Article Text

Abstract
Introduction Zinc is well known for its anti-inflammatory effects, including regulation of migration and activity of polymorphonuclear neutrophils (PMN). Zinc deficiency is associated with inflammatory diseases such as acute lung injury (ALI). As deregulated neutrophil recruitment and their hyper-activation are hallmarks of ALI, benefits of zinc supplementation on the development of lipopolysaccharides (LPS)-induced ALI were tested.
Methods 64 C57Bl/6 mice, split into eight groups, were injected with 30 µg zinc 24 hours before exposure to aerosolised LPS for 4 hours. Zinc homoeostasis was characterised measuring serum and lung zinc concentrations as well as metallothionein-1 expression. Recruitment of neutrophils to alveolar, interstitial and intravascular space was assessed using flow cytometry. To determine the extent of lung damage, permeability and histological changes and the influx of protein into the bronchoalveolar lavage fluid were measured. Inflammatory status and PMN activity were evaluated via tumour necrosis factor α levels and formation of neutrophil extracellular traps. The effects of zinc supplementation prior to LPS stimulation on activation of primary human granulocytes and integrity of human lung cell monolayers were assessed as well.
Results Injecting zinc 24 hours prior to LPS-induced ALI indeed significantly decreased the recruitment of neutrophils to the lungs and prevented their hyperactivity and thus lung damage was decreased. Results from in vitro investigations using human cells suggest the transferability of the finding to human disease, which remains to be tested in more detail.
Conclusion Zinc supplementation attenuated LPS-induced lung injury in a murine ALI model. Thus, the usage of zinc-based strategies should be considered to prevent detrimental consequences of respiratory infection and lung damage in risk groups.
- cytokine biology
- innate immunity
- neutrophil biology
- respiratory infection
- bacterial infection
Statistics from Altmetric.com
Key messages
What is the key question?
Does zinc have preventive potential in regard to acute lung injury?
What is the bottom line?
Zinc prevented lipopolysaccharides (LPS)-inhalation-induced lung damage, which was due to preservation of pulmonic barrier integrity as well as reduction of polymorphonuclearneutrophils recruitment and overreaction as observed in mice in vivo and verified using human cells in vitro.
Why read on?
The beneficial effects of zinc might not be limited to LPS or bacteria-induced lung injury, but zinc could be valuable to attenuate lung damage induced by mechanical stress, fumes and lung surgery.
Introduction
The essentiality of zinc as a gatekeeper of the immune system is well known1 and includes important roles in regulation of key polymorphonuclearneutrophils (PMN) functions.2–5 Zinc deficiency is strongly associated with a pro-inflammatory phenotype, an altered redox state, and was suggested to increase sensitivity to pulmonary injury elicited by hypoxia, alcohol abuse or sepsis.6–9 Also, altered membrane permeability and thereby decreased barrier function of especially lung, intestine and liver was described during zinc deficiency, explaining the more severe progression of inflammatory diseases observed in zinc-deficient patients.1 8 10 11 In this regard, benefits of zinc supplementation in normalisation of dysbalanced inflammatory immune reactions as well as in improving barrier function have been described.8 9 11–17
Increased recruitment and over-reactivity of neutrophils, peaking 4 hours after disease induction, are causative in several severe inflammatory diseases resulting in acute lung injury (ALI).14 18–24 Recruitment of PMN to the lung during ALI was found to be broadly governed by chemokines as well as by platelet-derived factors.23 ALI is also characterised by hyperinflammation and increased permeability of the alveolar-capillary barrier. This results in lung oedema, accumulation of protein-rich fluid and impaired oxygenation of arteries.25
Therapeutic approaches such as anti-inflammatory simvastatin and pioglitazone targeting neutrophils were quite successful in ameliorating the symptoms of ALI.21 22 Aspirin-mediated inhibition of platelet activation also decreased recruitment and activity of PMN and the inflammatory response.24 Unfortunately, pharmaceuticals have unwanted side effects such as damaging the gastrointestinal tract or the liver and can cause cancer.26–28 Therefore, the search for alternative treatments is ongoing, especially as ALI remains a significant source of morbidity and mortality especially in the critically ill.29 As zinc is known for its anti-inflammatory, anti-oxidative and immune-balancing activity as well as for its natural availability and its minimal-to-no side effects, its benefits to prevent or at least attenuate ALI were investigated in the current study.
Materials and methods
Animals
Sixty-four male C57Bl/6 mice, 8 weeks of age, (Janvier SAS; Le Genest-Saint-Isle, France) were kept under standardised conditions. The local ethical authorities approved all experiments and split into eight treatment groups with 4 to 10 mice each. The exact animal numbers used for each experiment are indicated within the figures. The mice were pretreated with 30 µg zinc per mouse (Unizink, Köhler Pharma, Germany) or vehicle (sodium chloride (NaCl)) via intraperitoneal injection, 24 hours before lipopolysaccharides (LPS) inhalation. Details can be found in online supplemental figure S1. For one set of experiments, zinc or NaCl were injected 1 hour after LPS-inhalation was finished.
Supplemental material
Cell isolation and culture
Heparinised peripheral venous blood from healthy volunteers was collected following informed consent and ethics committee approval. Primary neutrophils were isolated as described previously.5 NCI-H441 cells were a gift from Dr Singh, DWI – Leibniz Institute for Interactive Materials, RWTH Aachen University. U937 and NCI-H441 cells were cultured at 37°C in a humidified 5% carbon dioxide atmosphere in RPMI-1640 (Sigma-Aldrich, Germany) containing 10% FCS (PAA, Germany), 2 mM L-glutamine, 100 U/mL penicillin and 100 µg/mL streptomycin (all Sigma-Aldrich). Cells were preincubated with or without zinc sulphate (50 µM) alone or zinc sulphate (20 µM) in combination with pyrithione (10 µM) and stimulated with LPS (250 to 1000 ng/mL, Sigma-Aldrich) or granulocyte colony stimulating factor (G-CSF) (2000 U/ml, Chugai Pharma, Germany).
Murine model of LPS-induced acute lung injury
Acute lung injury was induced as described previously and in online supplementary figure 1 using aerosolised LPS.20 21 Thirty minutes before euthanasia, 5 µl fluorescein isothiocyanate (FITC)-Ly-6G (Gr1, eBioscience, Germany) and 100 µl FITC-dextran (30 mg/mL FITC-dextran; 70 kDa, Sigma-Aldrich) were applied by tail vein injection to label intravascular neutrophils.
Flow cytometry
To analyse the activation status of primary human neutrophils, cells were stained with anti-human CD35b and anti-human CD62L compared with the respective isotype controls (all BD Biosciences, Germany). Surface staining was assessed using a FACSCalibur and data analysis was performed using CellQuest software or using a FACSCanto II and FlowJo software (Tree Star, Ashland, Oregon). Labelling and gating strategy for cell pellets from bronchoalveolar lavage (BAL) of mice can be found in online supplementary figure S2.
Permeability
FITC-dextran (70 kDa (in vivo), 20 kDa (in vitro), Sigma-Aldrich) was used to assess vascular leakage or monolayer permeability. The fluorescence of the 100 µl BAL supernatant (FluoBAL) and of 50 µl serum (FluoSerum) was measured and permeability volume was expressed in microlitre (VPerm=(FluoBAL/100 µl)/(FluoSerum/50 µl)×BAL volume). For in vitro transwell experiments (3 µM pore size, Greiner Bio-One, Germany), dextran was added to the apical well and transport to the basal well was measured after 2 hours using a Spark 10 fluorescence well plate reader (Tecan Group, Switzerland). Permeability was calculated as fluorescence in basal well normalised to total fluorescence from apical and basal well.
Histology
After completion of the experiment, one part of the right lung was fixed in formalin, embedded in paraffin and stained with Mayer’s H&E for histological examination. Scoring of histological changes was done in compliance with the recommendation of the American Society.
Protein concentration of the BAL
The protein content of the BAL supernatants after centrifugation was assessed using the Bio-Rad protein assay based on the method of Bradford (Bio-Rad Laboratories Germany, absorbance at 700 nm).
ELISA
Serum and BAL samples were stored at −20°C until measurement, and only thawed once for cytokine detection. chemokine (C-X-C motif) ligand 1 (CXCL-1)/ Keratinocyte Chemotactic-1 (KC-1) and tumor necrosis factor (TNF) α protein production were quantified using OptEIA assays from BD Pharmingen according to the manufacturer’s instructions.
Neutral red test
Cellular viability was measured by neutral red uptake. NCI-H441 cells were loaded for 3 hours with neutral red (final concentration: 55 mM, Sigma), washed with phosphate buffered saline (PBS), lysed in a mixture of ethanol/water/acetic acid (50:49:1, Sigma) and neutral red uptake was quantified by its absorption at 540 nm using a Spark 10 (Tecan).
Neutrophil extracellular traps formation
Neutrophilextracellular trap (NET) structures were measured in mouse serum and supernatant of lung homogenate using myeloperoxidase (MPO)-DNA ELISA as described previously.24 A 96 well plate was coated with 50 µl/well MPO antibody (5 µg/mL, 4°C). After washing three times, 20 µl of samples were added to the wells with 80 µl immunoreagent (peroxidase-coupled DNA antibody and anti-Histone-Biotin antibody (cell death ELISA plus, Roche) and incubation buffer). After washing, peroxidase substrate was added, and absorbance was measured at 405 nm wavelength with a Spark 10. Values are presented as percentage increase of absorbance in comparison to the control group and as optical density. For primary neutrophils, the enhanced fluorescence emission of propidium iodide (PI, 10 mg/mL, Sigma) after its interaction with extracellular DNA was assessed. Cells were activated with phorbol myristate acetate (PMA) (50 ng/mL, Sigma) at 37°C for 4 hours, followed by PI staining for 5 min in the dark. The resulting fluorescence was recorded on a Spark 10 using an excitation wavelength of 360 nm and emission wavelength of 612 nm.
Flame atomic absorption spectrometry
Lung tissue was collected (approximately 50 mg) and digested in 2 mL nitric acid (American Chemical Society (ACS) grade reagent, ≥69%, Sigma) for 3 hours at 90°C. Lung tissue and serum zinc concentrations were measured by atomic absorption spectrometry (AAnalyst 800, PerkinElmer, Germany). Absorbance values for tissue zinc were calculated using a standard curve (0.1, 0.5 and 1.0 mg/L, zinc in nitric acid, TraceSELECT, Sigma) normalised to the wet tissue weight. This method allowed determination of µg zinc per gram wet tissue.
Reverse transcription and real-time PCR
Lung tissue was collected (approximately 50 mg) and total messenger RNA (mRNA) was isolated using TRIzol (Thermo Fisher) and complementary DNA (cDNA) was acquired through the use of a qScript cDNA Synthesis Kit (Quanta Biosciences, Germany) according to the manufacturer’s protocols. The primers for metallothionein-1, G-CSF, TNFα, the zinc transporters and the housekeeping gene glycerinaldehyde 3-phosphate dehydrogenase (GAPDH)15 30–32 were used as previously described. All samples were run in duplicate using a Step-1 (metallothionein(MT)-1, zinc transporters, GAPDH) or QuantStudio 3 (TNFα, G-CSF, GAPDH).15 Expression was calculated using a standard curve and values were normalised via the housekeeping gene.
Western blot
Western blot was performed as described previously.33 The phosphorylation state of signal transducer and activator of transcription 3 (STAT3) was analysed using primary antibodies against total STAT3, phosphorylated STAT3 (Tyr705) and phosphorylated STAT3 (Ser727) (all obtained from Cell Signaling Technologies, Germany). Resulting bands were quantified using ImageJ (ImageJ 1.46 r, Wayne Rasband, National Institutes of Health, USA).
Statistical analysis
Comparisons among groups and analysis of variance were implemented by GraphPad Prism software V.6 (GraphPad software, La Jolla, California, USA). Outliers were determined with the Grubbs’ outlier test. Mann-Whitney U test was performed for single comparisons and when only a few pairwise comparisons were of interest for the research question (figures 1 and 2, online supplementary figure S3-S5). P values <0.05 were regarded statistically significant for all tests and are illustrated in the figures by *, p values <0.01 are indicated by ** and p values ≤0.001 are indicated by *** . Kruskal-Wallis test with Dunn’s post-hoc test was performed for multiple comparisons (figures 3–6), p values <0.05 were regarded statistically significant, which is indicated by choosing different letters.
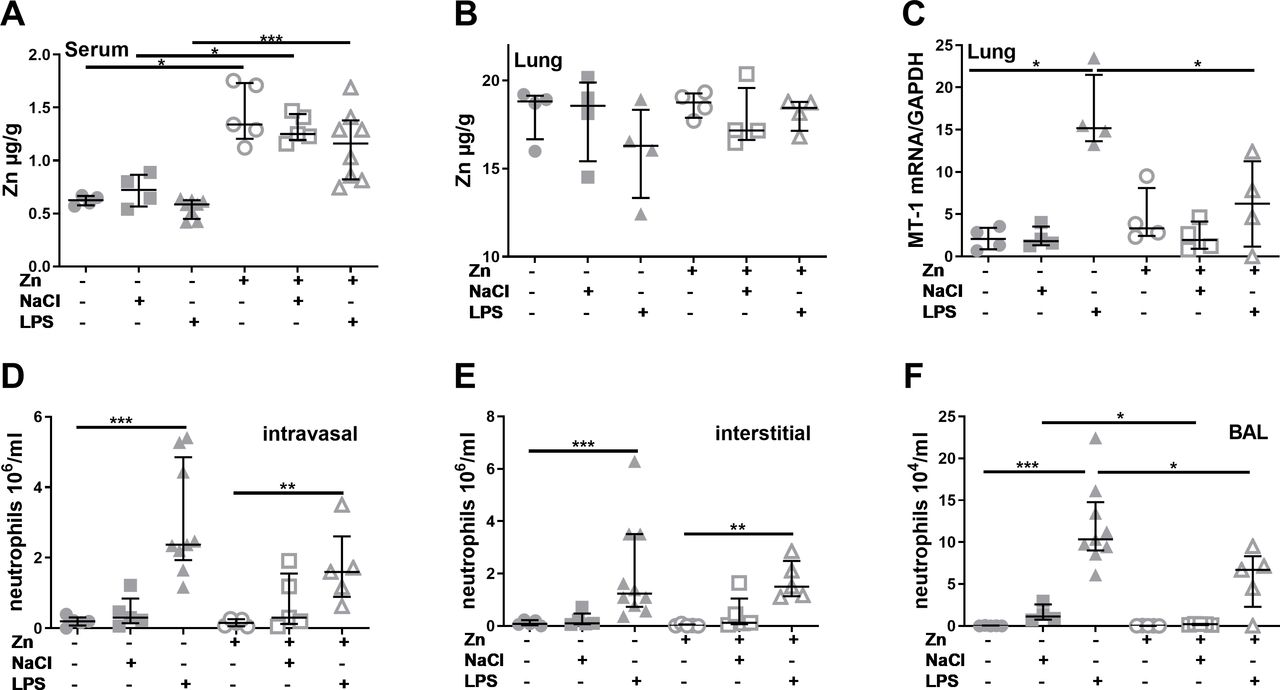
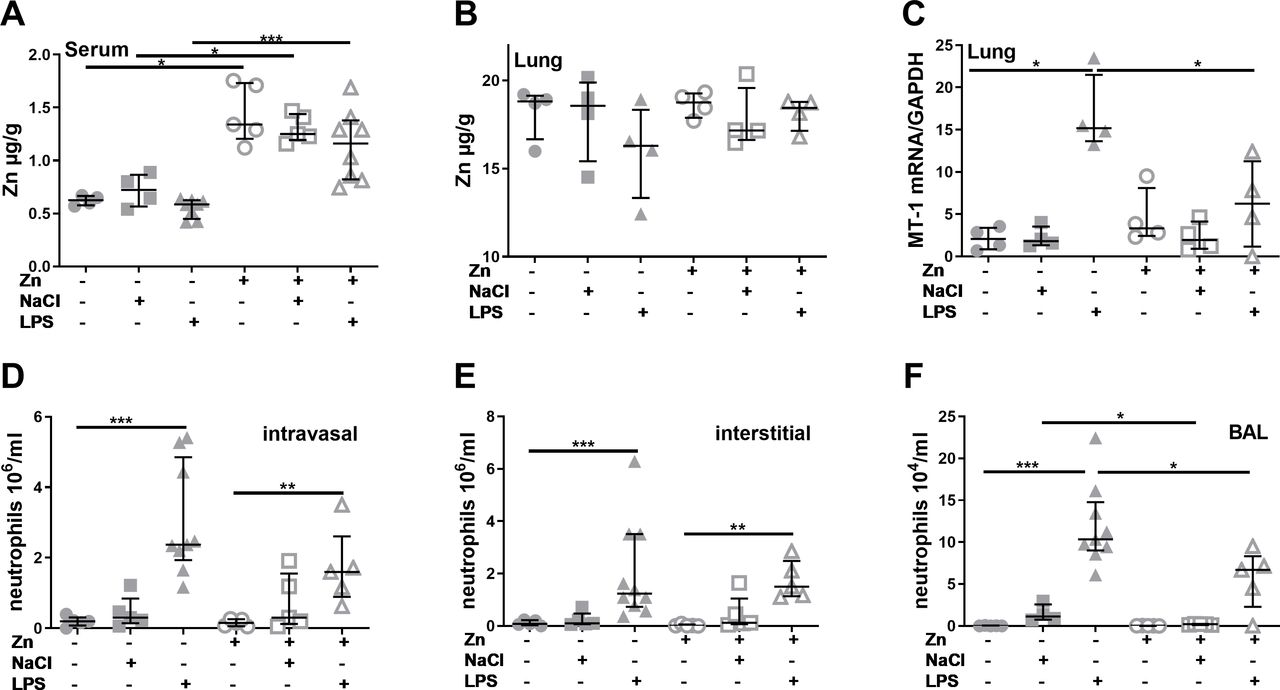
Zinc supplementation prior to acute lung injury induction alters zinc homoeostasis and polymorphonuclearneutrophils recruitment. Mice were treated as described in materials and methods. Serum and digested lung tissue were used to detect zinc content via atomic absorption spectrometry (A) + (B). Total mRNA from the lung was isolated, reverse transcribed into complementary DNA and quantified with SYBR green real-time PCR using a standard curve. The gene expression of murine MT-1 was normalised to the housekeeping gene GAPDH (C). Neutrophils were identified by their typical appearance in the forward scatter-side scatter and as CD45+, CD115– and PerCP- Gr1+ cells. Vithin the lung, FITC-Gr1 antibody was used to distinguish between interstitial neutrophils (CD45+, CD115–, PerCP- Gr1+, FITC-Gr1-) and intravascular neutrophils (CD45+, CD115–, PerCP- Gr1+, FITC-Gr1+). Intravasal neutrophils (D), interstitial neutrophils (E) and BAL neutrophils (F) are shown. Medians with quartile range for at least n=3 animals per group are shown. Statistical significance was tested by Mann-Whitney U test (*p<0.05; ***p<0.001). BAL, bronchoalveolar lavage; FITC, fluorescein isothiocyanate; GAPDH,glycerinaldehyde 3-phosphate dehydrogenase; mRNA, messenger RNA; MT, metallothionein; LPS,lipopolysaccharides;NaCl, sodium chloride; Zn, zinc.
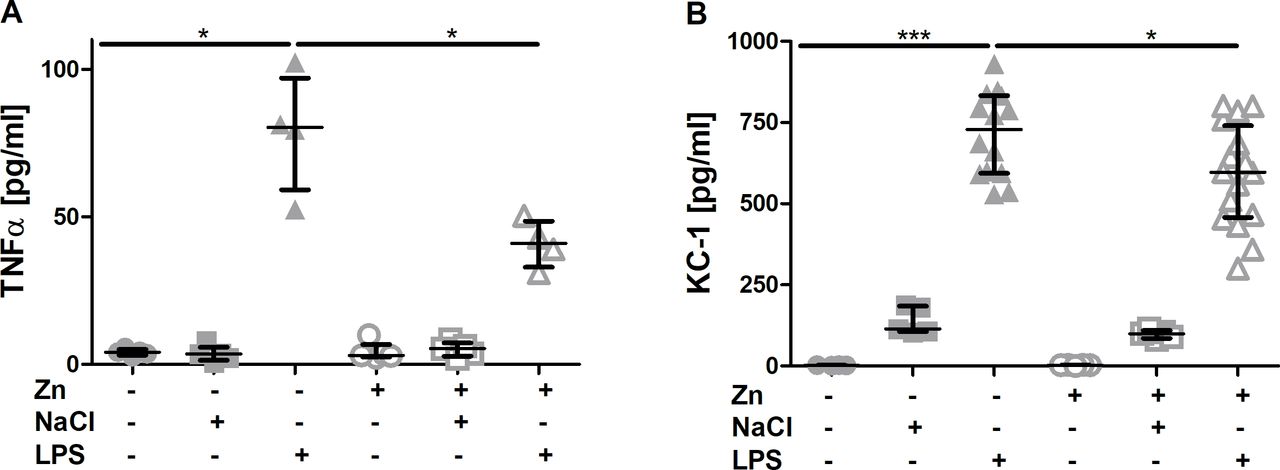
Effect of zinc supplementation on the inflammatory response. Mice were treated as described in materials and methods. TNFα (A) and KC-1 (B) content in the bronchial lavage fluid of the different experimental groups was assessed by ELISA analysing samples from at least n=4 animals per group. Medians with quartile range are shown. Statistical significance was tested by Mann-Whitney U test (*p<0.05). LPS, lipopolysaccharides; NaCl, sodiumchloride; Zn, zinc.

Consequences of zinc supplementation on acute lung injury-induced formation of neutrophil extracellular traps (NET). Mice were treated as described in materials and methods. NET formation was assessed in serum (A, C) and lung (B, D) either presented as per cent increase via measuring MPO concentration (presented as per cent increase of absorbance in comparison to the control group) (A, B) or measuring the optical density (C, D). Medians with quartile range for at least n=4 animals per group are shown. Statistical significance was tested by Kruskal-Wallis test. Significant difference at p <0.05, is indicated by different letters. MPO, myeloperoxidase; LPS, lipopolysaccharides;Zn, zinc.

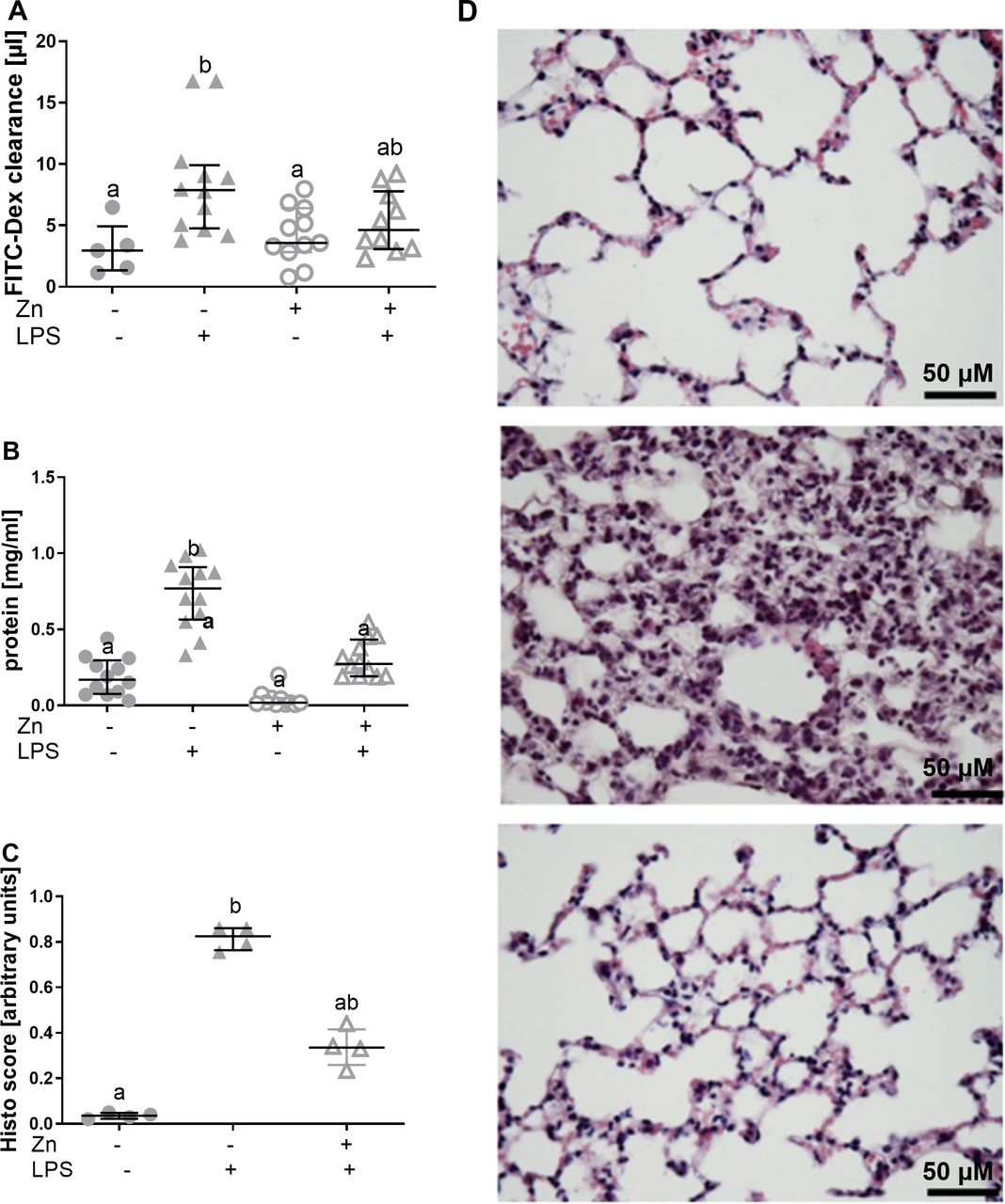
Zinc decreases LPS-induced lung damage. Lung permeability (A) and protein influx (B) are shown. Scoring of histological changes (C) was done in compliance with the recommendation of the American Society. Representative examples of Mayer’s H&E staining are shown (D) for control (top), LPS inhalation (middle), zinc supplementation prior to LPS inhalation (bottom). Medians with quartile range for at least n=4 animals per group are shown. Statistical significance was tested by Kruskal-Wallis test. Significant difference at p <0.05, is indicated by different letters. FITC-dex, fluorescein isothiocyanate-dextran; LPS, lipopolysaccharides; Zn, zinc.
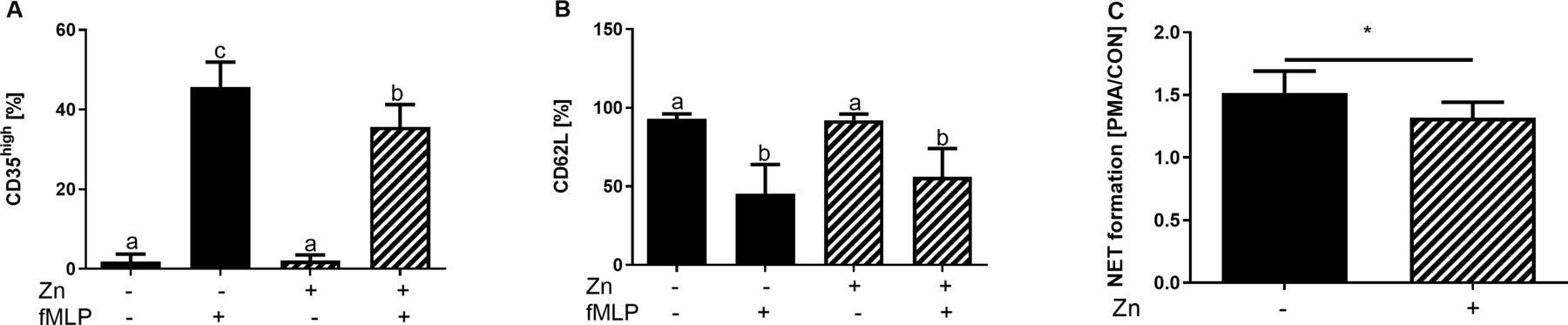
Consequences of zinc (Zn) supplementation on surface receptor expression and neutrophilextracellular trap (NET) formation by primary human neutrophils. Neutrophils were isolated from peripheral blood of healthy human volunteers, preincubated with or without zinc (50 µM) for 24 hours and stimulated with FMLP (A, B) for 4 hours. Flow cytometric data for CD35 (A) and CD62L (B) surface expression are shown. NET synthesis was assessed via propidium iodide fluorescence 4 hours after PMA stimulation. Fluorescence was normalised over unstimulated samples and is shown as means+SEM for at least n=7 neutrophil donors per group (C). Statistical significance was tested by Kruskal-Wallis or Mann-Whitney U test. Significant difference at p <0.05, is indicated by different letters or as *p <0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of zinc supplementation on LPS-induced damage to NCI-H441 cell monolayers. NCI cells were seeded into a 96 well plate (A), a 12 well Transwell plate (B) or a 12 well plate (C, D), preincubated with or without zinc (50 µM) for 24 hours and subsequently stimulated with or without LPS (250 ng/ml or 1000 ng/ml) for 4 hours. Neutralred test assay was performed and values were normalised to unstimulated control (A). For permeability analysis, apical wells of the transwells were loaded with FITC-dextran 2 hours after LPS stimulation (1000 ng/mL) was started and transfer of the fluorescent dye to the basal part was measured after another 2 hours and normalised to total FITC-dextran fluorescent (B). Total mRNA was isolated from the cells (C, D) and used to analyse expression of TNFα and G-CSF, quantified via a standard curve. Values were normalised to GAPDH mRNA levels. (E, F) U937 cells were FCS-starved (1% FCS), preincubated with ZnSO4 (20 µM) and pyrithione (10 µM) and stimulated with G-CSF [2.000 U/mL] for 60 min. Phosphorylation of STAT3 at tyrosine (Y)705 (E) or serine (S)727 (F) was analysed via western blot and normalised to total expression of STAT3 using a panSTAT3 antibody. Means+SEM for at least n=3 independent experiments are shown. Statistical significance was tested by Kruskal-Wallis test. Significant difference at p <0.05, is indicated by different letters. FITC,fluorescein isothiocyanate; GAPDH, glycerinaldehyde 3-phosphate dehydrogenase; mRNA, messenger RNA; LPS,lipopolysaccharides;Zn, zinc.
Results
Severe and systemic inflammatory diseases such as sepsis or pneumonia are well-known causes for the development of ALI.19 34 To investigate possible benefits of zinc supplementation for ALI prevention or amelioration, a total of 30 µg zinc per mouse was injected intraperitoneally 24 hours before ALI induction. This concentration is in a physiological range and has shown beneficial effects in earlier studies on multiple sclerosis.35 LPS inhalation was used to elicit the disease, which is a well-accepted model for ALI as it induces neutrophil recruitment, lung damage, pulmonary oedema and impairs gas exchange.36 Samples were taken 4 hours after disease induction, as this is the time when PMN levels in the lung peak and the cells are highly reactive.37
Zinc injection 24 hours prior to disease induction affects inflammation-induced changes in zinc homoeostasis and neutrophil recruitment
As a first step, effects of zinc supplementation and LPS inhalation on zinc homoeostasis were characterised by assessing zinc contents in serum and lung tissue as well as the expression of zinc transporters and MT-1, the primary intracellular zinc binding protein. While the zinc content of the lung tissue was not affected by zinc or LPS treatment (figure 1B), serum zinc levels were significantly higher in zinc supplemented animals (figure 1A). LPS treatment resulted in a slight but not significant decrease compared with the respective unstimulated controls. Expression of MT-1 in the lung was significantly increased after LPS stimulation (figure 1C). Interestingly, zinc supplementation blocked LPS-induced MT-1 expression.
Transport of zinc is regulated by 24 zinc transport proteins, either increasing cytoplasmic zinc levels (Zip1 – 14) or decreasing it (ZnT1 – 10). Zinc supplementation induced the expression of Zip8 and ZnT1 (table 1). All other transporters that were tested showed only trends as is summed up in table 1.
Expression of zinc transporters in the lung
A multitude of studies found PMN to be largely responsible for the tissue damage caused by LPS inhalation.19–21 23 In this study, inhalation of NaCl did not significantly induce PMN recruitment to the lungs, while LPS inhalation resulted in intravascular and alveolar lung infiltration by PMN within 4 hours (figure 1D,E). When zinc was injected 24 hours prior to disease induction, an increased recruitment of PMN compared with controls was still visible. However, especially in the BAL the amount of recruited PMN was significantly lower for zinc supplemented than non-supplemented mice.
Effects of zinc on PMN activity in serum and inside the lungs
To more closely investigate the activity of PMN, we chose an LPS-induced PMN-specific function, that is, the formation of NETs.4 Here, DNA with intercalated antimicrobial peptides such as MPO is secreted by PMN, forming a net to catch and destroy invading pathogens. MPO is also a very PMN-specific peptide, making this enzyme ideal for investigating PMN specific NET formation. DNA-bound MPO secretion was measured via ELISA and the optical density of DNA was detected.
As can be seen in figure 3, LPS inhalation induced the formation of NETs as observed in serum as well as in the lung homogenate. Moreover, both data sets indicate a decreased LPS-induced NET formation when zinc was supplemented 24 hours prior to ALI induction, underlining that zinc decreases PMN activity to control levels. In general, the extent of MPO concentration was much higher in lung homogenate than in the serum, suggesting a primarily local activation of PMN (figure 3).
Consequences of zinc supplementation on the LPS-induced TNFα and KC-1 (CXCL1) secretion and lung damage
Highly increased levels of pro-inflammatory cytokines as well as increased levels of chemokines such as KC-1 are hallmarks of inflammatory diseases including acute lung injury.19 To determine the local and systemic extent of the inflammatory response 4 hours after disease induction, TNFα levels were measured in BAL and serum of mice. LPS but not NaCl inhalation significantly increased TNFα levels in the BAL of mice after 4 hours. When zinc was injected 24 hours before LPS inhalation, levels were reduced to concentrations comparable to those found in controls (figure 2A). A significant reduction by zinc was found for KC-1 levels in the BAL (figure 2B). Changes of KC-1 and TNFα concentrations in serum were neither detected in animals after LPS inhalation nor in response to zinc injection (data not shown). This suggested that the inflammation was rather local at this early time point at least regarding mediators not specific to only PMN.
As a final step the hypothesis that zinc supplementation rescues the lung from severe damage was tested. Similar to what had been reported before, a significantly increased permeability as well as significantly higher protein influx into the alveoli was measured 4 hours after LPS inhalation. Zinc injection prior to inhalation was able to reduce the LPS-induced increased permeability to control level (figure 4A). The same was observed for protein influx suggesting protective effects of zinc against LPS-induced lung damage (figure 4B). This suggestion is underlined by the finding, that LPS alone induced significant lung damage as monitored by histological changes, while preventive zinc supplementation protected the lung (figure 4C,D).
Zinc decreases PMN activity and barrier damage in a human in vitro model
To verify key findings using human cells, primary neutrophils were isolated from healthy donors, preincubated with zinc for 24 hours and stimulated with LPS for 4 hours, just as performed in mice. Zinc preincubation ameliorated neutrophil activation by LPS, as was monitored by analysing CD35 and CD62L expression. LPS-induced expression of CD35 was significantly ameliorated by zinc and for CD62L loss, a decrease due to zinc administration was observed (figure 5A,B). Furthermore, NET formation was significantly lower in zinc supplemented fMLP-stimulated neutrophils compared with zinc adequate cells (figure 5C).
Also, zinc preincubation protected NCI-H441 pulmonary cells from LPS-induced cell death as was assessed using a vitality test (figure 6A). While LPS was able to increase permeability of a NCI-H441 monolayer, this effect was ameliorated, when the cells were supplemented with zinc 24 hours before high or low dose LPS-stimulation (figure 6B). Moreover, zinc supplementation reduced the LPS-induced expression of G-CSF and TNFα mRNA to control levels (figure 6C,D). G-CSF-induced STAT3-mediated signalling is well known to emergency granulopoiesis and PMN recruitment to tissues during inflammation.38 39 Thus, effects of zinc on G-CSF-induced signalling were investigated, using myeloid U937 cells as a model. figure 6 reveals that zinc preincubation significantly inhibits G-CSF-induced STAT3 phosphorylation at tyrosine 705 (activating phosphorylation), while phosphorylation at serine 727 (inhibiting) is increased by zinc (figure 6E,F).
Zinc supplementation 1 hour after LPS-inhalation does not affect relevant disease characteristics
During a final set of experiments, zinc was injected 1 hour after LPS-inhalation was finished, using the same concentrations as above. The numbers of PMN recruited to the lungs remained unaltered after therapeutic zinc injection (S3). Numbers appeared to be even higher in zinc treated animals, which was however not significant. No significant changes were found for the levels of TNFα and KC-1 in the BAL of mice that were injected with zinc after disease induction (S4). Results on lung damage were in line with this, as the histological score was slightly elevated as well as the protein influx and the clearance values (S5). Other times or dosages might need to be tested, but a positive effect of zinc during the early stages of disease seems to be unlikely.
Discussion
Altogether, the data from this study indicated that preventive zinc supplementation is able to reduce excessive lung damage by decreasing the LPS-induced recruitment as well as activation of PMN at least in regard to the first 4 hours after disease was induced. Direct protective effects of zinc on lung cells and on barrier integrity are possibly involved as well.
The one-time injection of 30 µg zinc per mouse caused a stable increase in serum zinc levels within 24 hours. Serum zinc levels are known to change quickly when zinc is supplemented, which is in line with our data.8 14
During regular acute phase response, zinc is transferred from serum to the liver, causing transient serum hypozincemia.17 31 In our study, however, which was the first investigating zinc homoeostasis shortly after LPS inhalation, we did not observe significant changes in serum or lung zinc concentrations within the first 4 hours after LPS inhalation. The fact that comparable studies analysed serum zinc after 24 hours explains the discrepancies. Also, LPS was inhaled in our study instead of injected and thereby caused a strong but rather local inflammatory response at the beginning, which probably spreads to the periphery at later time points. Results for MT-1 and zinc transporter expression also indicate that intracellular zinc of pulmonary cells is increased, supporting survival and regeneration of the cells, as has been shown for lung and liver cells before.7 16 31 40
In this project, we used TNFα expression to monitor the extent of local and systemic inflammatory reactions. As expected, high levels of TNFα were found in BAL of mice 4 hours after LPS inhalation. The decreased cytokine levels due to zinc supplementation measured here are in line with other studies describing anti-inflammatory effects of zinc and its ability to prevent overshooting inflammatory responses.8 14 17 Zinc is not completely abolishing the inflammatory response, which probably allows for a low grade of inflammation as response to pathogens and subsequent tissue healing. However, no TNFα was detected in serum of the mice, probably due to the early time point of investigation and the rather local activation of cells within the lung.
Recently it was shown that blood leucocyte counts negatively correlate with plasma zinc levels, resulting in highly elevated cell numbers in the blood when serum zinc levels are low.17 Our finding that zinc supplementation significantly decreased recruitment of PMN to the alveoli is novel, but in line with a study investigating allergic reactions, where decreased numbers of macrophages, basophils and lymphocytes were found in the lungs of zinc supplemented animals.7 In contrast to our study, higher zinc concentrations (minimum of 100 µg/mouse) were supplemented for longer periods of time (weeks).7 Our results revealed that a one-time treatment with 30 µg zinc per mouse 24 hours before LPS-inhalation was enough to achieve significant benefits regarding ALI amelioration when the first 4 hours were assessed, which is promising regarding the use of zinc in disease prevention.
Despite of zinc supplementation, LPS-inhalation still resulted in PMN recruitment to the lungs compared with NaCl-inhaling control animals. However, a complete blockade of PMN recruitment would not be conducive, thus zinc supplementation seems to offer an ideal tool to prevent hyperactivation of the innate immune system while preserving a basic immune response and sustain bacterial clearance.17
The decreased recruitment of the PMN to the lungs in zinc-supplemented animals, is probably the results of a combination of sustained lung integrity, decreased apoptosis of pulmonary cells, milder activation of PMN and lower chemokine expression:19
(I) Zinc is known to sustain membrane integrity, in case of the lungs in large parts due to its cytoprotective and anti-apoptotic activity in pulmonary endothelium, but also due to regulating the expression of tight junction molecules and protection from mechanical stretching stress.6 8 9 40 Our data show less lung permeability and damage in zinc supplemented animals and human pulmonary cell monolayers as well, hindering PMN from entering the lung alveoli. Increased survival and cytokine secretion of LPS-stimulated NCI-H441 cells that were pretreated with zinc, as well as reduced tissue damage, as shown histologically, underline this suggestion.
(II) In a previous in vitro study, zinc deficiency decreased chemotaxis, which was restored, when zinc was replenished,3 5 while our study shows that neutrophil recruitment was decreased by zinc supplementation in vivo. However, here the inflammatory response was induced by LPS, in contrast to chemotactic agents without further stimuli that were tested by others in vitro.5 The significance of our data is supported by results from psoriasis patients, suffering from zinc deficiency, where neutrophil migration was disturbed during disease, but normalised by zinc supplementation.2 The discovery that chemokine expression is decreased by zinc supplementation prior to LPS-inhalation offers one more explanation for our findings. Interestingly, our group recently observed significant effect of zinc in GM-CSF-induced signalling.33 Similar effects were seen when G-CSF-induced signalling was investigated in U937 cells. In summary, G-CSF levels as well as the signalling pathways it activates might play a role in zinc’s benefits during ALI, which is currently investigated further.
(III) Finally, zinc alters some platelet functions41 42 and might affect their chemokine production as was shown for treatment with aspirin.24 Although not investigated for platelets, zinc affects chemokine expression in other cells.5 Thus, this should be considered as an explanation here as well.
In addition to reduced recruitment, the results from this study indicate altered activation of PMN as well. This is underlined by the observation that NET formation within the lung fluid was much higher than what was detected in the serum. The hypothesis that zinc supplementation decreases the hyperactivation of neutrophils is further strengthened by our in vitro findings showing less LPS-induced activation of primary human neutrophils from peripheral blood when those were pretreated with zinc.
The decreased NET formation by PMN from zinc supplemented animals observed here deviates from former in vitro data, where zinc deficiency was shown to decrease NET formation by PMN.4 However, in the in vitro study by Hasan et al only zinc deficiency and replenishment, but no supplementation was tested.4 The in vitro data from primary human granulocytes from this study are in line with the in vivo data, showing decreased NET synthesis by zinc supplemented cells that were challenged.
One could question our hypothesis that zinc decreases reactivity of neutrophils because higher levels of NETs might simply be due to the increased numbers of cells. We cannot completely deny this objection. However, NaCl inhalation caused an increase in PMN recruitment to the lungs but did not affect NET formation. Furthermore, the number of PMN inside the lungs of zinc supplemented animals is still significantly elevated, while the levels of NET formation were not significantly higher than in the control animals; a hint, that ALI amelioration by zinc is due to both, lower PMN numbers and a reduction in their reactivity. Finally, NET synthesis in primary human neutrophils was investigated on per cell basis and results revealed ameliorated NETosis by zinc supplemented cells. As our study concentrated on investigating the early inflammatory phase during the first 4 hours after LPS-inhalation, long-term effects can only be predicted from what has been published in the literature. Although this is a weakness of the presented study, benefits of zinc supplementation can be expected in humans as well, taking into account results from later time points that were investigated in other studies, where benefits of zinc supplementation before disease onset were also shown.8 14 17
More particularly, the supplementation of zinc should be considered before any planned major surgery and other conditions that have a high risk for the development of ALI. In contrast to iron, there is no major and long-term store for zinc in the human body. However, in vitro data support the hypothesis that blood and tissue cells are able to take up zinc from the extracellular space during high zinc conditions. This would prepare especially the immune cells to react, but not overreact, to the stressful situations such as mechanical-induced, ventilation-induced or infection-induced lung damage that may occur during and shortly after surgery. Furthermore, preventive zinc supplementation may attenuate the overly intense drop in serum zinc levels that was associated to sepsis and morbidity therefrom and observed as response to severe infections and tissue damage in the clinic.17 43 44 Treatment of established disease with zinc is, however, not recommended by currently available data, but might be applicable, if treatment strategies are optimised.
To conclude, our results suggest that zinc supplementation is sufficient to reduce the number of highly reactive PMN entering the tissue while sustaining a moderate immune response to attack invading pathogens at least during the first 4 hours after ALI induction. Zinc’s support of lung integrity not only attenuates excessive PMN recruitment to the lungs, but probably also represents an obstacle preventing pulmonary pathogen entering the blood stream. Results of our study add zinc to the list of options to prevent the development of ALI which is a cheap, natural and low-in-side-effects alternative to currently applied therapeutics. The results underline the significance of a balanced zinc homoeostasis for an appropriate reaction to inflammatory stimuli and suggest to especially consider zinc supplementation of subjects at risk of zinc deficiency and of patients who are endangered of lung injury such as people undergoing surgery. Treatment of active inflammatory diseases using zinc remains at high risk and more studies are required to find suitable time points and zinc doses for application during established diseases.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors Gabriela Engelhardt and Zohreh Packsereshtmogharab contributed excellent technical assistance. Oliver Pabst, flow cytometry facility of the UK Aachen supported FACS measurements. Dr Singh, DWI – Leibniz Institute for Interactive Materials, RWTH Aachen University, kindly providing NCI-H441 cells.
Funding This study was funded by START-Program of the Faculty of Medicine, RWTH Aachen (grant number:116/16).
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval Ethics approval obtained from the local ethical authorities for all experiments (Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen AZ 84–02.04.2013–3062) and ethics committee (RWTH Aachen University Hospital, statement no. EK 023/05).
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available upon reasonable request. All data relevant to the study are included in the article or uploaded as supplementary information.
Linked Articles
- Airwaves