Article Text

Abstract
Asthma, an inflammatory disease of the airways, is a common condition, currently affecting about 9% of adults. Obesity is another disease with high prevalence in adults, and asthma that develops in obese individuals appears to be distinct from other forms of asthma, in being particularly severe and difficult to control even with oral corticosteroids. Moreover, recent studies suggest that obesity may have a causal relationship with asthma. In this review, we discuss possible obesity-driven metabolic and immunological pathways that might lead to asthma that is associated with obesity. Our understanding of the mechanisms that underlie this association will likely lead in the future to improved treatments for this significant unmet medical need.
- Asthma
- Cytokine Biology
- Asthma Mechanisms
- Innate Immunity
- Paediatric asthma
- Systemic disease and lungs
Statistics from Altmetric.com
Introduction
While obesity and asthma are common conditions, a specific association of obesity with asthma was only described 17 years ago in 1999 in a study of asthma among 61 000 women in the Nurses Health Study II.1 In that study, the investigators found that the relative risk of developing adult-onset asthma increased with body mass index (BMI), such that if the BMI was 30 or greater (meeting the definition of obesity), the risk of developing asthma increased by more than 2.5-fold.1 Initially, this idea was met with some scepticism, but additional studies confirmed the relationship: the odds of developing asthma increased with increasing BMI;2 higher BMI was associated with greater airway hyper-reactivity (AHR),3 a cardinal feature of asthma and weight reduction in obese patients improved asthma control, quality of life and AHR.4
Since the first study of asthma associated with obesity in 1999, the prevalence of obesity has continued to increase in industrialised societies, now affecting 38% of all adults in the USA (35% of men and 40% of women)5 while, concomitantly, the prevalence of asthma associated with obesity has also increased. Thus, the Centers for Disease Control recently reported that in 2011–2014 the prevalence of asthma among adults with obesity was 11.1%, significantly higher than that in adults with normal weight (7.1%).6 The prevalence of asthma is particularly high in obese women (14.6% compared with that in women with normal weight, 7.9%) for reasons that are not fully understood, but nevertheless, consistent with the idea of an important causative role of obesity for asthma.
Characteristics of obesity-associated asthma
Asthma is considered to be a heterogeneous and complex disease associated with several distinct phenotypes, some of which are still being defined. Traditionally, asthma has been divided into two major phenotypes. The first type, an allergic or type 2 phenotype, is triggered by exposure to allergens and associated with airway eosinophilia and adaptive immunity, involving T cells and B cells and immunological memory. The second type, a broad non-allergic or non-type 2 phenotype, is associated with exposure to air pollution and infection. These non-allergic asthma phenotypes are often associated with neutrophils in the airway and primarily with innate immunity, although adaptive immunity might also be involved. Since obesity is not generally associated with or dependent on allergic disease and immunological memory, the asthma associated with obesity has been thought to fit in with the non-type 2 phenotype. However, the precise mechanisms by which nutritional excess and adiposity lead to asthma have been poorly understood, and the role that allergy might play when present in some obese patients with asthma has not been clearly defined.
The specific clinical characteristics of obesity-associated asthma include poorly controlled asthma, severe disease associated with oral/systemic corticosteroid use, refractoriness to corticosteroid therapy, adult-onset type asthma in women and reduced serum IgE levels with few allergies. The severity of obesity-associated asthma likely explains the increased healthcare utilisation and reduced quality of life experienced by such patients.5 In addition, initial studies indicated that patients with obesity-associated asthma had low sputum eosinophils and increased airway neutrophils, although there was overall low airway inflammation and possibly a redistribution of eosinophils in the submucosa of the airways.7
More recent studies, however, have described a distinct early onset form of obesity-associated asthma, occurring in children, aged below 12 years,8 and associated with higher serum IgE levels, the presence of environmental allergies and a higher likelihood of prior intensive care unit admissions (more severe disease). In such obese patients with allergy and asthma, eosinophils may play an important role. Based on these observations, obesity-associated asthma may consist of two forms of severe asthma, one form (late onset) with a non-type 2 phenotype, and the other form (the early onset version) comprising a variation of type 2 asthma perhaps representing pre-existing allergic asthma, complicated by the development of obesity.9
Mechanisms of obesity-associated asthma
Obesity is known to directly alter lung mechanics by affecting airway wall thickness, respiratory rate, inspiratory effort and promoting sleep apnoea, which is frequently observed in patients with obesity-associated asthma. These characteristics can confound the clinical evaluation of asthma and lead occasionally to a misdiagnosis of asthma in obese individuals,10 as well as confound the understanding of how obesity might directly cause asthma. However, the observation that weight reduction with bariatric surgery improves asthma severity, control, AHR, lung function and quality of life, at least in the adult-onset form of obesity-associated asthma, suggests a direct effect of obesity in causing asthma.4 ,11 Note that, while weight reduction is beneficial in adult-onset obesity-associated asthma, weight reduction in obese patients with early onset disease with elevated serum IgE levels has limited effects on asthma.4 Together, these data suggest that the adult-onset form of this disease may be more directly caused by obesity than the early onset form, and that the early onset form may require aggressive treatment of both the obesity and type 2 allergic inflammation.12
Other mechanisms of obesity-associated asthma
While the early form of obesity-associated asthma may involve a complex interaction between type 2 inflammation and obesity, both the adult and early forms may involve metabolic pathways that can occur independently of type 2 inflammation. In this regard, adipokines produced by adipose tissue can affect the inflammatory process in the lungs, and might be involved in asthma pathogenesis in obesity. For example, adiponectin, an anti-inflammatory mediator that normally reduces airway inflammation, is decreased in obesity, whereas leptin, which can increase AHR, is increased in obesity.13–15 In addition, tumour necrosis factor (TNF)-α16 and oxidative stress17 ,18 are thought to contribute to the development of obesity-associated asthma. However, the precise pathways by which adipokines, TNF-α and oxidative stress result in asthma remain elusive.
In terms of metabolic and immunological parameters, obesity is widely recognised as a cause of chronic inflammation that contributes to the development of chronic diseases including type 2 diabetes, atherosclerosis and liver disease and some forms of cancer.19 Metabolically, caloric excess and the resultant metabolic-associated inflammation profoundly affect the immune system. For example, metabolic dysfunction in obesity is associated with an increase in inflammatory cytokines such as interleukin (IL)-1β, IL-6 and IL-12, as well as a reduction in anti-inflammatory regulatory T cells in adipose tissue20 and an increase in CD8+ T cells in adipose tissue.21 In addition, obesity may directly affect immune responses and antiviral T cells.22 For example, obesity is associated with more severe H1N1 influenza A pandemic infection in obese individuals with BMIs >35.23 Finally, during obesity there is a transition in adipose tissue macrophage polarisation from alternatively activated M2 macrophages to classically activated M1 macrophages that promote inflammation and insulin resistance.19 ,24 ,25 M2 macrophages are normally present in adipose tissue to maintain homeostasis. Thus, M2 macrophages act together with eosinophils, IL-4 and IL-13 to increase the thermogenic capacity of, and lipid utilisation in, adipose tissue, and mediate the conversion of white adipose tissue into beige fat, all of which ameliorates obesity.26 However, in obesity, there is a dramatic increase in the number of adipose tissue macrophages with a conversion of the M2 macrophages into the M1 form, which secrete TNF-α, IL-1β and IL-6. These cytokines then contribute to insulin resistance, pancreatic β cell death and type 2 diabetes.
The specific mechanisms that are responsible for the alteration of adipose tissue and the increase in M1 macrophages have only recently been understood. The mechanisms that translate obesity into chronic inflammation involve recently identified innate sensors of metabolic disturbance and tissue damage, and include inflammasomes. Inflammasomes are multiprotein platforms that oligomerise when activated by a variety of triggers, including products of tissue damage, for example, uric acid crystals, cholesterol crystals, extracellular ATP, low intracellular potassium concentrations as well as conserved pathogen characteristics such as foreign cytoplasmic double-stranded DNA, components of influenza A virus and some bacteria such as Neisseria and Salmonella. The activated inflammasome complexes, for example, NLR family, pyrin domain containing 3 (NLRP3), can then cleave procaspase 1, which can in turn cleave pro-IL-1β and IL-18 in many organs to drive inflammation. NLRP3, which can drive inflammation in auto-inflammatory disease such as gout, asbestosis, contact hypersensitivity and in certain viral and bacterial infections,27 appears to play an important role in obesity. This is because NLRP3 is activated by saturated fatty acids such as palmitate and stearate, as well as free cholesterol and cholesterol crystals, all known to be increased in obese patients, and by oxidative stress, which is also known to be present in adipose tissue in obesity (figure 1). Furthermore, islet amyloid polypeptide aggregates accumulate in diabetic pancreas tissue, a major hallmark of type 2 diabetes, also activates the NLRP3 inflammasome.28 During caloric excess, NLRP3 activation results in caspase 1 activation and secretion of IL-18, IL-6 and IL-1β, which is highly toxic to pancreatic β cells producing insulin.29 In support of this idea, studies in mice indicate that NLRP3 deficiency reduces the severity of high-fat diet-induced type 2 diabetes. Finally, neutralisation of IL-1 improves diabetes control in humans,30 confirming an important role for NLRP3 and IL-1 in type 2 diabetes.
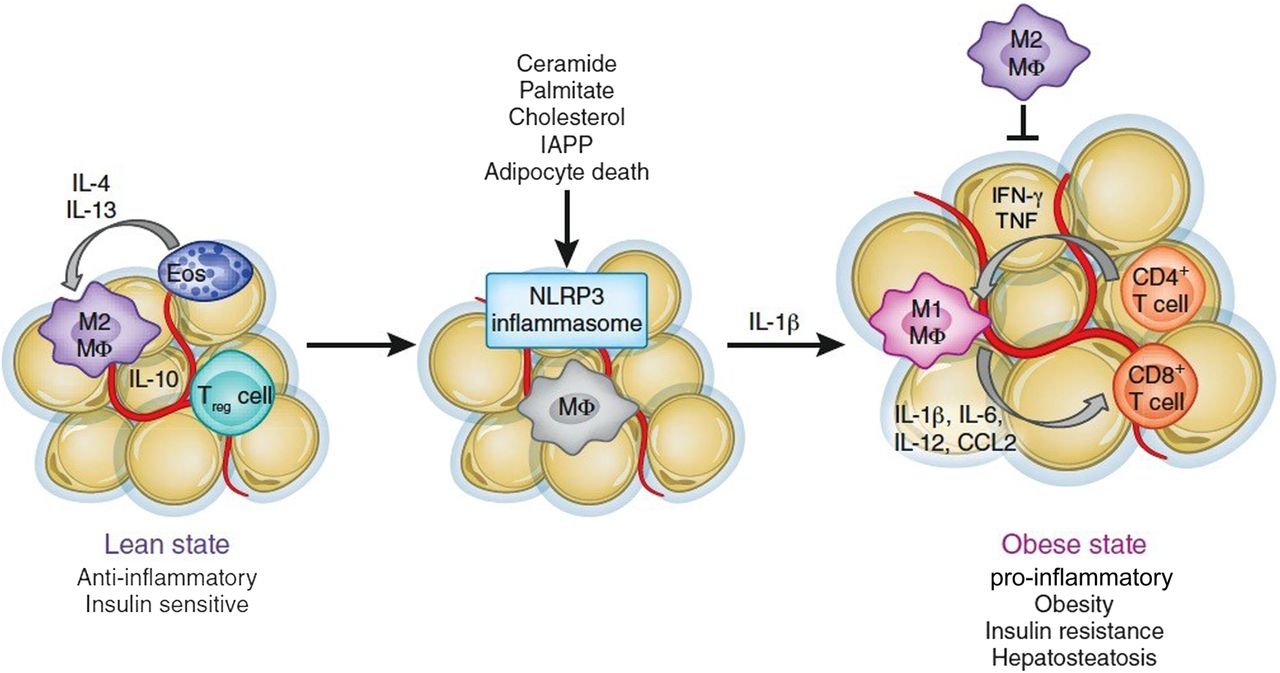
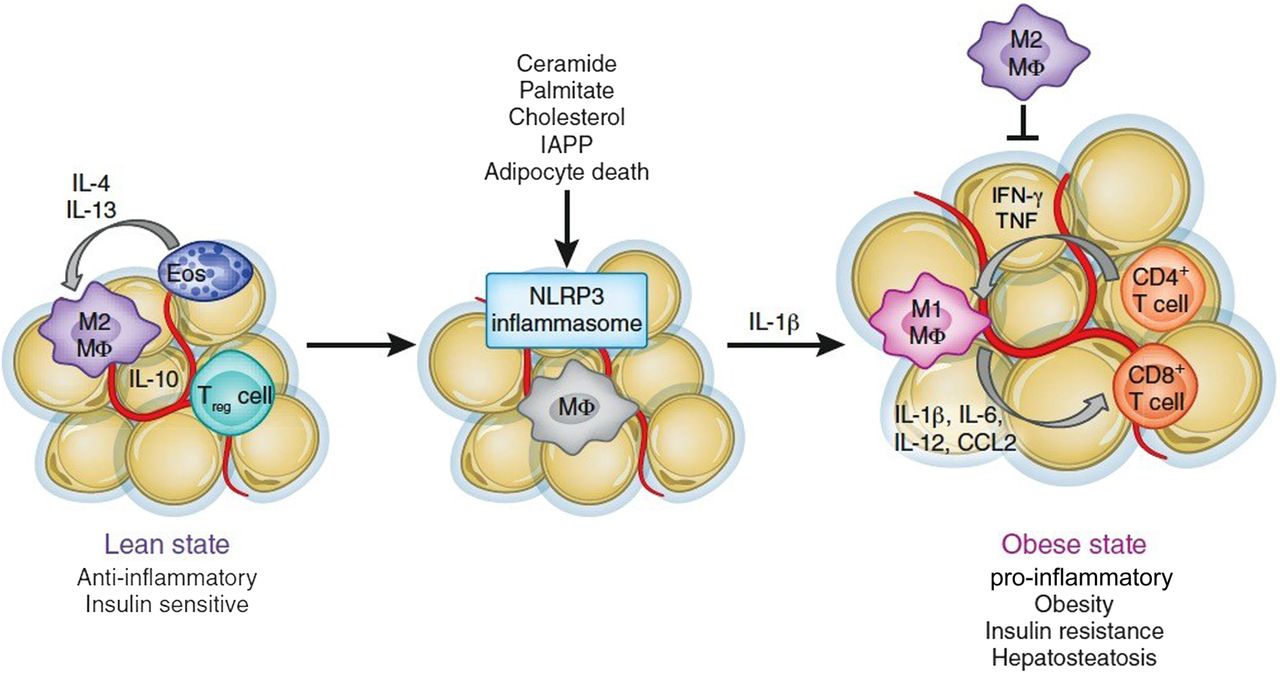{kind=link}
Inflammasomes fuel obesity-induced inflammation.19 IFN, interferon; IL, interleukin; IAPP, islet amyloid polypeptide; NLRP3, pyrin domain containing 3; TNF, tumour necrosis factor.
Obesity and asthma
While the understanding of obesity and type 2 diabetes has progressed rapidly in the past 5 years, the precise mechanisms that translate and drive obesity and inflammation into asthma have remained unclear. To understand the pathways that might lead to obesity-associated asthma, we established a mouse model, in which mice were fed a high-fat diet beginning at weaning at 3 weeks of age. Surprisingly, after consuming the high-fat diet for 12 weeks and becoming obese, the mice spontaneously developed AHR, without exposure to or priming with allergen.31 Moreover, the obesity-associated AHR was independent of adaptive immunity, as it occurred in obese Rag1−/− mice, which lack B and T cells, the mediators of immunological memory. This indicated that unlike most models of asthma, neither allergen-specific CD4 T cells nor allergen-specific IgE was required for the development of obesity-associated AHR. Furthermore, few eosinophils were found in the airways, consistent with the idea that obesity-associated asthma is allergen independent.
Since type 2 diabetes associated with obesity appears to be linked to the NLRP3 inflammasome, we asked if obesity-associated AHR also depended on the NLRP3 inflammasome. Surprisingly, AHR did not develop in obese Nlrp3−/− mice, whereas control wild-type obese mice developed robust AHR, indicating that in this model, NLRP3 might regulate the development of obesity-associated asthma,31 just as it appears to regulate type 2 diabetes. Moreover, the obesity-associated AHR also required IL-17A, since the obesity-associated AHR did not occur in obese Il17a−/− mice. In the lungs, the cells that produced IL-17 were primarily lymphocytes that did not express T-cell receptors or immunoglobulin receptors, but rather expressed CCR6, CD44, Sca-1, RORγt. These are markers of a newly described cell type, called type 3 innate lymphoid cells (ILC3s).
ILC3s belong to a family of recently described evolutionarily conserved, innate effector cells that can rapidly produce large amounts of cytokines.32 ,33 Type 1 ILCs, or ILC1s, produce interferon-γ and may facilitate inflammatory bowel disease, while type 2 ILCs, or ILC2s, produce IL-5 and IL-13 and may be involved in asthma phenotypes that are associated with eosinophilic inflammation. In contrast, ILC3s produce IL-17 and/or IL-22 and may be involved in autoimmune diseases. ILC3s respond to IL-1β, which is important, since NLRP3 activation induces IL-1β secretion, and since IL-1β protein was increased in the lungs of obese mice. Finally, in the lungs of obese mice, IL-1β was produced primarily by M1 macrophages, which were highly enriched, compared with that in the lungs of lean mice.
Moreover, treatment of mice with recombinant IL-1β by itself induced AHR, associated with a significant increase in ILC3s in the lungs, even in the absence of adaptive immunity or of type 2 cytokines.31 In addition, blockade of IL-1β signalling in the obese mice with an IL-1 receptor antagonist (anakinra) abolished obesity-induced AHR and reduced the number of ILC3s, demonstrating an important role of IL-1 in this model. Finally, examination of the bronchoalveolar lavage fluid from a small group of patients with severe asthma demonstrated the presence of ILC3s,31 suggesting that ILC3s producing IL-17 may play an important role in human asthma. M1 macrophages are known to produce IL-1β and IL-6, which is important, since increased production of IL-6 is another feature of metabolic dysfunction in obesity, and is associated with severe asthma in obese individuals.34 Together these studies suggest that the NLRP3 inflammasome, IL-1β and IL-17-producing ILC3s, all activated by the metabolic dysfunction in obesity, may comprise a non-type 2 mechanism that explains obesity-associated asthma, although additional mechanisms may be involved as well.
Treatment of obesity-associated asthma
As mentioned earlier, patients with obesity-associated asthma have disease that is refractory to standard asthma medications, including oral corticosteroids. This may be due to the fact that the metabolic innate immune mechanisms and ILC3s may be unresponsive to corticosteroids.35 ,36 While studies indicate that weight loss with bariatric surgery improves late-onset obesity-associated asthma, early onset obesity-associated asthma responds poorly to this treatment,12 and therefore additional treatments are critically needed for obesity-associated asthma, the prevalence of which is growing at an alarming rate.
Recently in 2014, mepolizumab (anti-IL-5 monoclonal antibody, GlaxoSmithKline) was approved by the Food and Drug Administration for the treatment of patients aged 12 years and older who have severe asthma with an eosinophilic phenotype. Mepolizumab, which has also been studied for other diseases such as COPD, hypereosinophilic syndrome, chronic rhinosinusitis with nasal polyps and eosinophilic esophagitis,37 is effective in reducing the number of eosinophils in the sputum and blood and in reducing asthma exacerbations and the need for treatment with systemic glucocorticosteroids.38 ,39 Interestingly, supervised cluster analysis of the clinical trial data showed that the subgroup cluster of patients that benefited the most from mepolizumab (cluster 4) was one characterised by raised blood eosinophils, obesity and a mean duration of disease of 18 years.40 We speculate that this subgroup (cluster 4) might actually include the patients described earlier with the early onset form of obesity-associated asthma. Indeed, patients in this cluster of obese patients with eosinophilia had a 67% reduction in exacerbations, compared with a 16%, 53% and 35% reduction in exacerbations in clusters 1, 2 and 3, respectively, suggesting that mepolizumab may be very effective for obese patients with asthma, presumably those with the early onset form.
For patients with the late-onset form of obesity-associated asthma, it is possible that interruption of some of the metabolic pathways that lead to asthma may be effective, although no current trials are in progress targeting IL-1, IL-6, IL-17, NLRP3, M1 macrophages or ILC3s in obese patients with asthma, as far as we know. An anti-IL-17 receptor A monoclonal antibody (brodalumab, Amgen/AstraZeneca) that prevents signalling by IL-17A and IL-25, has been studied in patients with moderate-to-severe asthma. However, while the outcomes between brodalumab-treated and placebo-treated patients were equivocal, it is possible that brodalumab may be effective only in a subpopulation of asthma patients that were not studied in this trial, for example, those with high IL-17 production, as is the case in obesity-associated asthma. Additional future studies with brodalumab or with other pharmaceutical agents, including biologics that neutralise IL-1 or IL-6 could potentially result in new and effective therapies for obesity-associated asthma. Moreover, because obesity alters the gut microbial community structure,41 ,42 and because gut bacteria can contribute to immunity, inflammation and disease, including obesity,43 examination of additional mechanisms by which the gut microbiome affects asthma in obese individuals may be promising. Clearly, as we learn more about the mechanisms by which obesity impacts asthma, we will be better able to develop effective treatments to address this serious unmet medical need.
In summary, asthma that is associated with obesity is a growing public health problem for which there are few effective treatments. Our understanding of the mechanisms that underlie this association is limited but growing. As this understanding broadens in the future, improved treatments will likely be examined and developed for this significant unmet medical need.
References
Footnotes
Funding National Institutes of Health (grant no. RO1 AI068085; U54 AI057159).
Competing interests DTU is an employee of Genetech Inc.
Provenance and peer review Commissioned; externally peer reviewed.