribronchial fibrosis using intratracheally instilled transforming growth factor (TGF)-β1 in BALB/C mice that facilitates a mechanistic approach to understanding the cellular and molecular pathways leading to airway fibrosis.
Methods: BALB/C mice were intratracheally instilled with either TGF-β1 or buffered saline. Airway fibrosis was assessed by light microscopy, hydroxyproline content, and polymerase chain reaction (PCR) for collagen I and III on microdissected airway samples. The lysyl oxidase inhibitor β-aminoproprionitrile (BAPN) was administered to TGF-β1 treated mice to block airway collagen deposition. Airway hyperresponsiveness was also measured after treatment with TGF-β1.
Results: During the 7 days after administration of TGF-β1 the mice developed increased subepithelial collagen which could be blocked by BAPN. Increased mRNAs for collagen types I and III were seen in microdissected airways 1 week after TGF-β1, and significantly increased total collagen was found in the airways 4 weeks after TGF-β1. A detectable increase in airway hyperreactivity occurred.
Conclusions: This new model should facilitate detailed study of airway remodelling that occurs in the absence of detectable cellular inflammation, and allow examination of the functional consequences of a major structural alteration in the conducting airways uncomplicated by inflammatory cell influx.
- collagen
- fibrosis
- transforming growth factor β1
Statistics from Altmetric.com
Remodelling of the conducting airways is an important pathophysiological process in many individuals with chronic asthma. Structural changes to the airway are believed to contribute to an irreversible decrement in lung function in these individuals,1–4 but this association has not been proven. Subepithelial deposition of collagen (types I, III, and V) and other extracellular proteins, fibroblast proliferation, mucus hypersecretion, and smooth muscle thickening are all evident in airway remodelling in many chronic asthmatics, but the cascade of events leading to these alterations is not well understood. There is a need for animal models that will allow elucidation of the mechanisms controlling each of these structural changes.
Antigen derived murine models of asthma mimic some, but not all, of the physiological and histological alterations in the lungs of asthmatic patients. Exposure of sensitised mice to ovalbumin, for example, causes chronic eosinophilic bronchitis5–7 and expansion of the subepithelial compartment with collagen deposition and myofibroblast proliferation. An innovative approach addressing these late stage changes and other specific components of airway remodelling in the absence of chronic bronchitis would be desirable. Indeed, a recent consensus workshop suggested that models focusing on specific contributors to the airway remodelling response would aid in identifying the factors involved in this process.8
Transforming growth factor-β1 (TGF-β1) appears to play an integral role in promoting the structural changes of airway remodelling, and is a focus of study both mechanistically and therapeutically.9–11 TGF-β1 belongs to a family of growth factors of critical importance in wound healing and fibrogenesis. It has been studied extensively in models of liver,12 kidney,13 and lung fibrosis.14,15 In asthmatics, increased TGF-β1 mRNA expression in bronchial biopsy specimens is seen compared with normal subjects, and levels correlate with the depth of subepithelial fibrosis.9 In addition, the content of TGF-β1 is increased in the bronchoalveolar lavage fluid in asthmatics,10 and is significantly increased in severe asthmatics with a prominent eosinophil influx in the airways.16 TGF-β1 may not only stimulate differentiation of fibroblasts to myofibroblasts, but also inhibit apoptosis of the myofibroblasts in the lung parenchyma causing extended survival of this population and excessive collagen deposition.17
We have developed a new model of late stage airway remodelling by direct intratracheal instillation of TGF-β1 into the lungs of mice. We hypothesised that, independent of a prominent inflammatory response, TGF-β1 could promote extracellular matrix collagen deposition in the airway submucosa.
METHODS
Intratracheal instillation of TGF-β1
BALB/C mice (16–20 g) purchased from Charles River Breeding Laboratories (Wilmington, MA, USA) were housed in the Animal Resources Center at our facility. Strains ICR, C57/BL6, and A/J were purchased from the same supplier. All mice were provided standard feed and water ad libitum for several days before the procedure to acclimatise to their new surroundings. Anaesthetised mice were positioned supine on an angled restraining stand and were orotracheally intubated with a long 22 gauge blunt tipped metal catheter.18–20 Endotracheal positioning was confirmed by palpation of the tracheal rings with the catheter tip. Immediately after intubation, 40 μl phosphate buffered saline (PBS, pH=7.4) or human TGF-β1 (50–400 ng; Promega, Madison, WI, USA) diluted in PBS was instilled by injection. The solution was distributed throughout the lungs by injection of a bolus of 50 μl air. Identical instillations were repeated in each mouse 24 hours later and the animals were returned to their cages for a further 5 days.
β-aminoproprionitrile (BAPN) injections
In a subset of the animals injected with TGF-β1, 0.5 ml intraperitoneal injections of 200 ng/kg of β-aminoproprionitrile (BAPN, n=9) or PBS (n=6) were given twice daily on days 1–6 after the first orotracheal administration of TGF-β1. In the BAPN treated group four mice received a total of 150 ng TGF-β1 and five received a total of 200 ng. All mice experienced a total experimental interval of 7 days.
Tissue staining
Animals were killed with an overdose of inhaled isoflurane and their lungs were fixed with an intratracheal infusion of 1% paraformaldehyde at a pressure of 25 cm H2O for 1 hour, then immersed in fixative overnight at 4°C. After fixation the lungs were placed in 70% ethanol and prepared for paraffin embedding. Lung sections of 5 μm thickness were made with special attention to cutting through the larger lobar bronchi in parallel. Dried lung sections were stained with 0.1% Picro-Sirius red stain F3BA (Pfaltz and Bauer, CT, USA)/0.5% picric acid for 30 minutes after deparaffinising.
Airway microdissection
Lobar bronchi and distal airways were dissected from either the right lung or both lungs under a dissecting microscope. For protein and hydroxyproline assays, the right lung was dissected from lungs fixed for 24 hours with paraformaldehyde and stored in 70% ethanol at 4°C. For mRNA determinations by real time polymerase chain reaction (PCR), lungs were dissected as quickly as possible, frozen immediately on dry ice, and stored at −80°C before analysis.
Protein and hydroxyproline determinations
The Micro BCA Protein Assay Reagent Kit (Pierce, Rockford, IL, USA) was used to determine the total protein content of the airway samples. Half of the airway preparation was homogenised in water with a Branson Sonifier 250 (VWR Scientific, San Francisco, CA, USA; duty cycle 40, output control 4) until the tissue was completely homogenised. Standard curves using bovine serum albumin were linear between 0 and 32 μg; 1:10 dilutions of 50 μl airway homogenate were routinely assayed, measuring the chromophore at 562 nm.
The hydroxyproline content of airway hydrolysates (6N HCl, 105°C, 24 hours) was measured using a colorimetric method.21 Standard curves were linear between 0 and 32 μg hydroxyproline.
RNA isolation from microdissected airways
Total RNA was isolated from fresh or frozen airways using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. 1 ml Trizol reagent was added to frozen airway samples and the resulting preparation was ground using a mortar and pestle for 5 minutes. Chloroform (200 μl) was added and the solution was centrifuged (12 000 rpm, 4°C) for 20 minutes. The aqueous layer was removed by aspiration with a pipette and an equal volume of isopropanol was added to the aqueous layer. After centrifugation for 17 minutes as above, the supernatant was discarded and the remaining pellet was washed in 75% ethanol and suspended in 20 μl DNase- and RNase-free water. Five μg of the total RNA was reverse transcribed into cDNA with Moloney murine leukemia virus reverse transcriptase (Invitrogen), following the manufacturer’s instructions.
Real time PCR analysis (RT-PCR)
A quantitative RT-PCR assay was performed using gene specific double labelled fluorescent probes and sets of specific primers in an ABI PRISM 5700 sequence detection system (PE Applied Bio-Systems, Foster City, CA, USA). PCR was performed in a 50 μl volume with 5.0 μl of a cDNA sample, 1 μmol/l of both forward and reverse primers, 2 mM MgCl2, 0.040 U/μl AmpliTaq Gold DNA polymerase, 200 nmol of probe, and 400 μM deoxyribonucleotide triphosphate mixture in 1× PCR-Gold buffer. Reaction mixtures were incubated at 50°C for 30 minutes. After inactivation of reverse transcriptase at 95°C for 12 minutes, 40 cycles of amplification were performed with denaturation at 95°C for 15 seconds and both annealing and extension at 60°C for 1 minute each. The primers for the α1(I) chain and α1(III) chain genes from mouse collagen and the β-actin gene from mouse were as follows: α1(I) chain gene for mouse collagen: (forward) ACGGCTGCACGAGTCACAC, (reverse) GGCAGGCGGGAGGTCTT; α1(III) chain gene for mouse collagen: (forward) GTTCTAGAGGATGGCTGTACTAAACACA, (reverse) TTGCCTTGCGTGTTTGATATTC; β-actin gene from mouse: (forward) ACTATTGGCAACGAGCGGTT, (reverse) CAGGATTCCATACCCAAGA AGGA.
Standard curves were constructed with the use of dilutions of a standard cDNA preparation from mouse airway (arbitrarily chosen) containing the RT-PCR products of each of the genes we wanted to analyse. To compensate for any differences in cell number and/or recovery of RNA, the copy number of each of the collagen mRNAs was determined relative to the β-actin mRNA determined simultaneously.
Airway reactivity
To assess airway hyperresponsiveness to methacholine a non-invasive whole body mouse plethysmograph (Buxco, Troy, NY, USA) was used to measure a derived index named “enhanced pause” (Penh).22 Penh is a reasonable correlate of airway responsiveness to non-specific inhaled stimuli such as methacholine, and provides an acceptable measure for comparison between experimental groups.22,23
Exposure to ovalbumin
The airway reactivity of TGF-β1 instilled mice was compared with that in ovalbumin sensitised mice. Four mice were sensitised via two intraperitoneal injections of 10 μg/0.1 ml chicken egg albumin (ovalbumin grade V=98% pure; Sigma, St Louis, MO, USA) with alum as an adjuvant on days 1 and 15 of the experiment.7 Exposures to ovalbumin aerosol, 10 ml of a 10 mg/ml (1%) solution, were started on day 28. Mice were exposed for up to 60 minutes three times a week for the duration of a given experiment. A sidestream nebuliser (Invacare Corporation, Elyria, OH, USA), ProNeb compressor nebuliser (Pari, Richmond, VA, USA), and Passport Compressor (Invacare, Sanford, FL, USA) were used to generate the aerosols. We have previously shown that the mass median aerodynamic diameter of the aerosol in our two nebulisers is 1.88 (0.14) μm and 1.95 (0.21) μm.24
Statistical analysis
Instat software (Graph Pad Software, San Diego, CA, USA) was used to analyse the data. A two tailed Mann-Whitney rank sum test for non-parametric comparisons was performed unless otherwise specified. Data are presented as mean (SE) values. A p value of <0.05 indicated that the differences were significant.
RESULTS
Sirius Red stain
All of the study animals were graded. Correlation between staining scores and the dose of TGF-β1 instilled was good (r=0.88). Sirius Red staining of airways from mice observed 1 week after instillation of TGF-β1 at a total dose of 150–400 ng showed increased collagen deposition in the subepithelial matrix compared with control animals instilled with PBS. Comparison of the stain scores between the individual readers showed no significant difference (TGF-β1 scores (n=12) were 2.9 (0.31) and 3.1 (0.40) for readers 1 and 2, respectively, p=0.48).
There was a highly significant difference between the staining scores in the TGF-β1 instilled mice and those in the PBS instilled controls (2.9 (0.31) v 1.2 (0.14), p=0.0015; n=12 and n=9, respectively; fig 1). Increased Sirius Red staining was seen up to 8 weeks after TGF-β1 instillation, the longest duration so far examined. After administration of BAPN for 1 week there was a significant reduction in stain intensity and depth in the treated group (n=9) compared with the TGF-β1 group (n=12) (1.14 (0.14) v 2.9 (0.31), p=0.0012; fig 2). There was no significant difference in stain scores between the PBS control and BAPN treated mice (1.2 (0.14) v1.14 (0.14), p=0.42).

Sirius Red stain grade for each group. Data are expressed as mean (SE) staining score. A grading system of 0–4 was established before the grading; airways were scored under ×100 power. The grading scale used was: 0=no peribronchial Sirius red stain; 1=evident Sirius red stain but not consistently increased in all airways; 2=slight consistent increase in Sirius red depth of stain; 3=increased uniform stain depth throughout the airways; 4=dramatically increased depth of stain in all airways. *p<0.05.

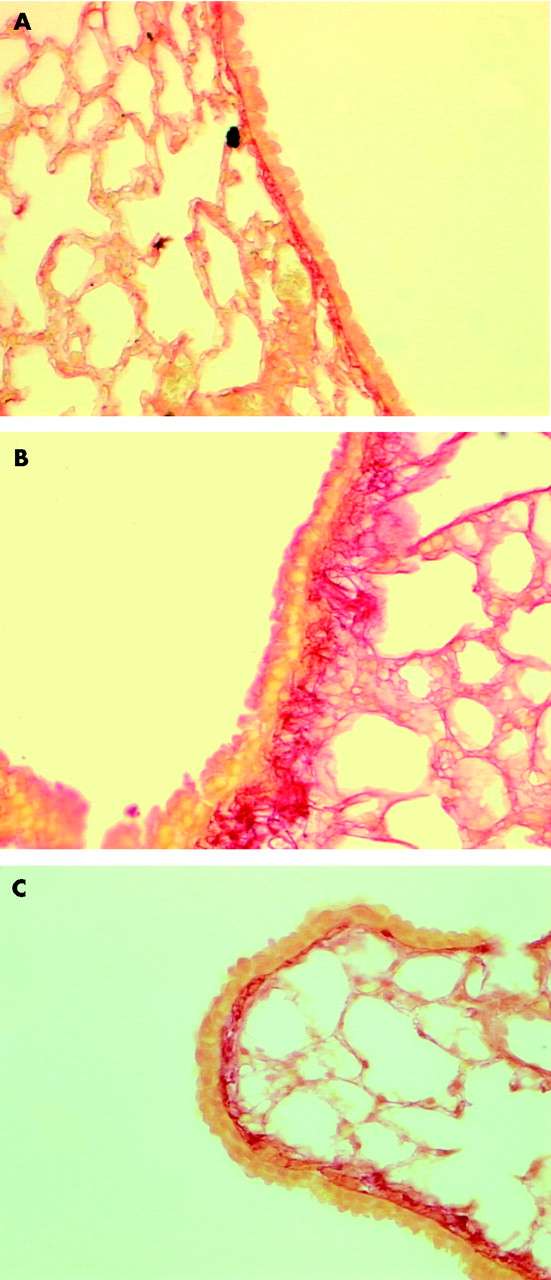
Sirius Red stained lung sections of mice (×200). (A) Phosphate buffered saline instilled control mouse; (B) transforming growth factor (TGF)-β1 instilled mouse (dose 200 ng); (C) TGF-β1 instilled mouse (dose 200 ng) treated with BAPN.
Response of various mouse strains to TGF-β1
Increased collagen deposition was seen in the peribronchial extracellular matrix of mice from three additional strains (ICR, A/J, and C57/BL6; n=8 each) compared with PBS instilled controls after intratracheal instillation of 100–400 ng TGF-β1. There were no significant differences in stain scores between the four mouse strains tested in response to TGF-β1 as evaluated by Sirius Red stain scores at 7 days.
Lavage cell counts
As shown in table 1, there was no significant difference in either the total number of cells or the relative percentage of pulmonary alveolar macrophages in whole lung lavages of control mice (instilled with PBS) and mice instilled with TGF-β1 Cell counts are given as per 10 high power fields. Total cell number lavaged can be estimated by multiplication by 900. Almost all (>90%) of the cells recovered from the mice treated with TGF-β1 were alveolar macrophages, suggesting a lack of inflammatory cell infiltration. No goblet cells were seen in the epithelium of TGF-β1 instilled mice after 7, 14, or 28 days, or in the control animals. By contrast, 25% of the epithelial cells in BALB/c mice exposed to ovalbumin for 4 weeks were PAS positive (data not shown).
Cell number and differential cell counts in lung lavage fluid
Airway total collagen determinations
The hydroxyproline content of each airway preparation was normalised to the mean protein level of the dissected airways in each group to correct for small differences in the yield of airways on microdissection. There were no significant differences in the hydroxyproline content of the control group at any of the three time points examined, so results for all three time points were combined. The amounts of measurable hydroxyproline were significantly greater 4 weeks after exposure to TGF-β1 (n=3) than in the pooled PBS control group (n=9; fig 3); there was an apparent trend towards increased amounts of hydroxyproline by week 2 (n=3).
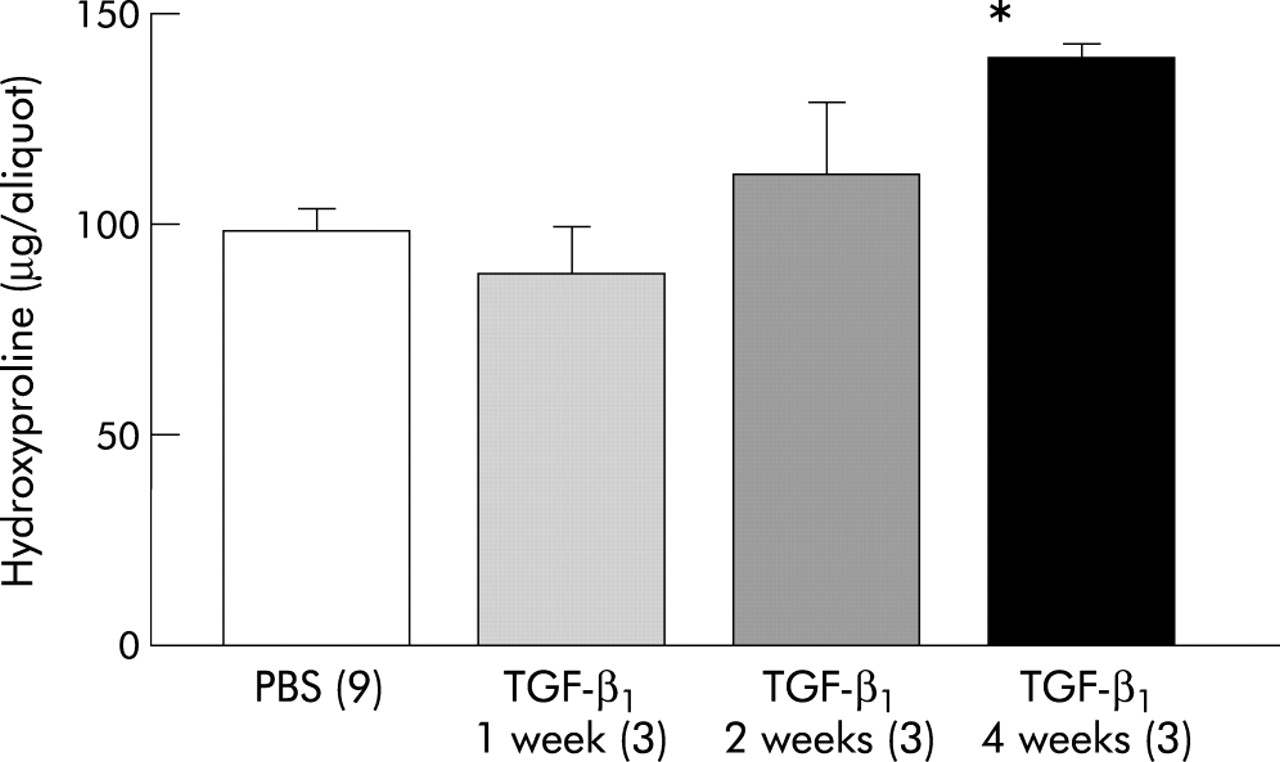
Hydroxyproline content of microdissected airways from mice at 7, 14, and 28 days. Data are expressed in μg/aliquot of lung hydrolysate ±1 SE. *p<0.05 compared with phosphate buffered saline (PBS) control.
Collagen mRNA content of isolated airways
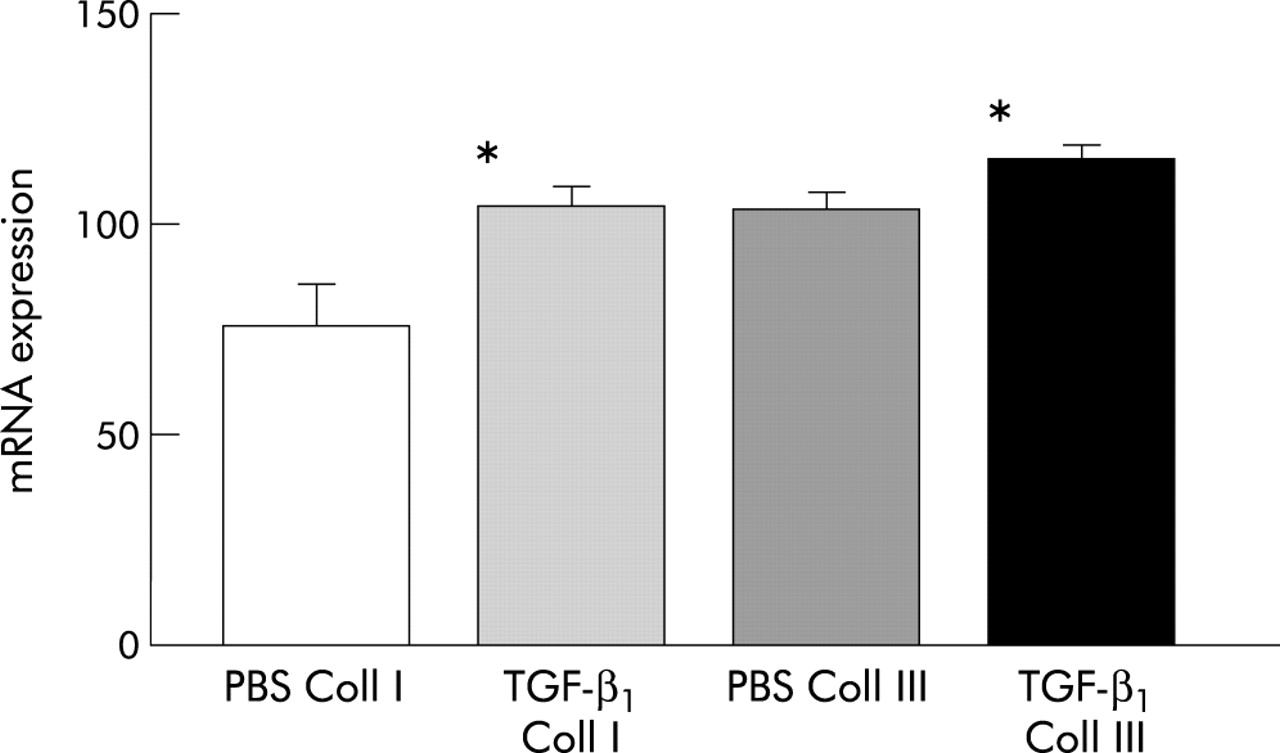
A significant increase in mRNA expression for collagen types I and III was seen 1 week after instillation of TGF-β1 compared with the control group (TGF-β1v PBS; α1(I): β-actin mean (SD) 104.4 (9.9) (n=5) v 75.4 (19.8) (n=4), p=0.011; α1(III): β-actin 116.2 (7.8) (n=5) v 103.5 (8.7) (n=4), p=0.03, one tailed t test; fig 4) Quantification of mRNA levels relative to the housekeeping gene β-actin by real time PCR indicated that the relative increase in the α1(I) and α1(III) mRNAs was approximately equal.

Airway mRNA expression for collagen I and III (40 cycles PCR) in mice instilled with transforming growth factor (TGF)-β1 (200 ng) and PBS. mRNA levels for collagen I and III are expressed relative to the housekeeping gene β-actin ±1 SE. *p<0.05 compared with phosphate buffered saline (PBS).
Airway responsiveness
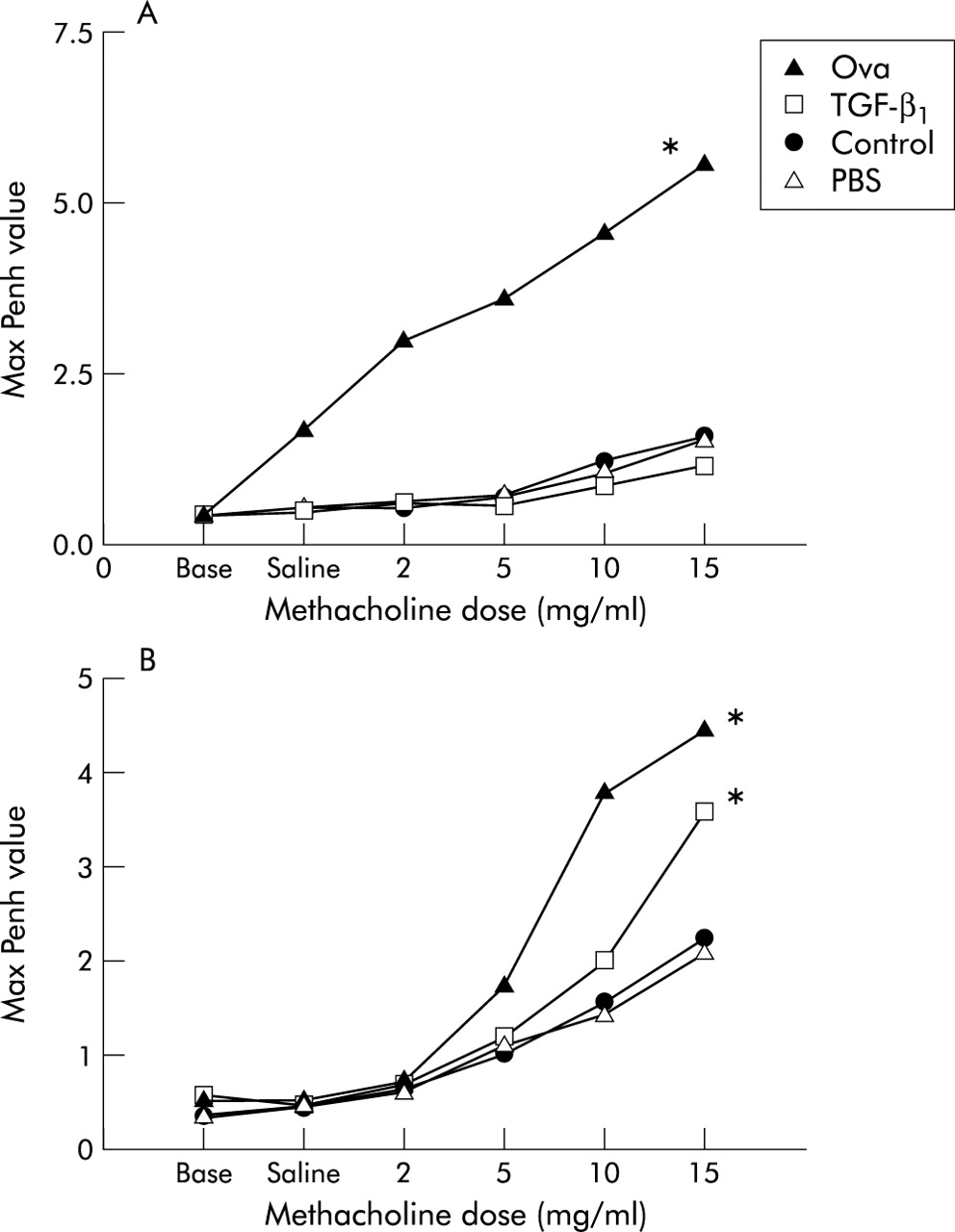
No change in Penh was observed in mice treated with TGF-β1 compared with saline instilled control animals 1 week after instillation (fig 5A). At 4 weeks mean Penh values were significantly greater in the TGF-β1 group than in the PBS or air control groups (fig 5B), but they were not as high as those of ovalbumin exposed mice.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Airway hyperresponsiveness in control BALB/C mice and mice instilled with either transforming growth factor (TGF)-β1 (200 ng) or phosphate buffered saline (PBS) at (A) 1 week and (B) 4 weeks. Data are expressed as mean maximum “enhanced pause” (Penh) values at increasing doses of methacholine for the group. Penh values of BALB/C mice exposed for 1 and 4 weeks to ovalbumin are shown as a positive control. There was a significant difference between the ovalbumin and the PBS instilled and air control groups at 1 and 4 weeks. *p<0.05 compared with controls (n=4 each) and between the TGF-β1 and PBS instilled and air control groups at 4 weeks.
DISCUSSION
TGF-β1 caused airway fibrosis with peribronchial collagen deposition in the subepithelial matrix of mice after 7 days without causing a significant inflammatory response or widespread fibrosis in the lung parenchyma. This change appears to resemble the late stage subepithelial collagen deposition seen with airway remodelling in some chronic asthmatics. BAPN, a known inhibitor of collagen cross linking and of collagen deposition, diminished the amount of this matrix protein detectable by Sirius Red staining. The increase in measurable peribronchial staining after TGF-β1 instillation correlated with an increase in airway collagen measured as 4-hydroxyproline and an increase in the airway content of mRNA for both collagen I and III.
Airway remodelling is a complex process and targeting specific cellular and molecular aspects of this process in vivo is difficult. Models that allow us to focus on specific components or mediators of the remodelling cascade are potentially beneficial. Ovalbumin sensitised mice have many of the physiological and pathological consequences seen in human asthma; they develop a pattern of chronic airway injury with structural changes consistent with remodelling.7,25,26 However, relatively large numbers of mice need to be exposed to ovalbumin for many weeks before studies of airway fibrosis can be performed.6,7 Interpretation of the cellular and molecular mechanisms underlying the observed changes is confounded by the severe inflammation provoked by this exposure regimen. The TGF-β1 model reproduces the late stage structural changes of airway fibrosis, dramatically minimising the time required for fibrosis to be manifest and thereby facilitating intervention studies such as in our experiment with BAPN. After 1 week subepithelial collagen deposition in BALB/c mice was similar histologically to that observed after 6–10 weeks of exposure to ovalbumin aerosol 4 weeks after sensitisation with intraperitoneal ovalbumin. Mechanistic inferences from simplified animal models such as TGF-β1 instillation can be further tested in more traditional animal models such as ovalbumin exposed mice.
There is increasing evidence to link the ubiquitous repair protein TGF-β1 with the airway changes seen in patients with persistent asthma.27 The upregulation via adenovirus vector of SMAD 7, an antagonist of TGF-β transduction signalling, significantly reduced bleomycin induced lung fibrosis.28 Treatment with anti-TGF antibody produced a favourable response in the lungs of bleomycin exposed mice.14 Similar approaches may also prove efficacious in models of chronic asthma. The possibility of studying the role of TGF-β1 in airway fibrosis independent of the contributions of other growth factors and cytokines seen with a prominent inflammatory response in an in vivo animal model is a highly attractive, if extremely reductionist, approach. It allows for focused study of peribronchial fibrosis and, perhaps, the role of the myofibroblast as it might pertain to airway wall remodelling. TGF-β1 may be acting by stimulating fibroblast differentiation to myofibroblasts, prolonging survival of the activated myofibroblasts by inhibiting apoptosis,29,30 or by other mechanisms not yet elucidated. This model should allow us to test these hypotheses directly.
The total number of cells recovered by lavage from the lungs of mice treated with TGF-β1 was the same as from the control animals, and more than 90% of the recovered cells were unambiguously classifiable as pulmonary alveolar macrophages. These animals therefore did not exhibit a clinically detectable lung inflammatory response. These results indicate that intratracheal administration of TGF-β1 is by itself adequate and sufficient to elicit increased collagen synthesis in the conducting airways of the treated mice. TGF-β1 in this model is acting directly as a fibrogenic stimulus for target cells in the airway submucosal region and is not acting indirectly as a proinflammatory cytokine.
BAPN significantly reduced the deposition of collagen in the subepithelial matrix in mice treated with TGF-β1. These results are consistent with previous studies of organ and tissue fibrosis in other models.31 We recognise that BAPN is a toxic compound in humans and would not serve as an appropriate therapeutic agent to prevent collagen deposition and cross linking chronically in the asthmatic population. However, it provides a good proof of concept, as well as a point of departure for further treatment trials.
We were concerned that the observed effects of intratracheal instillation of TGF-β1 could be an effect isolated to the BALB/C mouse strain, which is particularly sensitive to the effects of aerosolised ovalbumin and other stimulants of the Th2 response.32 Studies with other animal models have shown a variation in the inflammatory and fibrotic response to insult in different mouse lines.33 It is likely that the genetic locus for differing susceptibility to ovalbumin is upstream of TGF-β1 in the inflammatory cascade. Nevertheless, we believe our finding that the four mouse strains tested had similar airway changes following administration of intratracheal TGF-β1 supports the general validity of our results and suggests that these findings in the BALB/c mouse strain in response to this growth factor are likely to be predictive of findings in other animal models.
To evaluate the subepithelial changes in collagen occurring after TGF-β1 administration, we graded the airway changes by light microscopy. Neither evaluation with computer assisted image analysis techniques nor electron microscopy were used to assess the airway changes systematically in all lung sections. We chose to evaluate the sections stained with Sirius Red using low power (×100) light microscopy because we believed this lessened the possibility of selection bias. Our analysis focused on reviewing the entire lung section with description of the changes at the level of the lobar and distal bronchi. We developed a consistent scoring system that was easily reproducible by independent reviewers. Systematic morphometric evaluation has been found to be a reliable and consistent measure in other models,14 but we did not find this method provided more accurate information in a subsample of lung sections tested. Our findings by light microscopy were confirmed by biochemical analysis of airways microdissected from mouse lungs. Increased levels of 4-hydroxyproline were evident 4 weeks after TGF-β1 infusion, while mRNAs for collagens I and III were significantly increased in the experimental group at 7 days.
We measured Penh, a derived value that can be obtained non-invasively from unrestrained animals in mice instilled with TGF-β1. At 4 weeks we found a significant difference between animals instilled with TGF-β1, mice with fibrotic airways, and matched control animals. The Penh values seen in the TGF-β1 group were not as high as those seen in the ovalbumin exposed mice. It appears that airway fibrosis in the absence of inflammation may be a contributing factor to the chronic airway hyperreactivity in mice exposed to methacholine.
We have developed a novel murine model of late stage airway remodelling with increased collagen and extracellular protein deposition in the subepithelial matrix. Other components of airway remodelling such as goblet cell hyperplasia were not observed. Our approach allows for focused study in a well controlled model of a specific, and presumably detrimental, aspect of airway change in subjects with chronic asthma. While not a complete representation of the asthmatic airway, this in vivo model may be helpful in performing various mechanistic and therapeutic intervention experiments. Future research will focus on identifying the specific cell populations transformed by TGF-β1 and the cellular transduction mechanisms activated. Inhibition of the effects of TGF-β1 at this level in the remodelling process may be therapeutically appealing.
Acknowledgments
This work was supported in part by a Faculty Research Grant from the University of California, Davis Academic Senate and by a Research Training Fellowship from the American Lung Association of California (NJK) and grants from the Philip Morris Research Management Group, from NIEHS (ES05707), and from the University of California Health System (JAL).