Article Text

Abstract
Background Streptococcus pneumoniae is the most commonly identified pathogen in community-acquired pneumonia (CAP). Myeloid-related protein (MRP) 8/14 is a major component of neutrophils that is released upon infection or injury. MRP8/14 is essential for protective immunity during infection by a variety of micro-organisms through its capacity to chelate manganese and zinc. Here, we aimed to determine the role of MRP8/14 in pneumococcal pneumonia.
Methods MRP8/14 was determined in bronchoalveolar lavage fluid (BALF) and serum of CAP patients, in lung tissue of patients who had succumbed to pneumococcal pneumonia, and in BALF of healthy subjects challenged with lipoteichoic acid (a component of the gram-positive bacterial cell wall) via the airways. Pneumonia was induced in MRP14 deficient and normal wildtype mice. The effect of MRP8/14 on S. pneumoniae growth was studied in vitro.
Results CAP patients displayed high MRP8/14 levels in BALF, lung tissue and serum. Healthy subjects challenged with lipoteichoic acid demonstrated elevated MRP8/14 in BALF. Likewise, mice with pneumococcal pneumonia had high MRP8/14 levels in lungs and the circulation. MRP14 deficiency, however, was associated with reduced bacterial growth and lethality, in the absence of notable effects on the inflammatory response. High zinc levels strongly inhibited growth of S. pneumoniae in vitro, which was partially reversed by MRP8/14.
Conclusions In sharp contrast to its previously reported host-protective role in several infections, the present results reveal that in a model of CAP, MRP8/14 is misused by S. pneumoniae, facilitating bacterial growth by attenuating zinc toxicity toward the pathogen.
- Bacterial Infection
- Cytokine Biology
- Innate Immunity
- Neutrophil Biology
- Respiratory Infection
Statistics from Altmetric.com
Key messages
What is the key question?
-
What is the role of myeloid-related protein (MRP)8/14 in Streptococcus pneumoniae-induced community-acquired pneumonia (CAP) in a clinical and experimental setting?
What is the bottom line?
-
MRP8/14 is compartmentally released in CAP and is associated with increased lethality in a mouse model of CAP through the facilitation of pneumococcal growth by attenuating zinc toxicity toward the pathogen.
Why read on?
-
In sharp contrast to the previously reported host protective role of MRP8/14 in several infections, this study represents the first in vivo example of how a pathogen can misuse this protein for its own benefit.
Introduction
Community-acquired pneumonia (CAP) is a common infectious disease and a leading cause of sepsis.1 ,2 The most prevalent causative micro-organism in CAP is the Gram-positive diplococcus Streptococcus pneumoniae.1–3 Pneumococci are increasingly antimicrobial resistant,4 thus stressing the importance of gaining more insight into host defence mechanisms that influence the outcome of this prominent infection.
When pneumococci invade the lower respiratory tract, various inflammatory mechanisms ensue, including infiltration and activation of neutrophils,3 ,5 which is associated with the release of complexes of myeloid-related protein (MRP)8 and MRP14 (MRP8/14 or calprotectin).6 ,7 MRP8/14 induces a variety of innate immune responses, including induction of leukocyte recruitment and cytokine release via Toll-like receptor (TLR)4.7 In addition to its proinflammatory effects, MRP8/14 possesses direct antimicrobial properties directed against bacteria8–10 and fungi.11 ,12 MRP8/14 mediates at least part of these effects by chelation of manganese and zinc, elements that are generally important for microbial growth and virulence.13 Previous investigations have shown that manganese is critical for pneumococci to manage oxidative stress, allowing growth under normal aerobic conditions.14–17 Importantly, however, while low amounts of zinc are required for growth of S. pneumoniae as well,18 high zinc concentrations are toxic for this micro-organism due to competition with manganese for the binding to pneumococcal surface adhesin A (PsaA), the solute-binding protein of a specific manganese transporter, thereby preventing manganese uptake.14–17
Our group previously reported elevated plasma MRP8/14 levels in patients with severe pneumonia.19 Here, we speculated that MRP8/14 is released at sites of infection, and that in pneumonia caused by S. pneumoniae, MRP8/14 may significantly impact on host defence, at least in part, by interfering with the availability of zinc. Therefore, we set out to determine the role of MRP8/14 in CAP and pneumococcal pneumonia by combining observational studies in humans with functional studies in mice.
Methods
Human studies
Two groups of consecutive CAP patients were studied.1 Eight patients admitted to the intensive care unit (ICU) of St Luc University Hospital (Leuven, Belgium), from whom bronchoalveolar lavage fluid (BALF) was obtained from the site of infection as visualised on the chest X-ray2; 7 patients admitted to the emergency department (ED) of St Antonius Hospital (Nieuwegein, The Netherlands), from whom BALF was obtained from both the infected and contralateral uninfected lung, as well as serum on admission and 1, 2, 5, 10 and 30 days thereafter. Twenty-four patients without lung disease scheduled to undergo elective surgery in whom BALF was collected directly after induction of anaesthesia served as a control.20 Additionally, we analysed BALF samples from a previous study21 in which eight healthy subjects (mean age 23.4±1.3 years) who were challenged with 100 ng/kg lipoteichoic acid (LTA), from Staphylococcus aureus (Deutsche Sammlung von Mikroorganismen 20233)22 into a lung subsegment and sterile saline into the contralateral lung.21 Bilateral BAL was performed 6 h thereafter. The institutional ethics committees approved all human studies and written informed consent was obtained from all patients or their relatives. MRP8/14 was measured by ELISA.23 Staining of MRP8 and MRP14 of archival postmortem lung tissue from patients who had died from pneumococcal pneumonia (n=3) or from a non-pulmonary cause (n=3) was done as described.24
Mouse studies
The Animal Care and Use Committee of the University of Amsterdam approved all experiments. C57Bl/6 wildtype (wt) mice (Charles River Laboratories, Maastricht, The Netherlands) and MRP14 deficient (Mrp14−/−) mice24 ,25 (backcrossed >10 times to a C57Bl/6-background) were intranasally inoculated with 5×103 or 5×104 colony forming units (cfu) S. pneumoniae serotype 3 (ATCC 6303 Rockville, Maryland, USA) (n=7 –8 per strain) and sacrificed 6, 24 or 48 h thereafter. Collection and handling of samples, and quantitative cultures were done as described.26 ,27 In survival studies, mice (n=12–16 per strain) were monitored for up to 7 days after infection (5×104 cfu). BALF was obtained from a separate group of infected wt mice (n=6) at indicated time points as described.26 ,27 See online supplement for cytokine measurements and histology.
In vitro studies
See online supplement.
Statistical analysis
Data are expressed as box-and-whisker diagrams unless indicated otherwise. Differences within individual patients or volunteers were analysed by Wilcoxon-matched pairs test. Differences between patients and volunteers, or wt and mrp14−/− mice were analysed by Mann–Whitney U test. Survival was compared by Kaplan–Meier analysis followed by a log rank test. Values of p<0.05 were considered statistically significant.
Results
Patients with CAP show elevated lung and plasma MRP8/14 levels
To obtain a first insight into MRP8/14 release during pneumonia, we measured MRP8/14 levels in BALF from eight CAP patients requiring treatment in the ICU, and from seven CAP patients admitted to the ED (table 1). S. pneumoniae was the most commonly isolated pathogen (in total 7 out of 15 cases, or 46.7%). Patients admitted to the ICU displayed elevated levels of MRP8/14 in BALF compared with controls (figure 1A, p<0.001). In agreement, patients admitted to the ED displayed a profound compartmentalised increase of MRP8/14 at the site of infection (figure 1B, p<0.001 vs the contralateral lung); these patients also showed raised serum MRP8/14 levels, which remained elevated until 30 days after admission compared with normal serum values (figure 1C, p<0.05; at other time points: p<0.01). To obtain insight in MRP8/14 expression in lung tissue in human CAP, we performed immunohistochemical staining for MRP8 and MRP14 on stored lung tissue slides of patients who had succumbed to CAP caused by S. pneumoniae and from patients who had died from a non-pulmonary cause. While control lung tissue samples showed a small number of resident cells expressing MRP8 or MRP14, lung tissue from patients who had died from pneumococcal pneumonia demonstrated a strong increase in both MRP8 and MRP14 staining (figure 1D). Together, these data provide strong evidence for local release of MRP8/14 during human CAP at the site of infection.
Demographic and clinical characteristics, and microbiology results of patients with community-acquired pneumonia
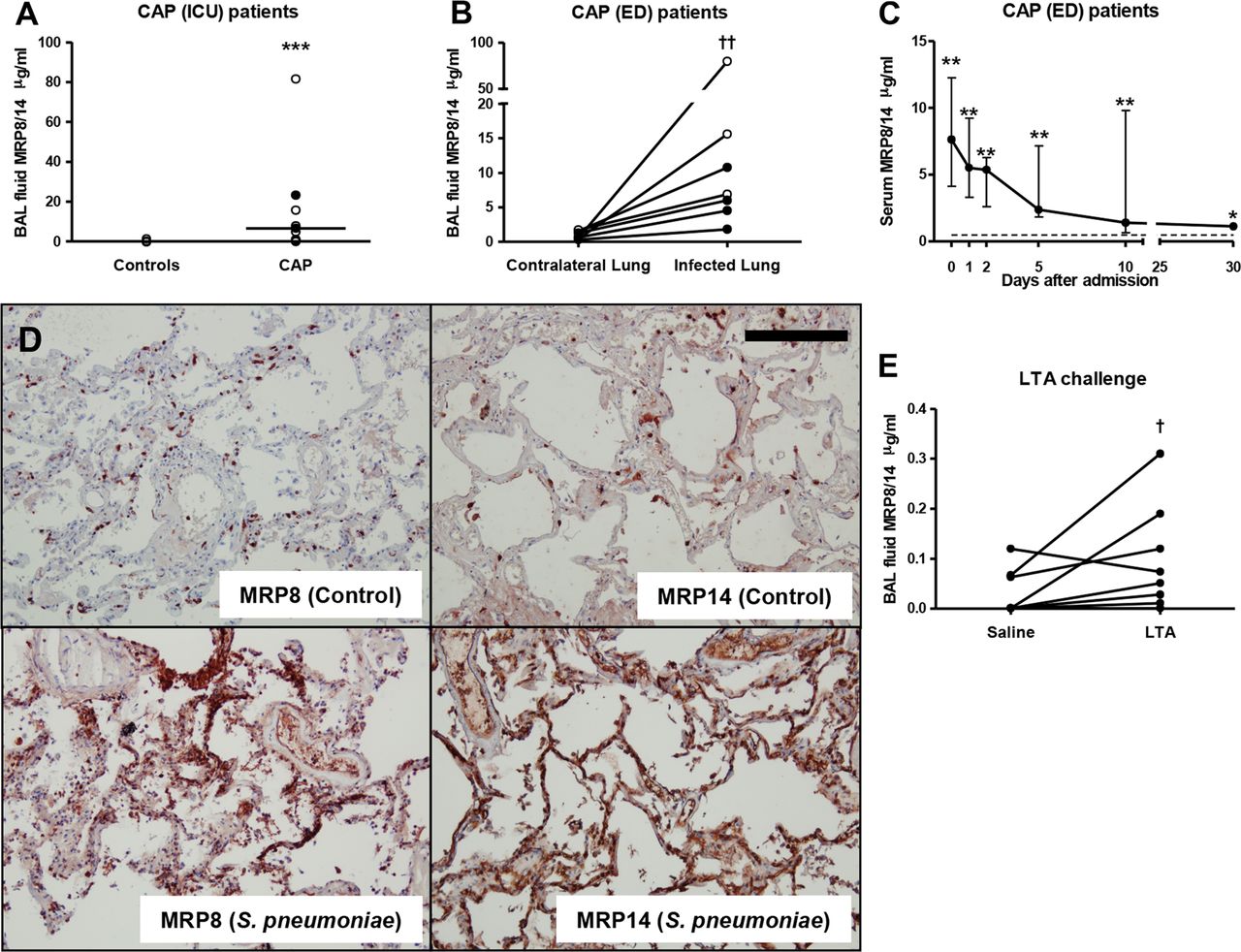
Human community-acquired pneumonia (CAP) and bronchial lipoteichoic acid (LTA) instillation result in an increase of myeloid-related protein (MRP)8/14 in lungs and plasma. (A) MRP8/14 levels in bronchoalveolar lavage fluid (BALF) from control patients and patients admitted to the intensive care unit with CAP. Each dot represents a measurement within an individual patient, with horizontal lines showing medians. Closed dots: Streptococcus pneumoniae pneumonia. Open dots: pneumonia due to other causative pathogen. (B) MRP8/14 levels in BALF from the infected and contralateral non-infected lung from patients admitted to the emergency department (ED) with CAP. Data are expressed as individual measurements (dots). Closed dots: S. pneumoniae pneumonia. Open dots: pneumonia due to other causative pathogen. Connections between dots express measurements from individual patients. (C) Serum MRP8/14 levels in serum from CAP patients admitted to the ED at indicated time points; the broken line represents the normal MRP8/14 value in a healthy individual measured at one timepoint. Data are expressed as median and IQR range. (D) MRP8 and MRP14 stainings in lungs of patients who had died from pneumococcal pneumonia or a non-pulmonary cause. Scalebar indicates 200 μm. (E) MRP8/14 levels in BALF from healthy subjects 6 h after intrabronchial instillation of LTA (100 ng/kg) or sterile saline (in the contralateral lung). Data are expressed as individual measurements (dots). Connections between dots express measurements from individual subjects; *p<0.05, **p<0.01, ***p<0.001 versus control BALF or serum, Mann–Whitney U test; †p<0.05, ††p<0.01 versus contralateral lung, Wilcoxon matched pairs test.
Next, we determined whether an acute bacterial stimulus can elicit local MRP8/14 release in the bronchoalveolar space of healthy humans. Indeed, 6 h after bronchial instillation, MRP8/14 concentrations were higher in BALF harvested from the lung segment challenged with LTA (a proinflammatory component of the Gram-positive cell wall) compared with BALF obtained from the saline challenged lung segment (p<0.05 vs saline, figure 1E).
Lung and plasma MRP8/14 levels are increased during murine pneumococcal pneumonia
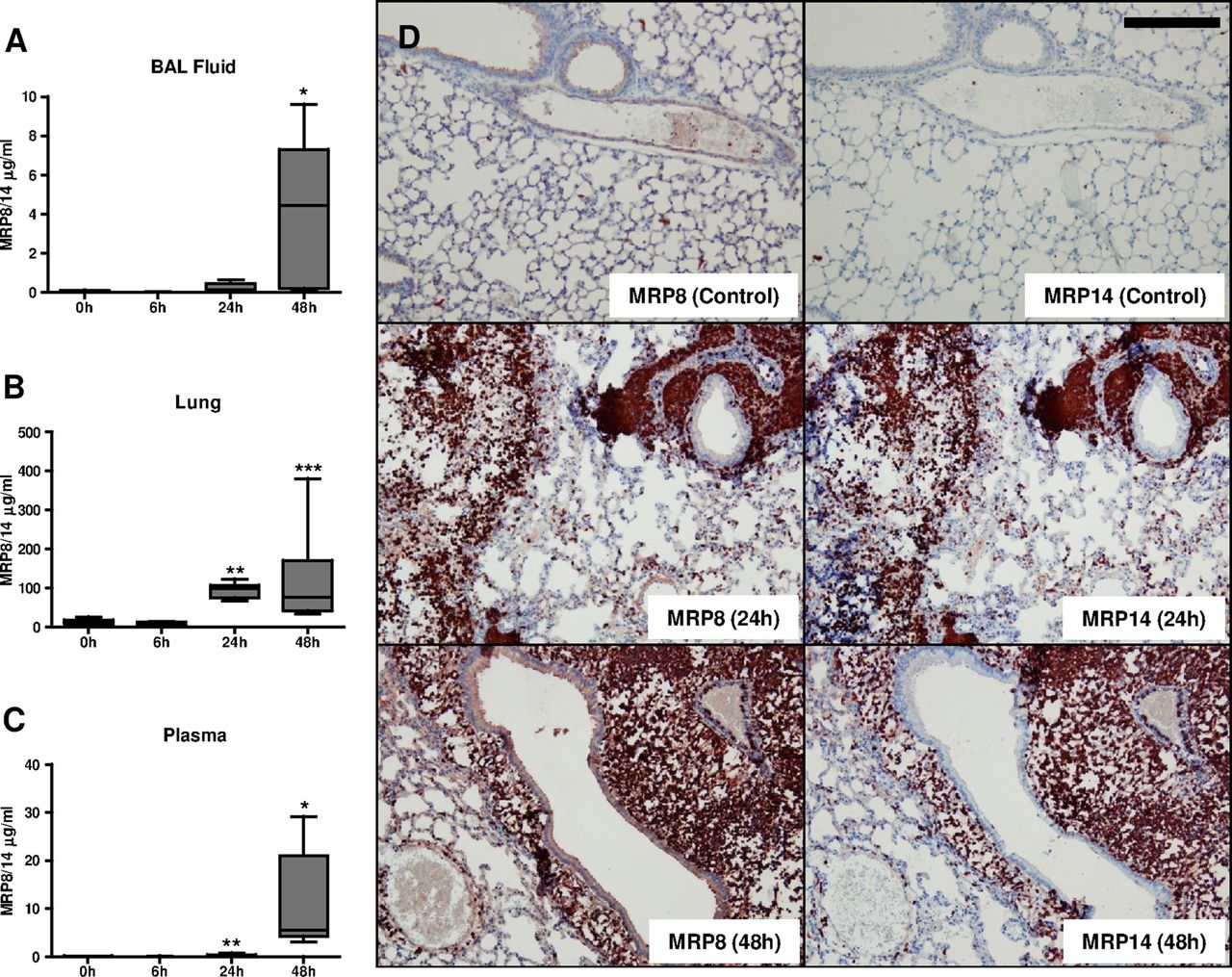
We used our well-established CAP model in mice, produced by intranasal infection with a virulent strain of S. pneumoniae26 ,27 to obtain insight into the functional role of MRP8/14. After pneumococcal infection (5×104 cfu), elevated local levels of MRP8/14 were measured in BALF (median 4.4 μg/mL at 48 h; figure 2A) and whole-lung homogenates (median 77.0 μg/mL at 48 h; figure 2B). In plasma, MRP8/14 was strongly enhanced after 48 h (median 5.6 μg/mL; figure 2C). We next stained lung tissue slides from naive and infected mice to establish the cellular source of MRP8 and MRP14 (figure 2D). Naive lungs showed a small number of MRP8 and MRP14 positive cells. MRP8 and MRP14 expressions both strongly increased at later time points, presumably due to large peribronchial neutrophilic infiltrates (see online supplementary figure S1). Together, these data indicate that murine pneumococcal pneumonia resembles human CAP with regard to high pulmonary and systemic expression of MRP8/14.

Murine pneumococcal pneumonia results in an increase of myeloid-related protein (MRP)8/14 in lungs and plasma. MRP8/14 levels in bronchoalveolar lavage fluid (A), whole-lung homogenates (B), and plasma (C) in naive mice and 6, 24 and 48 h after intranasal Streptococcus pneumoniae infection (5×104 cfu). Data are expressed as box-and-whisker diagrams depicting the smallest observation, lower quartile, median, upper quartile and largest observation. MRP8 and MRP14 stainings in lungs of naive mice and mice 24 h or 48 h after infection with S. pneumoniae (D). Scale bar indicates 200 μm; *p<0.05, **p<0.01, ***p<0.001 versus uninfected mice.
Mrp14−/− mice show delayed mortality and reduced bacterial growth and dissemination during pneumococcal pneumonia
To investigate a potential role for MRP8/14 in the outcome of pneumococcal pneumonia, we intranasally infected wt and mrp14−/− mice with S. pneumoniae (5×104 cfu) and monitored them for 7 days (figure 3A). Of note, mrp14−/− mice are functionally deficient for MRP8/14 as they lack MRP8 on a protein level, probably due to a higher turnover of isolated MRP8 in the absence of its binding partner MRP14.24 ,25 Remarkably, lack of MRP8/14 significantly prolonged survival: median survival times were 2.5 days for wt mice versus 3.7 days for mrp14−/− mice (p=0.005).

Mrp14−/− mice display diminished bacterial outgrowth and reduced mortality during pneumococcal pneumosepis. Survival of wildtype (wt) and mrp14−/− mice after intranasal infection with Streptococcus pneumoniae (5×104 cfu) (n=15 per group) (A). Bacterial loads in the lung (B), blood (C), spleen (D) and liver (E) after intranasal inoculation of 5×104 cfu S. pneumoniae in wt (grey) and mrp14−/− mice (white). In an additional experiment, we inoculated mice with 5×103 cfu S. pneumoniae (indicated as LD=lower dose). Data are expressed as box-and-whisker diagrams depicting the smallest observation, lower quartile, median, upper quartile and largest observation (8 mice per group at each time point); *p<0.05, **p<0.01 versus wt mice at the same time point.
We wondered whether the delayed lethality of mrp14−/− mice was associated with a difference in bacterial growth and dissemination. Therefore, we euthanased wt and mrp14−/− mice at 6, 24 or 48 h after pneumococcal infection (5×104 cfu), and examined bacterial loads in lungs, blood, spleens and livers (figure 3B–E). At 6 h, bacterial loads in the lungs of mrp14−/− mice were similar to those in wt mice. Twenty-four hours after infection, mrp14−/− mice displayed reduced bacterial outgrowth in all organs and blood (p<0.05 vs wt mice). After 48 h, mrp14−/− mice showed no difference in pulmonary bacterial loads compared to wt mice, but still lower pneumococcal burdens in blood and spleen (p<0.05). Similar results were obtained with a 10-fold lower dose of pneumococci (5×103 cfu): 48 h after infection, mrp14−/− mice had similar bacterial loads in their lungs and lower bacterial burdens in blood and distant organs relative to wt mice (p<0.01). Together, these data indicate that MRP8/14 facilitates bacterial growth in lungs and especially distant organs, thereby contributing to accelerated lethality during pneumococcal pneumonia-derived sepsis.
MRP14 deficiency has little impact on lung inflammation
To evaluate the role of MRP14 in lung inflammation we analysed lung tissue slides obtained from wt and mrp14−/− mice at different time points after pneumococcal infection. Lung inflammation increased in time to a similar extent in wt and mrp14−/− mice, revealing pathology consistent with severe pneumonia in both mouse strains (figure 4A–B).

Pulmonary inflammation during pneumococcal pneumonia is not affected by loss of myeloid-related protein (MRP)8/14. Total pathology score at indicated time points after infection with Streptococcus pneumoniae (5×104 cfu) in wildtype (wt) (grey) and mrp14−/− mice (white) determined according to the scoring system described in the online supplement (A). Data are expressed as box-and-whisker diagrams depicting the smallest observation, lower quartile, median, upper quartile and largest observation (8 mice per group at each time point). There were no statistically significant differences between the groups. Representative slides of lung H&E stainings of wt and mrp14−/− mice 48 h after intranasal S. pneumoniae infection (B). Scale bar indicates 200 μm. Pulmonary MPO, indicative for the number of neutrophils (C), chemokine (KC and MIP-2) (D–E) and cytokine (TNF-α, IL-6 and IL-10) (F–H) 6, 24 and 48 h after intranasal S. pneumoniae infection in wt (grey) and mrp14−/− mice (white). Data are expressed as box-and-whisker diagrams depicting the smallest observation, lower quartile, median, upper quartile and largest observation (8 mice per group at each time point); **p<0.01 versus wt mice at the same time point.
Several studies have implicated MRP8/14 as a mediator of neutrophil recruitment.28–30 We determined myeloperoxidase (MPO) levels in whole-lung homogenates as a measure of neutrophil numbers (figure 4C). At 6 h, lung MPO levels were low in both groups, but slightly higher in mrp14−/− mice (p<0.01 vs wt mice); MPO concentrations were much higher at later time points and not different between groups.
As an additional readout for lung inflammation, we measured chemokines (MIP-2, KC) and cytokines (TNF-α, IL-6, IL-10) in whole lung homogenates. Pulmonary cytokine and chemokine levels increased similarly in both mouse strains during the course of the disease, showing no differences between mrp14−/− and wt mice (figure 4D–H). Also, 48 h after infection with 5×103 cfu S. pneumoniae, lung chemokine and cytokine levels did not differ between groups (data not shown). Altogether, these data indicate that MRP14 deficiency has little if any impact on the extent of lung inflammation induced by pneumococci.
Impact of MRP14 deficiency on plasma cytokine levels
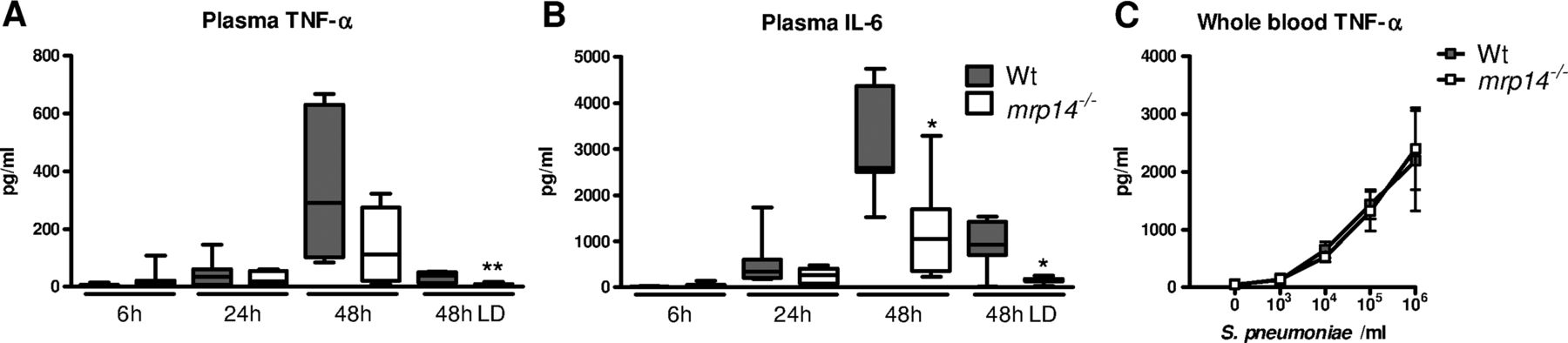
We next determined the role of MRP8/14 in systemic inflammation. Mrp14−/− mice showed lower plasma IL-6 concentrations at 48 h after infection with 5×104 cfu S. pneumoniae and lower plasma IL-6 and TNF-α levels after administration of 5×103 cfu pneumococci (p<0.01 and 0.05 vs wt mice) (figure 5A, B). As these lower cytokine levels could be a consequence of lower bacterial loads in the blood of mrp14−/− mice, we harvested whole blood from wt and mrp14−/− mice which was stimulated with increasing concentrations of growth-arrested S. pneumoniae. Wt and mrp14−/− whole blood produced similar TNF-α levels (figure 5C), suggesting that the lower systemic cytokine levels in mrp14−/− mice might be a reflection of the lower bacterial numbers.
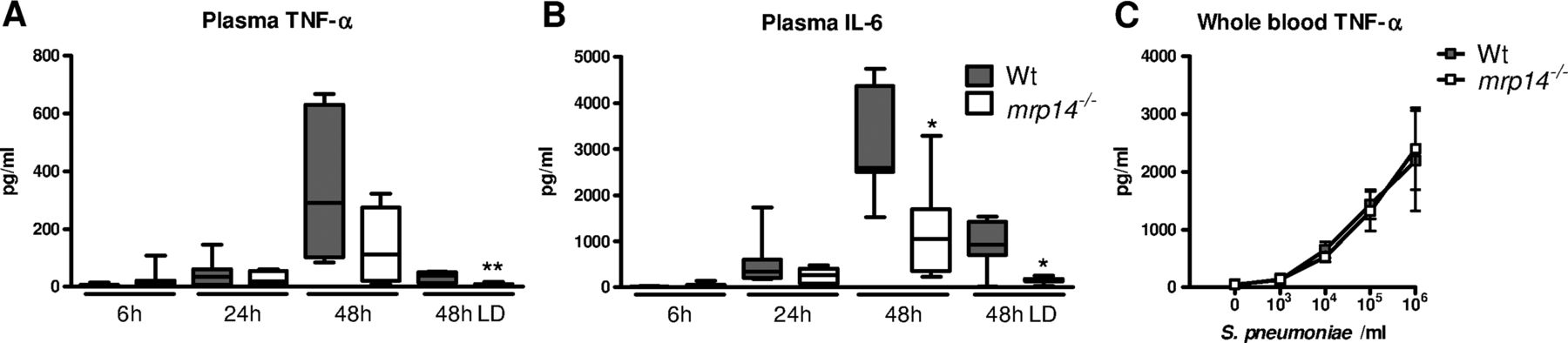
Plasma cytokine levels during pneumococcal pneumonia-derived sepsis. Plasma TNF-α (A) and IL-6 (B) 6, 24 and 48 h after intranasal Streptococcus pneumoniae infection (5×104 cfu) in wildtype (wt) (grey) and mrp14−/− mice (white). Data are expressed as box-and-whisker diagrams depicting the smallest observation, lower quartile, median, upper quartile and largest observation (8 mice per group at each time point). LD: lower dose (5×103 cfu); *p<0.05, **p<0.01 versus wt mice at the same time point. TNF-α levels after a 6 h stimulation of whole blood obtained from individual wt and mrp14−/− mice (n=4 per group) with log-increasing concentrations of growth-arrested S. pneumoniae (C). Data are expressed as means±SEM (4 mice per group).
MRP8/14 enhances pneumococcal growth by reducing zinc toxicity
MRP8/14 can bind zinc and manganese, which results in growth inhibition of several micro-organisms.8–12 ,31 ,32 While pneumococci also require zinc and manganese for growth, higher zinc concentrations competitively inhibit manganese uptake, rendering them hypersensitive to oxidative stress.14–17 Of interest, a previous study demonstrated an increase of serum zinc in murine pneumococcal pneumonia from approximately 15 to 640 μM. Manganese concentrations remained stable (0.4–0.7 μM).17 As MRP8/14 contains two metal-binding sites with high affinity for zinc, whereas, only one binds manganese,31 ,32 we hypothesised that metal binding by this heterodimer might be advantageous for the pathogen rather than for the host. We confirmed16 ,17 that increasing zinc concentrations (up to 200 µM) progressively inhibited pneumococcal growth in vitro in the presence of a constant manganese concentration (0.1 µM) (figure 6A). By comparison, zinc did not influence the growth of Klebsiella pneumoniae in vitro under these conditions (figure 6B), suggesting that pneumococci indeed have a relative susceptibility towards a high zinc/manganese ratio. Importantly, recombinant mouse MRP8 or MRP8/14, but not MRP14, reduced (100 µM) zinc-induced toxicity toward pneumococci (figure 6C). Without zinc addition, MRP8/14 (100 µg/mL) inhibited pneumococcal growth (figure 6D), mimicking its effect on other bacterial species.8–12 Together these data suggest that, in contrast with the antimicrobial effects exerted by the metal chelating properties of MRP8/14 on other bacterial species, the zinc-binding capacity of MRP8/14 is of benefit to S. pneumoniae.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Myeloid-related protein (MRP)8/14 reduces zinc mediated toxicity towards pneumococci. Growth of Streptococcus pneumoniae (A) and Klebsiella pneumoniae (B) was assessed for 6 and 20 h in the presence of 0.1 µM manganese and increasing concentrations of zinc. The growth-inhibiting effect of zinc towards pneumococci was reduced by increasing concentrations of MRP8 and MRP8/14 (C). MRP8/14 inhibits S. pneumoniae growth in zinc-free medium (D). Data are means±SEs of at least four replicates and representatives of triplicate experiments.
Discussion
Invasive infection is associated with the release of MRP8/14, which serves a complex role in a variety of innate defence mechanisms. MRP8/14 attracts neutrophils,28–30 mediates inflammatory processes via TLR4,7 and binds metals, which is antimicrobial against a range of bacterial8–10 and fungal11 ,12 pathogens. Here, we observed elevated MRP8/14 levels in the course of clinical and experimental Gram-positive pneumonia. In sharp contrast with earlier studies on the role of MRP8/14 in host defence against infection,8–12 we found that the presence of MRP8/14 was of benefit to S. pneumoniae rather than to the host, as reflected by reduced bacterial dissemination and delayed lethality of mrp14−/− mice in lethal pneumonia-derived sepsis. The mechanism by which S. pneumoniae exploits MRP8/14 likely involves the zinc chelating capacity of this heterodimer, thereby reducing zinc-mediated toxicity towards the pneumococcus.
We previously reported elevated MRP8/14 serum levels in patients with sepsis originating from different sources, including pneumonia.19 Accordingly, we demonstrate elevated serum MRP8/14 levels in newly admitted CAP patients. MRP8/14 levels were notably enhanced at the primary site of infection, as detected in BALF and lung tissue slides, corresponding to high levels in the abdominal cavity during peritoneal sepsis.19 Moreover, compartmentalised release of MRP8/14 was detected in healthy humans challenged with LTA in a lung subsegment. Together these data indicate that MRP8/14 is highly produced at the site of infection and released systemically during human pneumococcal pneumonia. Similarly, experimentally induced pneumococcal pneumonia resulted in high local and systemic release of MRP8/14 in mice, which is in agreement with earlier studies of murine bacterial pneumonia.9 ,30
Although MRP8/14 may exhibit proinflammatory effects under various pathophysiologic conditions,7 ,19 we were unable to demonstrate such a role during pneumonia. We observed largely unaltered pulmonary cytokine responses and lung pathology. The reduced systemic cytokine levels in the current study were most likely caused by lower bacterial burdens in the circulation, as whole blood from mrp14−/− mice produced similar TNF-α levels as wt blood upon stimulation with growth-arrested pneumococci. Previous studies have shown that MRP8/14 is able to potentiate lipopolysaccharide-induced signalling via TLR4, while by itself exerting only minimal proinflammatory effects.7 Although S. pneumoniae may activate TLR4 via its virulence factor pneumolysin,33 TLR2 is the predominant pattern recognition receptor that mediates lung inflammation induced by this pathogen.34 ,35 Moreover, we previously reported that MRP8/14 does not contribute to lung inflammation during Gram-negative (Klebsiella) pneumonia9 wherein TLR4 is the main receptor driving the inflammatory response.36 ,37 Together, these data indicate that endogenous MRP8/14 does not amplify inflammation in the lungs during pneumonia caused by a gradually growing bacterial load, that is, in the context of a likely clinical scenario.9
Many previous investigations have established an antibacterial role for MRP8/14 in in vivo infection models,8–12 which is dependent on the chelation of both zinc and manganese.31 ,32 Of note, MRP8 and MRP14 homodimers only bind zinc at their His-Glu-Xaa-Xaa-His (HEXXH) zinc-binding amino acid motif and do not impair microbial growth.38 When infected with pyogenic bacteria, such as S. aureus,8 K. pneumoniae9 or Acinetobacter baumanii,10 mrp14−/− mice displayed enhanced bacterial outgrowth or mortality. In contrast to these earlier findings, lack of MRP8/14 reduced bacterial burdens in the lung, blood and distant organs and improved survival in pneumococcal pneumonia derived sepsis. This apparent inconsistent finding seemed surprising at first glance, especially since pneumococci, like the pathogens listed above, critically requires both zinc and manganese for normal (aerobic) growth.14–17 Higher zinc concentrations however, during pneumococcal infection,17 have significant toxicity towards this bacterium.16 ,17 Zinc irreversibly binds to PsaA, thereby blocking intracellular uptake of manganese. Uptake of manganese is essential for manganese superoxide dismutase to render pneumococci less susceptible to oxidative stress and polymorphonuclear leukocyte killing.16 ,17 We postulate that high MRP8/14 concentrations reduce zinc/manganese ratios at infected body sites during pneumococcal pneumonia-derived sepsis, thereby providing a benefit for the pathogen rather than for the host. Previous investigations have indeed shown that MRP8/14 has two high-affinity metal binding sites for zinc, whereas only one is able to bind manganese.31 ,32 In our in vitro experiments, addition of recombinant MRP8/14 and MRP8 (most likely through zinc chelation at its HEXXH zinc-binding site38), but not MRP14 partially restored growth of pneumococci in a high zinc/manganese ratio environment, revealing a likely mechanism for enhanced bacterial growth and, consequently, higher lethality in mrp14−/− mice during pneumococcal sepsis. Such a mechanism would also explain the opposite phenotypes of mrp14−/− mice during pneumonia caused by S. pneumoniae (reported here) and K. pneumoniae,9 since Klebsiella clearly proved not susceptible to high zinc concentrations. Of interest, very recently, Neisseria meningitidis was also reported to misuse MRP8/14 to promote its growth in vitro, albeit by a different mechanism, namely by binding MRP8/14 and using this protein as a zinc source in low zinc culture conditions.39
A limitation of our study is that we were not able to directly measure zinc and manganese ratios at tissue level, at sites infected by pneumococci, as the state-of-the-art technique to measure localised metal distribution in tissues, that is, laser ablation inductively coupled plasma mass spectrometry, requires visible inflammatory lesions,8 which is not a pathologic feature of pneumococcal infection. Furthermore, to establish the role of MRP8/14 in the reduction of zinc toxicity and enhanced pneumococcal growth, additional studies should be conducted, such as supplementing mice with MRPs and/or feeding them with low or high zinc diets.
In conclusion, we document that human CAP is associated with profoundly elevated MRP8/14 concentrations in lungs and the circulation. Using our model of murine pneumococcal pneumonia, we show that the presence of high MRP8/14 levels is associated with enhanced bacterial outgrowth and lethality, most likely through binding of zinc by MRP8/14 and, thereby, reduction of zinc-mediated toxicity towards the bacterium. This study shows for the first time that the elemental binding properties of MRP8/14 can be misused by a clinical-relevant pathogen.
Acknowledgments
The authors thank Marieke ten Brink and Joost Daalhuisen for their expert technical assistance during the animal experiments, and Regina de Beer for performing histopathological and immunohistochemical stainings.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
-
Contributors AA performed the study, collected, analysed and interpreted the data and wrote the first draft of the article. TV, HE, BLM, MADvZ, YZ, JJH and SF assisted in collection and interpretation of the data. P-FL, XW, MJS, JCG and DHB provided human samples. JR, EPS, CvtV and AFdV assisted in interpretation of the data. TvdP designed the study and assisted in interpretation of the data. All authors contributed in revising the manuscript and finally approved the version of the manuscript to be published.
-
Funding This work is supported by a grant from the Landsteiner Foundation for Blood Transfusion Research (project LSBR 0706) and a clinical research talent fellowship of the University Medical Center Utrecht (2014 to MADvZ).
-
Competing interests None.
-
Ethics approval Scientific and ethics committees of the Academic Medical Center (Amsterdam, The Netherlands), St. Luke University Hospital (Brussels, Belgium), and/or St. Antonius Hospital (Nieuwegein, The Netherlands.
-
Provenance and peer review Not commissioned; externally peer reviewed.