Article Text

Abstract
Rationale Mutations in the cystic fibrosis transmembrane regulator (CFTR) gene form the basis of cystic fibrosis (CF). There remains an important knowledge gap in CF as to how diminished CFTR activity leads to the dominant inflammatory response within CF airways.
Objectives To investigate if extracellular vesicles (EVs) contribute to inflammatory signalling in CF.
Methods EVs released from CFBE41o-, CuFi-5, 16HBE14o- and NuLi-1 cells were characterised by nanoparticle tracking analysis (NTA). EVs isolated from bronchoalveolar lavage fluid (BALF) from 30 people with CF (PWCF) were analysed by NTA and mass spectrometry and compared with controls. Neutrophils were isolated from the blood of 8 PWCF to examine neutrophil migration in the presence of CFBE41o- EVs.
Results A significantly higher level of EVs were released from CFBE41o- (p<0.0001) and CuFi-5 (p=0.0209) relative to control cell lines. A significantly higher level of EVs were detected in BALF of PWCF, in three different age groups relative to controls (p=0.01, 0.001, 0.002). A significantly lower level of EVs were released from CFBE41o- (p<0.001) and CuFi-5 (p=0.0002) cell lines treated with CFTR modulators. Significant changes in the protein expression of 126 unique proteins was determined in EVs obtained from the BALF of PWCF of different age groups (p<0.001–0.05). A significant increase in chemotaxis of neutrophils derived from PWCF was observed in the presence of CFBE41o EVs (p=0.0024) compared with controls.
Conclusion This study demonstrates that EVs are produced in CF airway cells, have differential protein expression at different ages and drive neutrophil recruitment in CF.
- cystic fibrosis
- paediatric lung disaese
- neutrophil biology
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is the key question?
To investigate if airway extracellular vesicles (EVs) contribute to inflammatory signalling in cystic fibrosis (CF).
What is the bottom line?
This pilot study provides several novel insights on airway EVs in CF.
Why read on?
This study demonstrates greater EV release in CF airways compared with controls and connects CFTRto EV release in epithelial airway cells. The study demonstrates unique protein fingerprints in bronchoalveolar lavage fluid EVs at different ages illustrating a potential for EVs as biomarkers. The study provides information on how CF airway cells may contribute to neutrophil activation through the release of EVs.
Introduction
Cystic fibrosis (CF) is a progressive genetic disease that causes persistent airway infection ultimately leading to respiratory failure and early death. It is caused by functional deficiency of the cystic fibrosis transmembrane regulator (CFTR) gene with the most common genetic mutation, Phe508del (also known as F508 del), occurring in 70% of people with CF (PWCF). Mutations in the CFTR gene form the basis for the clinical manifestations of CF which affects epithelial innate immune function and airway clearance in the lung. Epithelial dysfunction has also been implicated in modulating chronic inflammation in CF airways, resulting in the initiation of a proinflammatory cascade that leads to the recruitment of leucocytes into the airway. Excessive neutrophil recruitment to the lung underlies inflammatory-mediated lung damage in CF.1
There still remains an important knowledge gap in CF as to how diminished CFTR activity leads to the dominant inflammatory response that causes tissue destruction and eventual challenges to organ function. We propose to examine if CF inflammatory cell function can be regulated by extracellular vesicles (EVs) released from lung epithelial cells with mutated CFTR. Exosomes (50–150 nm) are small vesicles actively secreted by most living cells, that contain both proteins involved in exocytosis and signalling within the microenvironment.2 A role for small vesicles regulating inflammatory cell recruitment has recently emerged in airway diseases. Macrophage-derived exosomes dampened the inflammatory response in the lung by inhibition of proinflammatory STAT activation in alveolar epithelial cells.3 Similarly, a role for exosomes in mediating inflammation and contributing to disease pathogenesis has also been reported in asthma and COPD.4 5 Other vesicles such as small micropatricles/ectosomes (100–500 nm)6 and exomeres (30–70 nm)7 have also been reported to be biologically significant in disease states. A recent study identified proinflammatory signatures in respiratory vesicles from a small number of PWCF8 highlighting the potential of EVs in modulating inflammation in CF.9
We hypothesised that EVs released from CF lung epithelial cells play a role in regulating leukocyte migration into the CF airways and may help understand disease pathogenesis in CF. In this study, we employed comparative proteomics to characterise the global activity and protein composition of EV’s released from CF airway cell lines and present in the bronchoalveolar lavage fluid (BALF) from PWCF. We functionally examined the role of epithelial derived EVs in modulating neutrophil chemotaxis and degranulation and explored the role of the S100 A12/RAGE pathways as potential modulator of EV regulated chemotaxis. Additionally, we analysed EVs in the presence of current CFTR correctors to investigate the relationship between CFTR dysfunction/rescue and EV release.
Materials and methods
All detailed materials and methods including descriptions of immunoblotting, flow cytometry, electron microscopy, neutrophil transmigration and protein pathway analysis are available in online supplementary materials.
Supplemental material
Subjects
BALF and blood were obtained through The Study of Host Immunity and Early Lung Disease in Cystic Fibrosis (SHIELD-CF) from children with CF and from adults with CF attending St. Vincent’s University Hospital (SVUH). Control samples were obtained from SHIELD-CF and Tallaght University Hospital (TUH). Subject characteristics are listed in online supplementary table 1. Non-CF control samples were obtained from children and adults without CF presenting with recurrent respiratory infection.
Supplemental material
Cell culture
HBE41o- and NuLi-1 expressing wild-type (WT) CFTR, CFBE41o- and CuFi-5 cells homozygous for F508del-CFTR, CuFi-4 heterozygous for F508del-CFTR and G551D were cultured in medium as previously described.10 11
SiRNA transfections
CFBE41o- cells were transfected with 5 nM siRNA (Invitrogen, Grand Island, New York) as described previously.10 Target siRNAs were to S100 A12 (assay ID s12433, Ambion).
Collection and processing of BALF
BALF was collected and processed as previously described12 in CHI at Crumlin, SVUH and TUH.
EV fraction Isolation
EV fractions were isolated by differential ultracentrifugation. Briefly, cellular supernatants or BALF samples were centrifuged at 3000g for 30 min to remove cell debris followed by 10 000g centrifugation for 30 min at 4°C to remove large microparticles and 120 000 g for 2 hour at 4°C (XL-70 ultracentrifuge, Beckman-Coulter, Villpinte, France) to pellet the EVs.
Nanoparticle tracking analysis (NTA)
Particle size distribution in cellular supernatants and BALF samples was determined by NTA using a NanoSight NS300 system (Malvern Technologies, Malvern, UK) as previously described.13
Neutrophil isolation and migration
Neutrophils were isolated from whole blood as previously described.14
Neutrophil migration
Migration assays were performed using 12 mm Costar Transwell-COL collagen-coated 3.0 µM pore Polytetrafluoroethylene (PTFE) membrane inserts (Sigma Aldrich). Neutrophils were seeded (5 × 105 cell/mL) on the upper chamber. The lower chamber was filled with RPMI medium or supplemented with HBE41o-/CFBE41o- EVs (10 µg) or fMLP (100 nM). After 3 hours, the number of migrated cells was counted under a microscope (10×). Data were counted and analysed using ImageJ (https://imagej.nih.gov/ij/).
Myeloperoxidase assay (MPO)
MPO release was measured from isolated neutrophils post chemotaxis as per manufacturing instructions (Sigma-Aldrich).
Mass spectrometry
EVs isolated from BALF from PWCF and CF bronchial epithelial cell lines were digested and analysed by mass spectrometry as previously described.10
Analysis of tandem mass spectra
The Q-exactive raw data files were de novo sequenced and cross searched against a Human UniProtKB database Proteome IDi UP000005640, November 2017 entries using MaxQuant V.1.5. Proteins were quantified by Label-free Quantification using MaxQuant software as previously described.15 Excel was employed to finally analyse the MaxQuant data, using fold change cut-off values of 1.5.
Statistical analysis
All quantified data are presented as the mean ± SEM for at least three independent experiments. Student paired and unpaired t-test analysis and analysis of variance (ANOVA) was conducted using Prism 6 (GraphPad Prism, San Diego, California, USA). One way ANOVA was used for statistical analysis involving three or more groups. Bonferroni multiple comparisons post-hoc test was used to compare multiple groups to one group. A summary table of all experiments and statistical tests are listed in table 1.
Summary table of experiments and statistical tests including samples number, sample type, associated figure code, statistical test and summary p values generated by analysis of variance (ANOVA) or unpaired t-tests
Results
CFTR expression in both CFBE41o- cells (figure 1A), and CuFi-5 cells (figure 1D) and their WT controls was confirmed by immunoblotting. The size and number of EVs released from CF bronchial cells lines, CFBE41o-/CuFi-5, were determined by NTA. A significantly higher concentration of EV particles were released from CFBE41o- cells (figure 1Bi, Bii; p<0.0001) and CuFi-5 (figure 1Ei,Eii; p=0.0209) compared with the WT controls. We observed a marked presence of vesicles between 50 and 250 nm (figure 1B and E, online supplementary figure S1) by NTA. Additionally, EV fractions were analysed by transmission electron microscopy (TEM) which detected vesicles in the 50–250 nm range (online supplementary figure S2). We confirmed an expression and enrichment of known vesicle markers (CD63, CD9) in EVs isolated from CFBE41o- and CuFi-5 cell lines (figure 1C,D; online supplementary figure S3). We also confirmed expression of additional exosomal markers annexin V/flotillin 1 in isolated EVs and an absence of ER secretory protein calnexin in the isolated EV fraction (online supplementary figure S3). These are markers recommended by International Society for Extracellular Vesicles (Minimal Information for the Study of Extracellular Vesicles- MISEV 2018 guidelines).16 We, therefore refer to this fraction as an EV fraction of the small EV subtype that potentially contains exosomes, small microparticles and exomeres.
Supplemental material
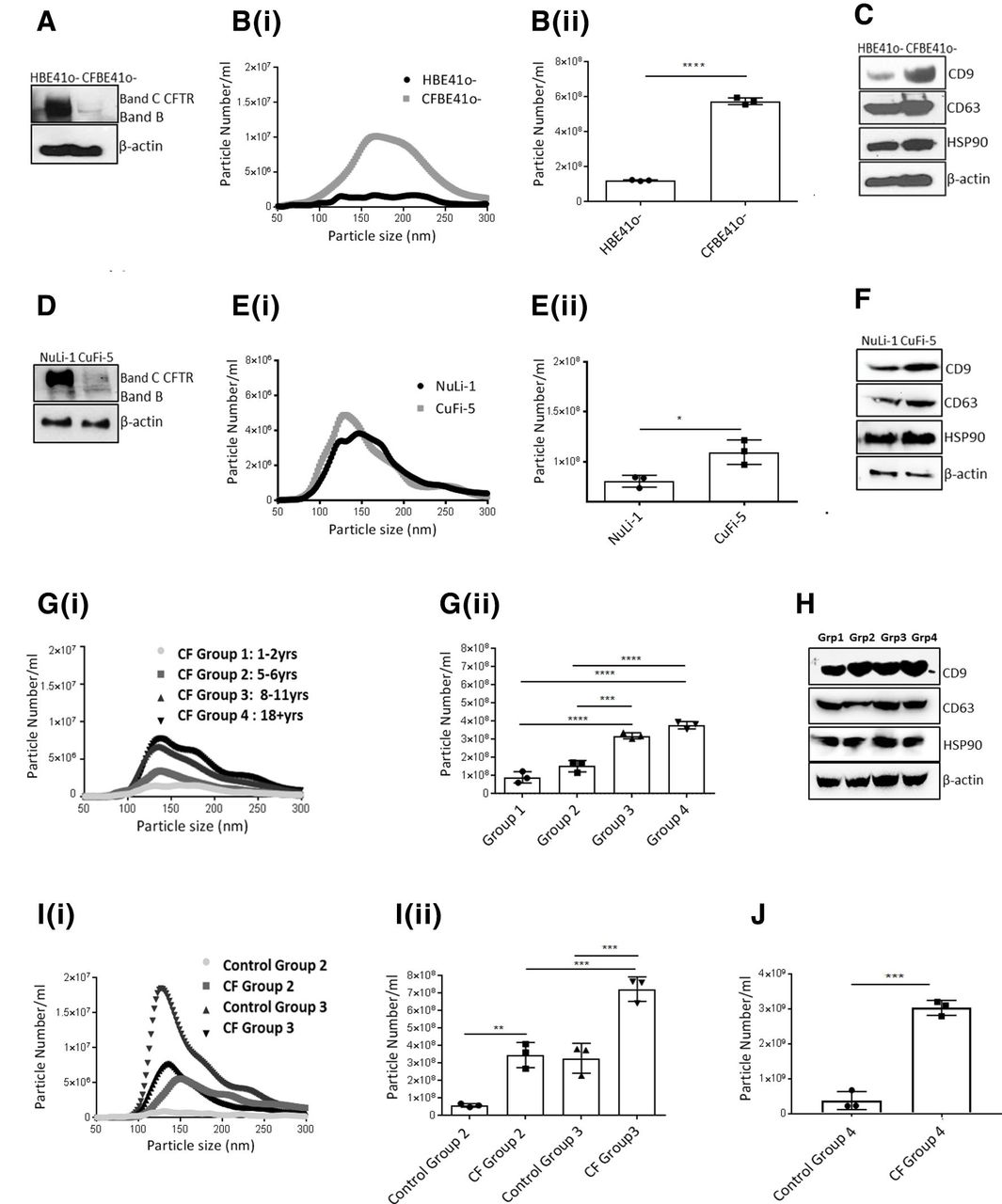
Extracellular vesicle (EV) release is upregulated in cystic fibrosis (CF) bronchial models and bronchoalveolar lavage fluid (BALF) from people with CF (PWCF). (A) Protein expression of CF transmembrane regulator (CFTR) was determined by immunoblotting in HBE41o- and CFBE41o- cells. (B) (i) Nanoparticle tracking analysis (NTA) of EV particles isolated from HBE41o- and CFBE41o- cells was performed in triplicate and (ii) quantified. Histograms represent the average of three experiments. Statistical analysis used was unpaired Student t-test, and error bars denote mean±SD, p<0.0001(****). (C) Protein expression of CD9, CD63 and HSP90 in EVs isolated from HBE41o- cells and CFBE41o- cells was determined by immunoblotting. (D) Protein expression of CFTR in NuLi-1 and CuFi-5 cells was determined by immunoblotting. (E) (i) NTA analysis of EV particles isolated from NuLi-1 and CuFi-5 cells was performed in triplicate and (ii) quantified. Histograms represent the average of three experiments. Statistical analysis used was unpaired Student t-test, and error bars denote mean±SD, p<0.05 *(p=0.0209). (F) Protein expression of CD9, CD63 and HSP90 in EVs isolated from NuLi-1 and CuFi-5 cells was determined by immunoblotting. (G) EVs were isolated from BALF of PWCF from four different age groups (group 1=1–2 years, group 2=5–6 years; group 3=8–12 years, group 4=18+ years) and (i) NTA analysis of particles was performed and (ii) particles were quantified. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.001*** (p=0.0003), p<0.0001****. *P values were generated by analysis of variance (ANOVA) test followed by Bonferroni adjustment. (H) Protein expression of CD9, CD63 and HSP90 in EVs isolated from BALF from patients with CF in (G) was determined by immunoblotting. (I) EVs were isolated from BALF of PWCF from two different age groups (group 2=5–6 years; group 3=8–11 years) and their age-matched control patients (non-CF) and (i) NTA analysis of particles was performed and (ii) particles were quantified. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.01** (p=0.0043) and p<0.001 *** (p=0.0005). *P values were generated by ANOVA test followed by Bonferroni adjustment. (J) EVs were isolated from BALF of PWCF and controls (group 4—adults). Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.001***(p=0.0002).
We isolated EVs from the BALF of PWCF homozygous for the F508del mutation. Subject demographics are included in online supplementary table 1. NTA analysis of BALF revealed an age-dependent increase in EVs (50–250 nm range) from PWCF in four different age groups (figure 1Gi) which was quantified in triplicate (figure 1Gii). The expression of markers, CD9 and CD63, was confirmed across all age groups by immunoblotting in EV fractions (figure 1H). We further investigated whether release of EVs differed between the BALF of PWCF and age-matched non-CF controls. NTA analysis revealed a significantly higher number of EVs (50–250 nm range) in children from group 2 (5–6 years; p=0.0043) and group 3 (8–11 years; p=0.0007) compared with their age-matched non-CF controls (figure 1Ii). This was quantified in triplicate (figure 1Iii). A significantly higher number of EVs was observed in group 4 compared with controls (p=0.0002; figure 1J). There was also an overall increase in EVs isolated from group 1 (1–2 years) compared with age-matched controls but this was not significant (p=0.0809; online supplementary figure S4).
CFBE41o- and CuFi-5 cells were treated with kinase inhibitors based on previous work.10. Expression of CFTR and autophagy marker LC3 was measured by immunoblotting. An increase in total CFTR expression was observed following treatment with three kinase inhibitors, AKT8 (5 µM), everolimus (5 µM) and MK-2206 (5 µM), in CFBE41o- (figure 2A) and CuFi-5 (figure 2C) cell lines. All three kinase inhibitors were were able to restore defective autophagy mechanism in CFBE41o- (figure 2A) and CuFi-5 (figure 2C) cells. AKT8, everolimus and MK-2206 inhibitors significantly reduced EV release in CFBE41o- cells (figure 2B; p=0.0003, p=0.0016, p=0.001) and CuFi-5 cells (figure 2D; p=0.001, p<0.0001). CFBE41o- and CuFi-5 cells were treated with lumacaftor (5 µM), tezacaftor (5 µM), ivacaftor (1 µM) or combinations of each. Restoration of autophagy and was observed (LC3) in CFBE41o- and CuFi-5 cells after drug treatments. An increase in CFTR stability was detected, particularly when combination treatments were used (figure 2E and G). We observed a significantly decreased release of EVs from both CFBE41o- (p<0.001) and CuFi-5 (p=0.0004, p=0.0015) cells following treatment with combinations of ivacaftor with lumacaftor and tezacaftor, respectively (figure 2F and H).

Extracellular vesicle (EV) release can be modulated by cystic fibrosis transmembrane regulator (CFTR) correctors and potentiators. (A) CFBE41o- cells were treated with 5 µM AKT-VIII, 5 µM everolimus and 5 µM MK-2206 and immunoblotting was performed for CFTR, LC3I/II and β-actin. (B) CFBE41o- cells were treated with AKT-VIII, everolimus and MK-2206 as above (A) and nanoparticle tracking analysis (NTA) was performed. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.001***(p=0.0003) and p<0.01** (p=0.0016, p=0.0010), respectively. (C) CuFi-5 cells were treated with 5 µM AKT-VIII, 5µM everolimus and 5 µM MK-2206 for 48 hours and immunoblotting was performed for CFTR, LC3I/II and β-actin. (D) CuFi-5 cells were treated with AKT-VIII, everolimus and MK-2206 as above (C) and NTA analysis was performed in triplicate. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.001***(p=0.0001) and p<0.0001 **** respectively. (E) CFBE41o- cells were treated with 5 µM lumacaftor, 5 µM tezacaftor, 1 µM ivacaftor alone or in combination with 1 µM ivacaftor for 24 hours and immunoblotting was performed for CFTR, LC3I/II and β-actin. (F) CFBE41o- cells were treated as above (E) with lumacaftor, tezacaftor alone or in combination with ivacaftor and NTA analysis was performed. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.0001**** for all treatments. (G) CuFi-5 cells were treated with 5 µM lumacaftor, 5 µM tezacaftor alone or in combination with 1 µM ivacaftor and immunoblotting was performed for CFTR, LC3I/II and β-actin. (H) CuFi-5 cells were treated with the same inhibitors as in (G) and NTA analysis was performed. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.001***(p=0.0004) and *p<0.01 (p=0.0015), respectively, for lumacaftor and tezacaftor combination treatments. All the above p values (A–H) were generated by analysis of variance test followed by Bonferroni adjustment. (I) CuFi-4 (G551D mutation) cells were treated with 10 µM ivacaftor and (i) NTA analysis of particles was performed and (ii) particles were quantified. Histograms represent the average of three experiments. Statistical analysis used was paired Student t-test, and error bars denote mean±SEM, p<0.01** (p=0.0036). (J). EVs were isolated from bronchoalveolar lavage fluid from people with CF with G551D mutation and NTA analysis of particles was performed (i) and particles were quantified (ii). Histograms represent the average of four biological triplicates. Statistical analysis used was paired Student t-test, and error bars denote mean±SD.
We then analysed EV release from in CuFi-4 cells (F508/G551D). Following treatment of CuFi-4 cells with ivacaftor (10 µM) for 24 hours, NTA analysis revealed a significant reduction in the number of EVs in CuFi-4 compared with NuLi-1 cells (p=0.0036; figure 2Ii), which was quantified in triplicate (figure 2Iii). As a control, we measured EV release from WT HBEo- and NuLi-1 cells after treatment with CFTR inhibitor-172. A significant decrease in EV release was observed from both treated cell lines compared with untreated (p<0.01; online supplementary figure S5). To further investigate whether these findings are clinically relevant, we measured EV concentration in the BALF of PWCF with a Gly551Asp mutation. BALF samples were obtained from children before (control) and after ivacaftor treatment. A decrease in EVs in the BALF of children treated with ivacaftor was observed compared with controls. However, it was not significant (figure 2Ji, Jii); this is most likely due to the small sample size available.
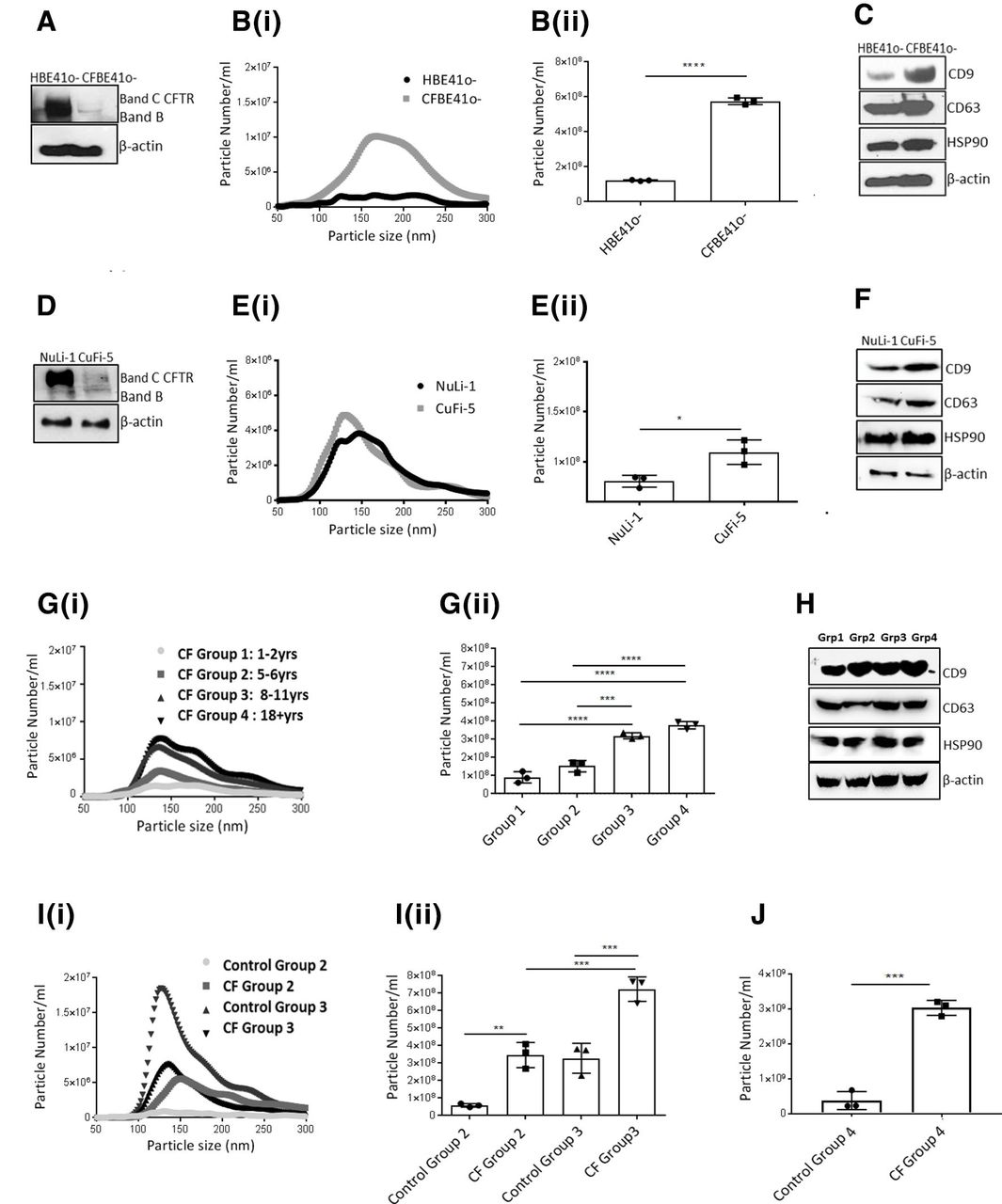
EV fractions from CFBE41o-and HBE41o- cells were subjected to protease digestion and the composition of the peptide mixture was determined using MS. A schematic is presented in figure 3A. EV proteins identified in CFBE41o- and HBE41o- cells by MS had at least one unique peptide per protein with a false discovery rate of ≤1% accepted using the Max Quant software. Proteins identified in at least two biological replicates of three were considered for further analysis. Using these criteria, 349 proteins were identified in CFBE41o- EVs and 157 in HBE41o- EVs. The complete list of EV proteins in these cell lines is available in online supplementary table 2. Ingenuity pathway analysis (IPA) revealed that CFBE41o- EVs were enriched immune pathways (acute phase response) and integrin signalling (online supplementary figure S6). A list of highlighted proteins present in the EVs and their corresponding pathways are presented in figure 3C. Validation of expression of EPCAM, VCAM and S100 A12, was confirmed by immunoblotting in both CFBE41o- and CuFi-5 EV fractions (figure 3D) to complement a number of exosomal markers already validated and shown in figure 1C and figure 1F (CD63, CD9, Hps90).
Supplemental material

Extracellular vesicles (EVs) isolated from cystic fibrosis (CF) bronchial cells and bronchoalveolar lavage fluid (BALF) fluid from people with CF (PWCF) are enriched in proteins involved in leucocyte recruitment/activation. (A) EVs were isolated from CF bronchial epithelial cell lines and BALF from persons with CF and subjected to mass spectrometry (MS) analysis. An experimental schematic is shown. (B) The number of proteins identified by MS in EVs isolated from CFBE41o-cells and HBE41o- cells in at least two replicates by the MaxQuant algorithm. Data displayed using Venny Software http://bioinfogp.cnb.csic.es/tools/venny/. (C) A selection of proteins (7) identified in EVs isolated from CFBE41o-cells and HBE41o- cells are displayed in the table and their corresponding pathway affiliation as determined by Reactome (https://reactome.org/). (D) Protein expression of CD9, CD63, Hsp90, EPCAM, VCAM and S100 A12 was determined in EVs isolated from CFBE41o-, HBE41o-, CuFi-5 and NuLi-1 cells to validate MS results (C). (E) EVs were isolated from 20 BAL samples obtained from PWCF (4 age groups) and controls were then subjected to MS analysis. The number of proteins identified by MS using the MaxQuant algorithm in BALF EVs isolated from the four groups of PWCF is displayed using Venny Software. (F) Subcellular mapping of all BALF EV proteins using FunRich (http://www.funrich.org/) is displayed. (G) Pathways affiliated with all the proteins identified in BALF from the four groups of PWCF as determined by Reactome (https://reactome.org/).
EVs were isolated from BALF of 20 PWCF of different age groups: paediatric group 1 (1–2 years), paediatric group 2 (5–6 years), paediatric groups 3 (8–11 years) and adult (18+yrs) in addition to 10 control samples. Clinical demographics of each group as before are outlined in online supplementary table 1. Proteomics analysis was performed on BALF EVs from PWCF and 1151 unique proteins were identified across all individuals. A Venn diagram mapping the number of proteins identified per group is displayed in figure 3E. A full list of proteins identified in BALF EVs of different age groups is available in online supplementary table 3. Subcellular fractionation using the FunRich database revealed that the proteins identified in BALF EVs are highly enriched in extracellular/exosomal proteins (figure 3F). Reactome pathway analysis of the BALF EV samples demonstrated significant enrichment in the expression of genes involved in inflammatory responses such as neutrophil degranulation and regulation of complement factors (figure 3G).
Supplemental material

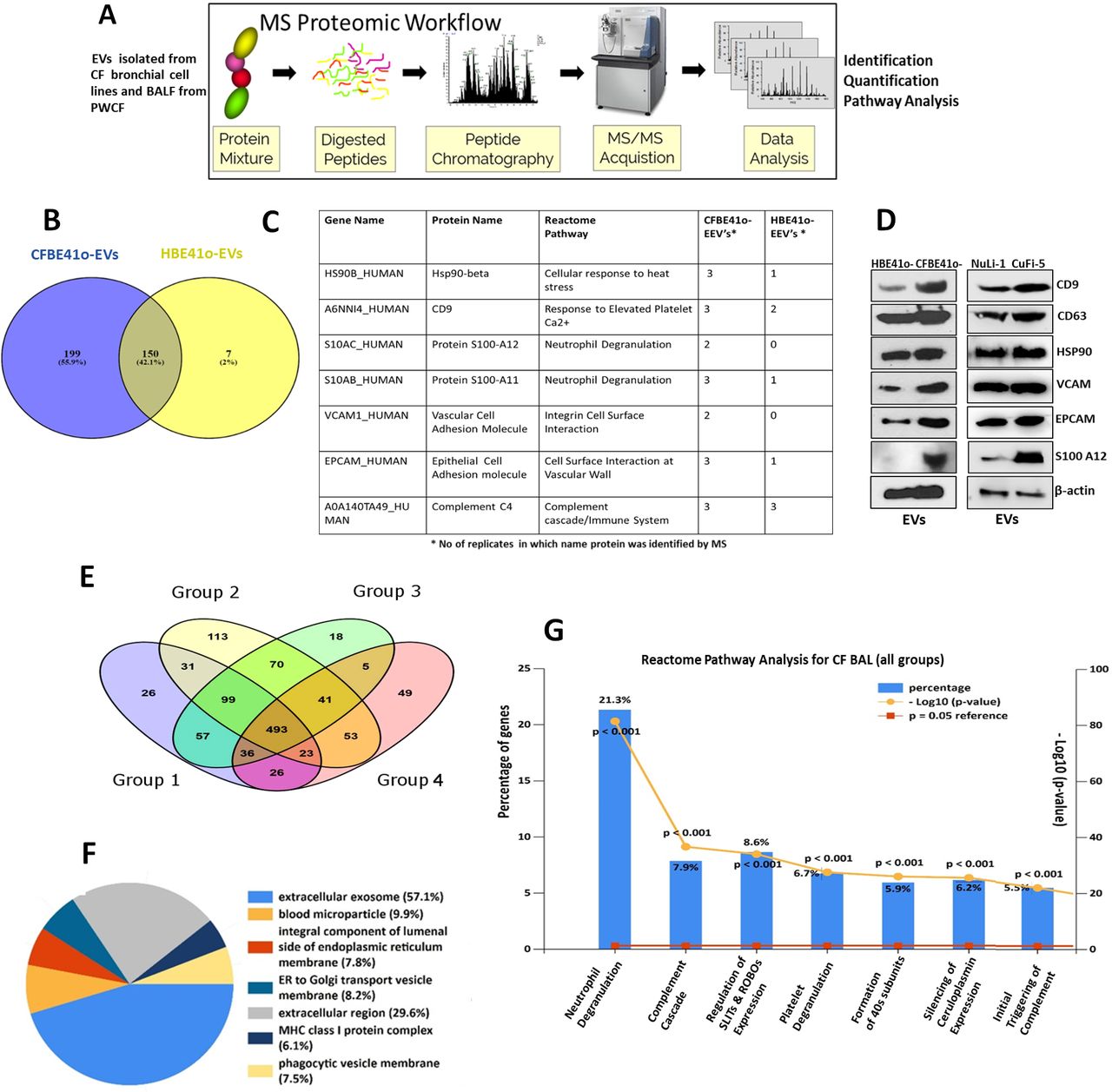
Bronchoalveolar lavage fluid (BALF) extracellular vesicles (EVs) have differential protein expression in people with cystic fibrosis (PWCF). Proteins identified in all EV fractions from the 20 BALF samples from PWCF (groups 1–4) in figure 3 were quantified by MaxLFQ (Accurate Proteome-wide Label-free Quantification) using MaxQuant V.1.5. Excel was used to further analyse the resulting normalised label-free quantification (LFQ) intensities for each protein in the four cystic fibrosis (CF) clinical groups. Cut-off values of 1.5 were used to determine protein expression fold change (ratios) between the four different CF age groups and statistical analysis was performed using unpaired t-test to identify significantly altered protein expression levels at different ages. Data are available in online supplementary table 4. (A) Eighteen proteins with significantly altered protein expression levels in BALF EVs isolated from PWCF in different age groups are highlighted. Protein expression upregulated and downregulated in groups 2–4 are compared with group 1 (1–2 years). (B). Six BALF EV proteins with significantly altered protein expression levels across the four age groups (1–4) are highlighted. The histograms represent LFQ intensities for five experiments and error bars represent the SD, p<0.05 (0.0040–0.0346). * P values were generated by analysis of variance test followed by Bonferroni adjustment. (C) The abundance of EV proteins isolated from CF paediatric patients (5–6 years) and controls were compared with clinically available interleukin (IL)-8 values. Correlation of proteins (protein abundance as determined by LFQ intensity) with recorded clinical IL-8 value for that patient (determined by ELISA) was calculated using Pearson correlation analysis. The correlation score-R and level of significance are listed for each protein.
Supplemental material
We quantified protein intensities across the different age groups using the MaxQuant algorithm which incorporates normalised label-free quantification (LFQ) (online supplementary table 4). This was to compare proteins upregulated and downregulated at different ages. Significance was then calculated between group 1 and the other age groups. In total, 45, 116 and 42 significantly changing proteins were identified between groups 1 and 2, groups 1 and 3, and groups 1 and 4, respectively (online supplementary table 4; p<0.001–0.05), confirming that there are changes in EV content with age in CF. Of note, the changes in adult protein expression may also relate to exacerbation status or chronic airway infection as well as age (online supplementary table 1). Encouragingly, significant changes were detected between groups of PWCF with stable disease (groups 1 and 2, groups 1 and 3). IPA was performed on proteins significantly different between the groups. Significantly enriched pathways changed with age; for example, complement activation was enriched in the older CF groups (online supplementary figure S7). A table of 18 proteins whose expression is significantly (p=0.01–0.05) altered between the different age groups is displayed in figure 4A which included S100 A12, CD53, that and cathelicidin antimicrobial peptide.17–19 A plot illustrating protein expression changes for six differentially regulated proteins between the younger and older age groups is displayed (figure 4B). Proteins identified in the paediatric control group and adult groups are listed in online supplementary table 5. Protein expression levels of 10 EV proteins in these control groups compared with their expression in CF EVs are displayed in online supplementary figure S8. In order to determine whether there was any correlation between EV protein expression changes and established clinical markers of disease in CF, we performed correlation analysis between interleukin (IL)-8 values measured in CF BALF and protein expression levels in BALF EVs. There was a significant correlation between IL-8 protein levels as determined by ELISA and the expression of 29 EV proteins as determined by MS (p<0.001–0.05; online supplementary table 6; figure 4C).
Supplemental material
Supplemental material

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Extracellular vesicles (EVs) from cystic fibrosis (CF) bronchial cell models induce chemotaxis of neutrophils which is mediated by EV-derived S100 A12. (A) Healthy donor neutrophils were incubated with PKH26-PE-labelled exosomes from CFBE41o- cells analysed by flow cytometry. Phycoerythrin (PE) fluorescence was measured in the neutrophil population post-EV incubation. (B) Neutrophils were stained with APC-conjugated CD66b and fluorescence was determined post-EV incubation. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.05*(p=0.049). (C) Healthy donor neutrophils (n=3) were exposed to CFBE41o- /HBE41o- EVs (10 µg) and 100 nM fLMP for 3 hours and myeloperoxidase (MPO) neutrophil activity was measured. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.05*(p=0.0440). P values were generated by analysis of variance (ANOVA) test followed by Bonferroni adjustment. (D) Neutrophils were isolated from healthy volunteers and neutrophil migration through a trans-well chamber was measured in response to treatment with CFBE41o-/HBE41o- EVs. The histograms represent an average of five experiments, and error bars denote mean±SD, p<0.05*(p=0.0352). P values were generated by ANOVA test followed by Bonferroni adjustment. (E) Healthy donor neutrophils were treated with CFBE41o-/HBE41o- EVs and immunoblotting performed for RAGE, p38, phospho p38, p44, phospho p44 and actin. (F) Neutrophils were isolated from healthy volunteers and neutrophil migration through a trans-well chamber was measured in response to treatment with CFBE41o- EVs treated with siRNA S100 A12. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.05*(p=0.0169). P values were generated by ANOVA test followed by Bonferroni adjustment. (G) Neutrophils were isolated from the blood of two paediatric groups of people with CF (PWCF). Neutrophil migration was measured in both paediatric group A (2–4 years) and group B (13–17 years) and compared with healthy controls (D) in response to CFBE41o-/HBE41o- EV treatment. Histograms represent the average of five (group A) and three (group B) experiments, and error bars denote mean±SD, p<0.01**(p=0.0024), (p=0.0023) and p<0.05* (p=0.0257). P values were generated by ANOVA test followed by Bonferroni adjustment. (H) Neutrophils were isolated from PWCF (group B) and neutrophil migration through a trans-well chamber was measured in response to treatment with CFBE41o- EVs treated with siRNA S100 A12. Histograms represent the average of three experiments, and error bars denote mean±SD, p<0.01**(p=0.0078). P values were generated by ANOVA test followed by Bonferroni adjustment.
To functionally assess the effects of EVs on neutrophil biology, CFBE41o- EVs were labelled with PKH26 fluorescent dye and the uptake was assessed by flow cytometry. Labelled EVs were taken up by healthy donor neutrophils (figure 5A). Neutrophils exposed to CFBE41o- EVs also displayed an increase in CD66b expression (figure 5B), indicating neutrophil activation. Neutrophils also increased in both the size and granularity in the presence of the EVs (online supplementary figure S9). To assess the ability of bronchial cell EVs (10 μg) to stimulate neutrophil degranulation, MPO activity was assessed. CFBE41o- EVs increased MPO neutrophil activity compared with RPMI and WT EVs (figure 5C; p=0.044). CFBE41o- EVs increased healthy neutrophil migration (figure 5D; p=0.0352) and transmigration (p=0.0002; online supplementary figure S10) through a coated CFBE41o- cell layer when used as a chemoattractant in cell migration assays. The RAGE receptor and downstream MAPK pathway activation were analysed. An increase in RAGE, phospho ERK and p38 activation was observed in neutrophils (isolated from healthy donors) following stimulation with CFBE41o- EVs compared with control (figure 5E). Due to the presence of S100 A12, a RAGE ligand in CFBE41o- and CF BALF EVs we examined if S100 A12 derived from bronchial EVs may regulate neutrophil chemotaxis. Gene silencing of S100 A12 in cell lysates and EVs was successfully performed and reduced expression of S100 A12 was observed (online supplementary figure S11a). A significant reduction in neutrophil chemotaxis was observed when S100 A12 expression in EVs was reduced via gene silencing (figure 5F; p=0.0169). Neutrophils were isolated from the blood of two groups of children with CF, paediatric group A (2–4 years) and group B (13–17 years). CFBE41o- EVs increased CF neutrophil migration in both groups compared with controls (online supplementary figure S12). There was a significant increase in chemotaxis of neutrophils from PWCF compared with neutrophils from healthy donors when stimulated with CFBE41o- EVs (figure 5G; p=0.0024). As RAGE activation was also induced in neutrophils from a person with CF after stimulation with CFBE41o- EVs (online supplementary figure S11b), we examined CF neutrophil chemotaxis using CFBE41o- EVs where S100 A12 expression had been reduced by gene silencing. Again a reduction in CF neutrophil chemotaxis was observed when S100 A12 expression in EVs was reduced (figure 5H).
Discussion
In the CF airway, the progressive inflammatory response begins early in life, impairing host defences, causing structural damage to the airway wall and impairing lung function.20 CF is caused by mutations in the CFTR gene; however, investigators have questioned whether the inflammatory response seen in CF is secondary or if the primary CFTR defect is a factor.21 We hypothesised that examining the role of EVs in CF could shed some light on this important question. In this study, we detected higher EV concentration in CF bronchial supernatants and in BALF from PWCF compared with their respective controls. Of interest, the number of EVs released increased with age in PWCF. This observation is supported by studies that have demonstrated exosome release increases with disease progression in a variety of disease states, including liver disease, pancreatic cancers and neurodegenerative disorders.22
We further explored whether EV release may inversely correlate with CFTR expression. Encouragingly, we observed a decrease in EV release when CFTR expression and stability increased with a series of AKT inhibitors and CFTR correctors, including lumacaftor, tezacaftor and ivacaftor used in clinical practice for PWCF.23 24 Decreased EV release also correlated with an increase in autophagy induction (LC3-II) in CF airway cells following treatment with AKT inhibitors and CFTR correctors. This observation could be explained by a coordinated relationship between the exosome release and autophagy pathways for the maintenance of cellular fitness.25 The inability of protein F508delCFTR to achieve a correct folded state disrupts cellular networks which protect the cell from acute stress26 and exosome secretion could serve an alternative way to alleviate stress when recycling pathways are compromised. We also detected a decrease in EV release from cell lines and PWCF harbouring the Gly551Asp mutation following ivacaftor treatment, which is known to increase the time that activated CFTR channels remain open. A relationship between CFTR channel activity and EVs was previously investigated in EV-transduced CF cells27; however, a more in-depth analysis into the relationship between EV release and CFTR activity would be required.
We performed MS analysis to define proteins and pathways enriched in EVs isolated from CF airway cell lines. Pathway analysis revealed a significant enrichment of proteins associated with acute inflammatory stress and infection in CFBE41o- EVs. The presence of inflammatory proteins in CFBE41o- EVs in the absence of any infection supports the possibility of an intrinsic inflammatory state constitutively existing in CF lungs as supported by other studies.28 29 CFBE41o- EVs were also significantly enriched in integrin proteins, for example, VCAM1 which mediates leucocyte adhesion to epithelial/endothelial surfaces.30 The presence of VCAM1 suggests that EVs could potentially cross the epithelial layer to surrounding blood vessels inducing migration of leukocytes into the alveolar space.31 Our data suggest that CFBE41o- EVs can regulate both migration and transmigration of blood-derived neutrophils through a CF epithelial monolayer.
We obtained BALF from 20 PWCF from four different age groups and performed MS analysis on the EV fractions to determine their protein content. There has been previous proteomic studies looking at protein expression changes in BALF and sputum from PWCF; however, they are not focused on EVs across a spectrum of age groups, highlighting the novelty of this study.32 Pathway analysis of these differentially expressed proteins in BALF EVs mapped to inflammatory pathways including cytokine signalling (IL-8 and IL-12) supporting previous findings on cytokine expression in CF airways.1 Interestingly, we observed an increase in S100 A12 expression levels in all CF groups. S100 A12 has been reported to stimulate migration of leukocytes to inflammatory sites.17 S100 proteins were previously identified in BALF taken from a CF paediatric group using MS33 and another study recently identified the presence of S100 A12 in a small number of CF airway EVs supporting our findings.8
In order to determine whether EVs isolated from CF bronchial cell lines could have functional relevance in modulating inflammatory response, we examined the ability of CFBE41o- EVs to bind and activate neutrophils from donors. Encouragingly, we observed that EVs were able to activate neutrophils, as well as stimulate neutrophil degranulation and induce chemotaxis. This supports a recent study by Majundar et al, that demonstrated that purified exosomes from cancer cell lines can activate resting neutrophils and elicit chemotactic activity in a LTB4 receptor-dependent manner.34 In order to identify some potential signalling drivers of neutrophil migration, we examined if CFBE41o- EVs could stimulate the RAGE receptor and its downstream pathways in neutrophils. RAGE has previously been shown to mediate neutrophil adhesion to, and subsequent migration across, intestinal epithelial monolayers.35 We then showed that when S100 A12, a ligand for RAGE, was gene silenced from EVs derived from CFBE41o- cells, a significant decrease in migration of neutrophils was observed. Although further mechanistic studies would be required to explore this pathway, these data suggest that CFBE41o- EVs have a potential functional role in neutrophil activation which may at least partially be mediated by S100 A12 RAGE signalling.
Overall, this study on CF EVs has provided exciting initial data on EV released from CF airway cell lines and in BALF from PWCF compared with their relative controls. We discovered that EVs from PWCF have differential protein expression at different age groups. Additionally, we discovered that current clinically used CFTR modulators can regulate EV release providing additional insights into their mechanisms of actions. Our data have also indicated that CF EVs can regulate chemotaxis of neutrophils, which can be regulated in part through S100 A12/RAGE signalling pathways. We acknowledge that subject numbers in this translational study are small; however, we feel that this study provides us with valuable and potentially very useful information in a novel and developing area in CF.
References
Footnotes
ZU and MPW are joint first authors.
Correction notice This article has been corrected since it was published Online First. Figure 3 has been corrected due to an incorrect actin included in figure 3D and the sentence relating to this has been amended from, ‘Validation of expression of EPCAM, VCAM and S100 A12, and exosomal markers (CD63, CD9, Hps90) was confirmed by immunoblotting in both CFBE41o- and CuFi-5 EV fractions (figure 3D).’ to ‘Validation of expression of EPCAM, VCAM and S100 A12, was confirmed by immunoblotting in both CFBE41o- and CuFi-5 EV fractions (figure 3D) to complement a number of exosomal markers already validated and shown in figure 1C and figure 1F (CD63, CD9, Hps90).’
Contributors Conception and design: JC, PM, ZU and MW. Experiments: ZU, MW, RR, ED, HC, KB and AT. Clinical data collection and support: RR, JL, HD, SCD, EM, SC, PM and BL. Analysis and interpretation: ZU, MW, HC, DM, KB, JC, SD, PM and AT. Manuscript preparation: JC, PM, ZU, MW and AT.
Funding This publication has emanated from research conducted with the financial support of The National Children’s Research Centre under Project Grant No C/17/3. HC and DF are funded by the European Union’s Horizon 2020 research and innovation under grant agreement no. 686098.
Patient consent for publication Not required.
Ethics approval The study was approved by the research ethics committee of Children’s Health Ireland at Crumlin, St. Vincent’s University Hospital and Tallaght University Hospital where informed consent was obtained from each participant.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article or uploaded as supplementary information.
Linked Articles
- Airwaves