Article Text

Abstract
Background Diabetes is a risk factor for respiratory infection, and hyperglycaemia is associated with increased glucose in airway surface liquid and risk of Staphylococcus aureus infection.
Objectives To investigate whether elevation of basolateral/blood glucose concentration promotes airway Staphylococcus aureus growth and whether pretreatment with the antidiabetic drug metformin affects this relationship.
Methods Human airway epithelial cells grown at air–liquid interface (±18 h pre-treatment, 30 μM–1 mM metformin) were inoculated with 5×105 colony-forming units (CFU)/cm2 S aureus 8325-4 or JE2 or Pseudomonas aeruginosa PA01 on the apical surface and incubated for 7 h. Wild-type C57BL/6 or db/db (leptin receptor-deficient) mice, 6–10 weeks old, were treated with intraperitoneal phosphate-buffered saline or 40 mg/kg metformin for 2 days before intranasal inoculation with 1×107 CFU S aureus. Mice were culled 24 h after infection and bronchoalveolar lavage fluid collected.
Results Apical S aureus growth increased with basolateral glucose concentration in an in vitro airway epithelia–bacteria co-culture model. S aureus reduced transepithelial electrical resistance (RT) and increased paracellular glucose flux. Metformin inhibited the glucose-induced growth of S aureus, increased RT and decreased glucose flux. Diabetic (db/db) mice infected with S aureus exhibited a higher bacterial load in their airways than control mice after 2 days and metformin treatment reversed this effect. Metformin did not decrease blood glucose but reduced paracellular flux across ex vivo murine tracheas.
Conclusions Hyperglycaemia promotes respiratory S aureus infection, and metformin modifies glucose flux across the airway epithelium to limit hyperglycaemia-induced bacterial growth. Metformin might, therefore, be of additional benefit in the prevention and treatment of respiratory infection.
- Airway Epithelium
- Bacterial Infection
- COPD Exacerbations
- Respiratory Infection
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/3.0/ and http://creativecommons.org/licenses/by-nc/3.0/legalcode
Statistics from Altmetric.com
Introduction
Hyperglycaemia is associated with increased risk of Staphylococcus aureus infection in the respiratory tract,1 ,2 Hyperglycaemia is a risk factor for nasal colonisation with S aureus3 increased pathogen load in cystic fibrosis (CF),4 ,5 increased exacerbation frequency and poor outcomes from exacerbations in people with chronic obstructive pulmonary disease (COPD).1 ,6 In intensive care, intubated patients with hyperglycaemia had more S aureus in their bronchial aspirates and this was associated with increased glucose concentration in the layer of fluid that lines the airways (airway surface liquid (ASL)).2
Glucose concentration in human ASL is normally ∼0.4 mM, 12.5 times lower than that of blood.7–9 ASL glucose concentrations are raised in people with respiratory disease, including acute viral rhinitis,10 COPD1 and CF.9 ASL glucose concentrations are also increased in experimental11 and diabetic hyperglycaemia10 and are further increased in people with both respiratory disease and diabetes mellitus.5 ,9
We developed an in vitro model of airway glucose homeostasis and showed that under normal conditions glucose predominantly diffuses from blood/interstitial fluid across the respiratory epithelium into the ASL via paracellular pathways, and this is limited by epithelial permeability.8 ,12–14 Uptake via apical and basolateral GLUT transporters also restricts glucose accumulation in ASL,12–15 and rapid metabolism of glucose helps to maintain low intracellular glucose concentrations. This provides a driving force for glucose uptake and limits the transcellular transport of glucose, leading to equilibrium between ASL and intracellular glucose concentrations.13 ,14 In this model, increasing the diffusion gradient for glucose across the epithelium (eg, hyperglycaemia) and increasing paracellular diffusion of glucose via reduced transepithelial resistance (RT) (eg, inflammation) would result in raised ASL glucose concentration.12 ,13 As S aureus growth is promoted by glucose in microbial culture5 we sought to demonstrate experimentally that hyperglycaemia promotes growth of S aureus in the airway through modification of these processes.
Metformin is used in the clinical treatment of type II diabetes to reduce blood glucose. However, it has also been reported to decrease permeability,16–19 exhibit anti-inflammatory effects16 ,20 and increase glucose uptake in epithelial tissues.21–23 We therefore investigated whether metformin could modify these processes and contribute to a change in epithelial glucose homeostasis and S aureus growth.
Methods
Bacterial culture
S aureus strains 8325-4 and JE2 and Pseudomonas aeruginosa strain PA01 were used for the experiments. A single colony of each organism/strain was incubated overnight at 37°C in RPMI medium. The following day fresh medium was added and culture grown to log phase, OD600 of 0.5 (about 2×107 colony-forming units (CFU)/ml).
Mouse infection model
To assess the effects of glucose on bacterial growth in the airways we developed an animal model of S aureus lung infection, based on previous studies.24–26
Wild-type (WT) or db/db (leptin receptor-deficient) C57BL/6, 6–10 weeks old, were obtained from Harlan (Titlarks Acre, UK). Where stated, mice were treated with 800 μg metformin (40 mg/kg) intraperitoneally every day from day 2 before infection. Blood glucose was measured at the same time each day. Mice were inoculated intranasally with 1×107 CFU of live or heat-killed S aureus 8325-4 in 100 μl. Infectious doses were confirmed by plating out serial dilutions. Twenty-four hours after infection, mice were culled. Bronchoalveolar lavage (BAL) fluid was recovered by inflating the lungs five times with 1 ml of phosphate-buffered saline (PBS) via an excision in the trachea. Dilutions of BAL fluid were plated out onto brain heart infusion agar plates to count the CFU. BAL cells were counted by trypan blue exclusion, centrifuged onto glass slides and stained with haematoxylin and eosin to identify neutrophils. Interleukin 6 (IL-6) was measured in BAL supernatants by ELISA using manufacturer's protocols (R&D Systems).
The features of this model are an acute weight loss associated with neutrophilic influx into the airways, which is dependent upon live bacteria and not seen with heat-killed bacteria. Despite the use of inbred mice and a single bacterial strain, there is some variance between degrees of neutrophilia and cell recovery which may be due to other factors such as cage dominance, which can affect the response to infection. Having established this model in WT mice, it gave us a platform to study the effect of hyperglycaemia on airway infection (see online supplementary figures 1A–D).
Airway epithelial cell culture
H441 epithelial cells were grown on permeable membrane supports (Transwells, Corning, USA) at the air–liquid interface to form confluent polarised monolayers, as previously described.12 Monolayers were pretreated with 0, 0.03, 0.3 or 1 mM metformin (added to the basolateral medium) 18 h before apical inoculation with S aureus. Monolayer RT was measured using an epithelial voltohmmeter (WPI, UK). The glucose concentration of the surface liquid overlying H441 monolayers was measured using glucose oxidase analyser (Analox Instruments, UK), as previously described.12 Primary human bronchial epithelial (HBE) cells were obtained from endobronchial brushings and cultured as previously described.27 Cells were transferred onto transwell permeable supports and grown at the air–liquid interface to form confluent monolayers.28
In vitro co-culture model
Bacterial cultures were diluted in glucose-free RPMI medium, and 5×105 CFU (8325-4 and JE2) or 1×108 CFU (PA01) in 50 μl was applied to the apical surface of H441 or HBE monolayers. The basolateral side of the airway epithelial–bacterial co-cultures was bathed in Krebs salt solution (in mM): NaCl, 117; NaHCO3, 25; KCl, 4.7; MgSO4, 1.2; KH2PO4, 1.2; CaCl2, 2.5 (equilibrated with 5% CO2 to pH 7.3–7.4). Hyperglycaemia was modelled by applying basolateral glucose concentrations of 10, 20 and 40 mM. For co-cultures grown in bilateral glucose, S aureus cultures were supplemented with 10 mM glucose. Co-cultures were placed in a CO2 incubator at 37°C for 7 h, then each was homogenised and CFU calculated by plating out serial dilutions.
S aureus culture in H441 epithelial fluid secretions
Apical secretions were obtained from cells grown in the presence of 10, 20, 40 mM basolateral glucose, with or without 1 mM metformin (18 h), in the presence or absence of heat-killed S aureus to mimic proinflammatory stimuli. Glucose-free Krebs salt solution (100 μl) was applied to the apical surface of H441 monolayers and removed after 7 h. The glucose concentration of each sample was normalised to 10 mM to allow detection of any difference in growth due to factors other than glucose. 5×105 CFU of S aureus were applied to 100 μl of each fluid secretion sample in a 96-well plate and incubated for 7 h. S aureus CFU were calculated as above.
Co-culture cytokine analysis
In parallel 7 h co-cultures, the apical fluid was removed for cytokine analysis using an in-house luminex with reagents from R&D Systems, UK.
Paracellular L-glucose flux and D-glucose uptake experiments
Paracellular movement of glucose across H441 monolayers was measured by analysis of radiolabelled [14C]-L-glucose transepithelial flux. Experiments were initiated by adding 1.0 ml Krebs salt solution containing 1.0 μCi of [14C]-L-glucose plus 10 mM radiolabelled equivalent glucose to the basolateral side of the transwells and 0.1 ml of glucose-free Krebs solution to the apical side. Apical and basolateral samples were taken after 1 h and the concentration of radiolabelled glucose was analysed using a scintillation counter.
Transepithelial movement of [14C]-L-glucose across ex vivo mouse tracheas was measured in an Ussing chamber, as previously described.12
[14C]-D-Glucose uptake experiments on H441 monolayers were performed, as previously described.12
Chemicals and reagents
All chemicals and reagents were obtained from Sigma, Poole, UK unless otherwise stated.
Statistical analysis
Values are reported as mean±SEM. Statistical analysis was performed using analysis of variance and post hoc Bonferroni multiple comparison or Student t test. p Values<0.05 were considered significant.
Results
Hyperglycaemia increases S aureus growth in the airways of infected mice
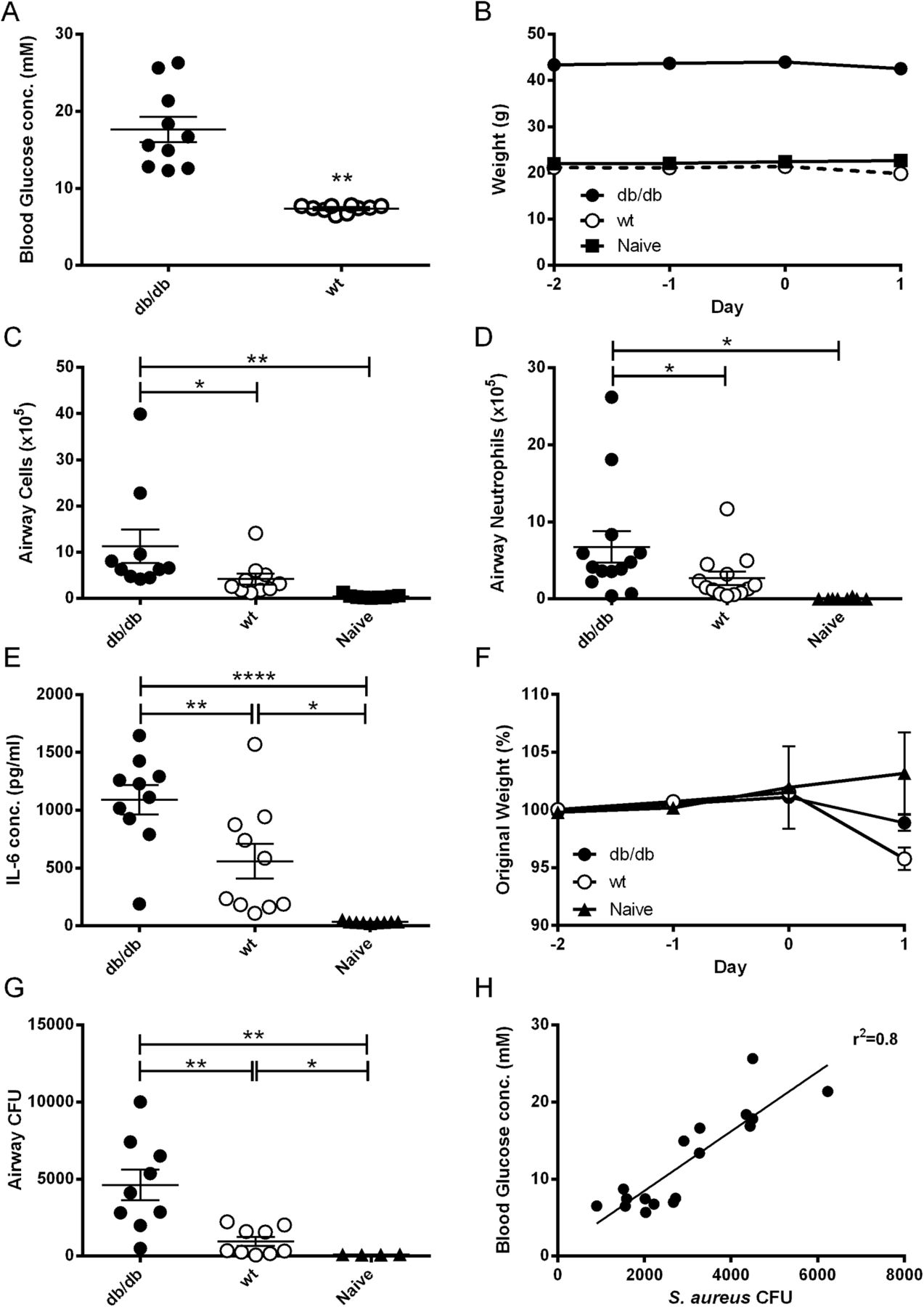
Mice deficient for the leptin receptor (db/db) exhibited significantly higher blood glucose concentrations (17.7±1.6 vs 7.4±0.1 mM; p<0.01; n=10; figure 1A) and were heavier (p<0.001; n=10; figure 1B) than WT mice. Twenty-four hours after infection with S aureus 8325-4, there were more inflammatory cells, neutrophils and cytokines (IL-6) present in the airways of db/db than WT mice or naïve mice (non-infected WT mice) (p<0.05; n=10–13; figure 1C, D and E). All infected mice lost weight after infection (figure 1F). Crucially, db/db mice had significantly more bacteria in their airways after infection than WT mice (p<0.01; n=9; figure 1G), and there was a direct correlation between blood glucose and CFU (r2=0.8, p<0.0001; n=18; figure 1H) supporting clinical observations in humans.

Comparison of Staphylococcus aureus numbers in BAL fluid of infected db/db and wild-type (WT) mice. Leptin receptor deficient (db/db) or WT C57BL/6 mice were inoculated with 107 CFU of S aureus strain 8325-4 intranasally. (A) Blood glucose concentration on the day of infection. (B) Weight measured over the time of the experiment. (C) Number of cells; (D) number of neutrophils; (E) IL-6 concentration in BAL fluid on day 1 after infection. (F) Percentage weight loss measured during the experiment. (G) Bacterial CFU recovered from BAL fluid on day 1 after infection. Individual mice are shown as data points, the horizontal bars represent mean ± SEM of n=9–13 (pooled experiments). *p<0.05, **p<0.01, ****p<0.0001. (H) Correlation of blood glucose concentration with CFU recovered from BAL fluid, n=18. BAL, bronchoalveolar lavage; IL, interleukin.
Metformin reduces airway S aureus growth in diabetic mice without lowering blood glucose
Metformin treatment significantly reduced bacterial load in the airways of db/db mice compared with PBS treatment (p<0.01; n=9; figure 2A), to levels similar to those seen in WT mice (p>0.05). Surprisingly, this reduction in airway S aureus numbers occurred despite metformin having no significant effect on blood glucose levels over this period of time (p>0.05; n=10; figure 2B). Metformin had no effect on S aureus load in WT mice (p>0.05; n=9; figure 2A). There was no difference in total cell (n=10, figure 2C) or neutrophil (n=9, figure 2D) recruitment to the airways or BAL IL-6 (n=14, figure 2E) concentration after infection, between metformin-treated and PBS-treated mice.
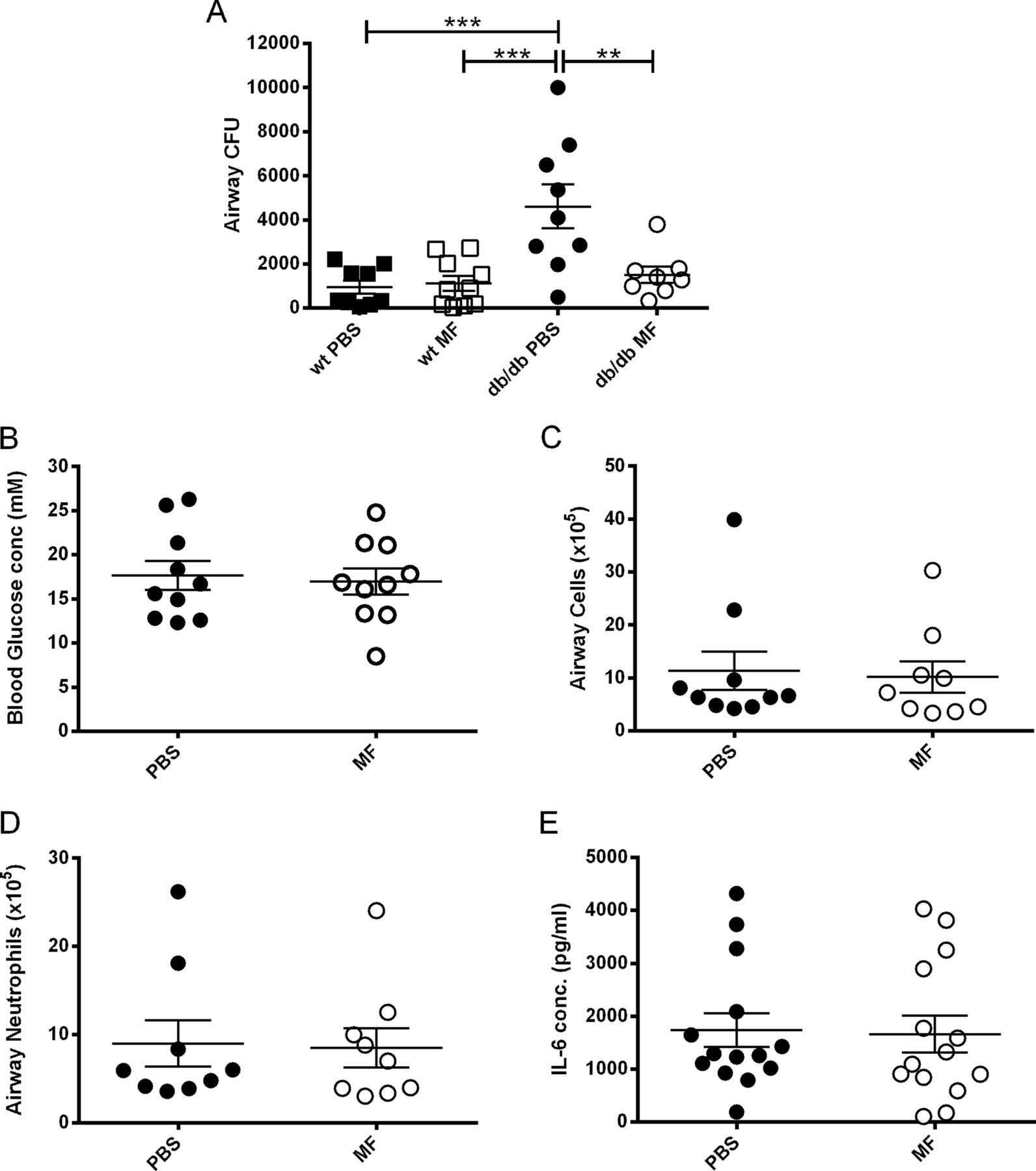
Metformin (MF) reduces airway Staphylococcus aureus numbers in hyperglycaemic mice, without lowering blood glucose. Leptin receptor deficient (db/db) mice were inoculated with 107 CFU of S aureus strain 8325-4 intranasally. Before infection mice were treated with 200 μl 40 mg/ml metformin or phosphate-buffered saline (PBS) intraperitoneally. (A) Comparison of bacterial CFU recovered from BAL fluid of untreated and metformin treated wild-type (WT) and db/db mice on day 1 after infection. (B) Blood glucose concentration on the day of infection. (C) Number of cells; (D) number of neutrophils; (E) interleukin 6 (IL-6) concentration in BAL fluid on day 1 after infection. Individual mice are shown as data points, the horizontal bars represent mean ± SEM of n=9–14 ± SEM (pooled experiments), **p<0.01, ***p<0.001. BAL, bronchoalveolar lavage.
Hyperglycaemia increases apical S aureus growth in airway epithelia–bacteria co-cultures
Increasing basolateral glucose from 10 to 20 or 40 mM in H441 airway epithelial co-cultures produced a stepwise increase in apical S aureus 8325-4 growth (2.7±0.4×107 to 13.0±2.4×107 CFU/ml in 10 and 40 mM, respectively, p<0.01; n=9, figure 3A). Similarly, apical S aureus 8325-4 growth was also increased by increasing basolateral glucose from 10 mM to 20 mM in primary HBE monolayers (1.8±0.2×107 to 2.5±0.2×107 CFU/ml respectively; p<0.05; n=4, figure 3B). Application of 10 mM glucose to apical and basolateral solutions significantly increased apical S aureus growth compared with 10 mM (basolateral only) control (p<0.05; n=12; figure 3A). No S aureus colonies were seen under any condition, in basolateral fluid suggesting that the bacteria did not cross the epithelial barrier over the 7 h time course. Elevation of basolateral glucose promoted the growth of S aureus JE2 (p<0.05; n=3; figure 3C). Furthermore, the growth of P aeruginosa (PA01), was also promoted by increasing basolateral glucose from 10 to 40 mM (p<0.05; n=6; figure 3D).

Staphylococcus aureus growth across airway epithelial monolayers is dependent on basolateral glucose levels and is inhibited by metformin. Airway epithelia–bacteria co-cultures were grown in the presence of varying concentrations of basolateral glucose (10, 20, 40 mM glucose) or 10 mM bilateral glucose (10/10) without (black bars) or with pretreatment with metformin (1 mM; 18 h pre-treatment; hatched bars). Bacterial CFU were measured 7 h after infection (percentage compared with growth in the presence of 10 mM basolateral glucose). (A) H441 and S aureus 8325-4 co-culture, n=9–12; (B) primary human bronchial epithelial cells and S aureus 8325-4 co-culture, n=4; (C) H441 and S aureus JE2 co-culture, n=4; (D) H441 and Pseudomonas aeruginosa co-culture, n=12. *p<0.05, ****p<0.0001, †p<0.05 compared to 10mM glucose pretreated with metformin.
Epithelial cells secrete a number of proteins and substances on their apical surface.29 Some have antimicrobial properties; others could promote the growth of bacteria. Therefore, we tested whether basolateral glucose-induced changes in H441 fluid secretions influenced apical bacterial growth. No effect on S aureus growth was seen in fluid secretions (normalised for glucose concentration) produced by different hyperglycaemic conditions in the absence (figures 4A) or presence of heat-killed bacteria (figure 4B). These data are consistent with basolateral glucose, not other glucose-dependent factors, as a determinant of apical S aureus growth.

Staphylococcus aureus growth in fluid secretions from H441 monolayers. (A) S aureus growth over 7 h in fluid secretions (with glucose concentration normalised to 10 mM) obtained from the apical surface of H441 epithelial monolayers after exposure to different concentrations of basolateral glucose (10, 20, 40 mM glucose; black bars) or 40 mM glucose + metformin (MF) pretreatment (1 mM; 18 h pretreatment; hatched bars), n=4, or (B) in the presence of heat-killed bacteria, and different concentrations of basolateral glucose (10, 20, 40 mM glucose; black bars) or 40 mM glucose + metformin pretreatment (hatched bars) n=4. (C) S aureus growth over 7 h across H441 airway epithelial monolayers, after pretreatment with different concentrations of metformin (all in the presence of 10 mM basolateral glucose), n=5. (D) S aureus growth in culture media only (without cells) containing varying concentrations of metformin, n=4. All data are shown as percentage of control *p<0.05, ***p<0.001, ****p<0.0001.
Metformin inhibits hyperglycaemia-induced S aureus growth in airway epithelia–bacteria co-cultures
Metformin pretreatment significantly reduced apical S aureus growth at all basolateral glucose concentrations (p<0.05; n=9; figure 3A). However, metformin had no significant effect on S aureus growth in co-cultures where glucose was at a concentration of 10 mM in both apical and basolateral solutions (n=12; figure 3A). Metformin reduced the growth of S aureus in a dose-dependent manner (figure 4C). The lowest dose, 30 μM metformin, reduced apical S aureus growth in the presence of 10 mM basolateral glucose by 29±10% (p<0.05; n=4).
Metformin had no direct effect on S aureus growth in bacterial culture at any concentration tested (n=4; figure 4D).
Granulocyte-macrophage colony-stimulating factor (GM-CSF), interferon γ (INFγ), interleukins (IL-1α, IL-6), monokine induced by gamma interferon (MIG/CXCL9) and transforming growth factor β (TGFβ) concentrations were raised in the ASL of H441 monolayers after the addition of apical S aureus (p<0.0001; n=12; figure 5A–F). All except GM-CSF, were significantly reduced by metformin (p<0.0001; n=12). The concentration of basolateral glucose had no effect on cytokines produced in the presence or absence of S aureus or in the presence and absence of metformin (data not shown).

Metformin reduces Staphylococcus aureus-induced cytokine secretion. The concentration (pg/ml) of (A) GM-CSF; (B) IFNγ; (C) IL-1α, (D) IL-6, (E) MIG/CXCL9; (F) TGF-β was measured in the fluid (50 µl) covering the apical surface of H441 monolayers after 7 h in the absence and presence of apical S aureus,±metformin pretreatment, n=12. ****p<0.0001. GM-CSF, granulocyte-macrophage colony-stimulating factor; IFN, interferon; IL, interleukin; MIG, monokine induced by gamma interferon; TGF, transforming growth factor.
Metformin reduces ASL glucose and transepithelial glucose flux
ASL glucose
Glucose concentration of the ASL covering the surface of H441 airway epithelial monolayers increased when basolateral glucose concentration was raised from 10 to 40 mM (p<0.0001; n=4; figure 6A). Metformin treatment produced a significant decrease in ASL glucose from 2.3±0.3 to 1.3±0.3 mM in 40 mM basolateral glucose (p<0.05; n=4).
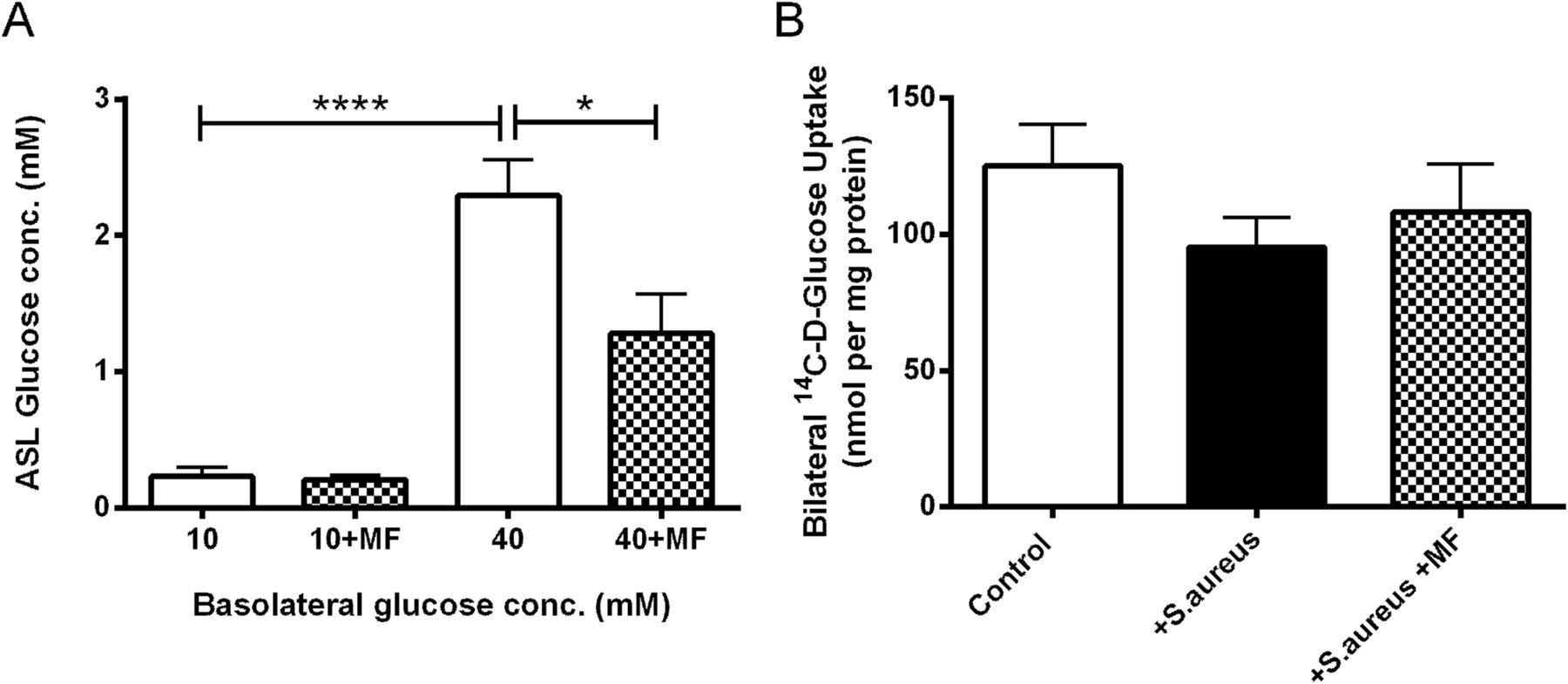
Metformin (MF) lowers airway surface liquid (ASL) glucose concentrations in vitro, but has no effect on epithelial glucose uptake. (A) Glucose concentration of ASL in the presence of 10 and 40 mM basolateral glucose without (open bars) or with metformin pretreatment (hatched bars), n=4. (B) Bilateral glucose uptake assessed using radiolabelled glucose (14C-d-glucose) applied to both the apical and basolateral surfaces, n=7. *p<0.05, ****p<0.0001.
Cellular glucose uptake
D-Glucose uptake by airway epithelial monolayers was not significantly affected by the presence of S aureus in either control or metformin-treated conditions (p>0.05; n=8; figure 6B).
Transepithelial resistance (RT) and paracellular glucose flux
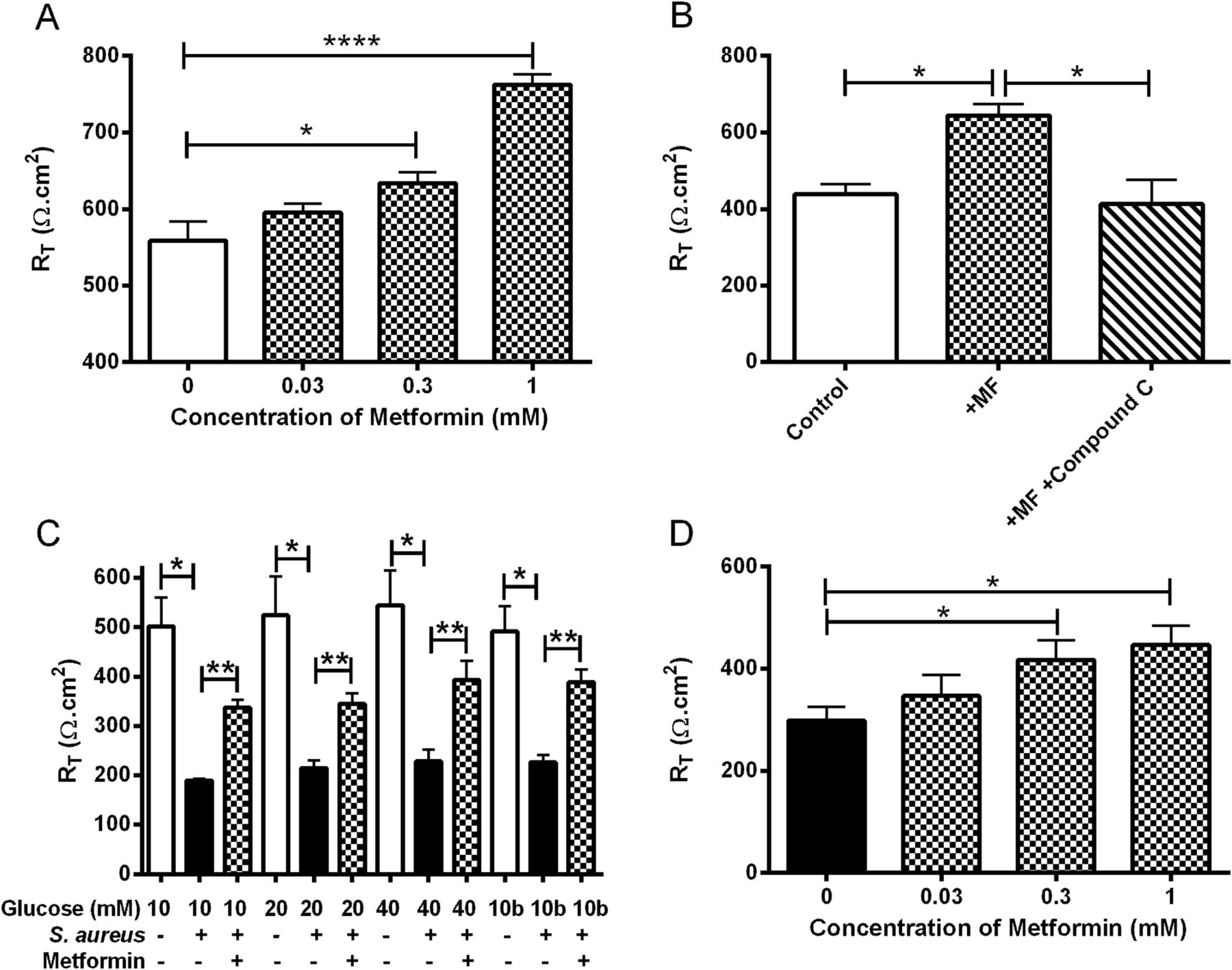
Metformin induced a dose-dependent increase of RT in H441 monolayers. Addition of 1 mM metformin increased monolayer RT from 559±25 to 762±14 Ω.cm2 (p<0.0001; n=15; figure 7A), which was prevented by pretreatment with the AMP activated protein kinase (AMPK) inhibitor compound C (p<0.05; n=3; figure 7B). The addition of S aureus to the apical surface produced a significant decrease in RT at all glucose concentrations tested (p<0.001; n=6; figure 7C). Metformin treatment attenuated the fall in RT caused by apical S aureus (p<0.01; n=6; figure 7C) in a dose-dependent manner (figure 7D). The concentration of glucose bathing the cells had no significant effect on RT in either the presence or absence of apical S aureus (p>0.05; n=6).

Metformin (MF) reverses the reduction of transepithelial resistance caused by Staphylococcus aureus. The effect of metformin (1 mM, 18 h pretreatment, hatched bars) on H441 airway epithelial monolayer RT (Ω cm2) in the (A) absence (n=15) and (B) presence of AMPK inhibitor compound C (n=3; 80 µM pretreatment for 1 h before addition of metformin). (C) Effect of S aureus addition (7 h; black bars) on untreated (open bars) and metformin pretreated (hatched bars) H441 monolayer RT, n=6. (D) Effect of different concentrations of metformin on H441 monolayer RT after 7 h co-culture with apical S aureus, n=5. *p<0.05, **p<0.01, ****p<0.0001. AMPK, activated protein kinase; RT, transepithelial electrical resistance.
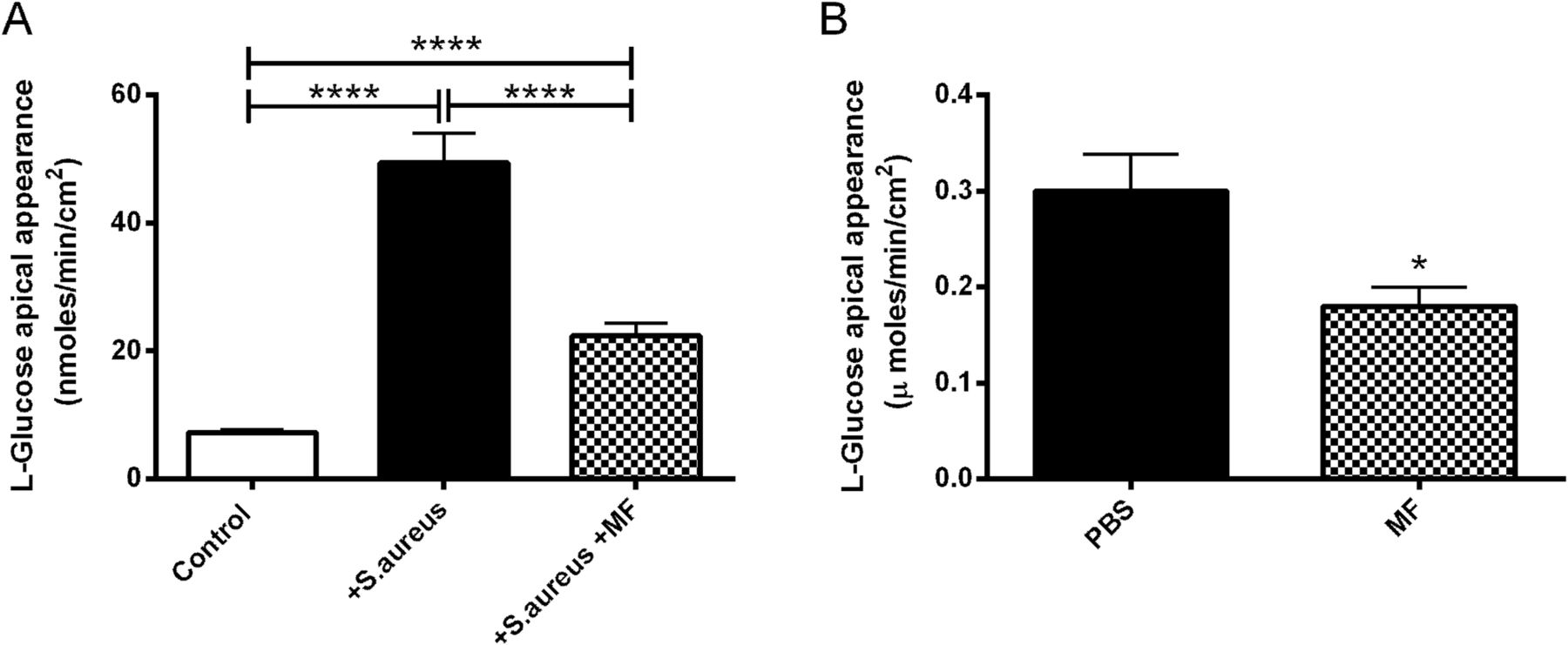
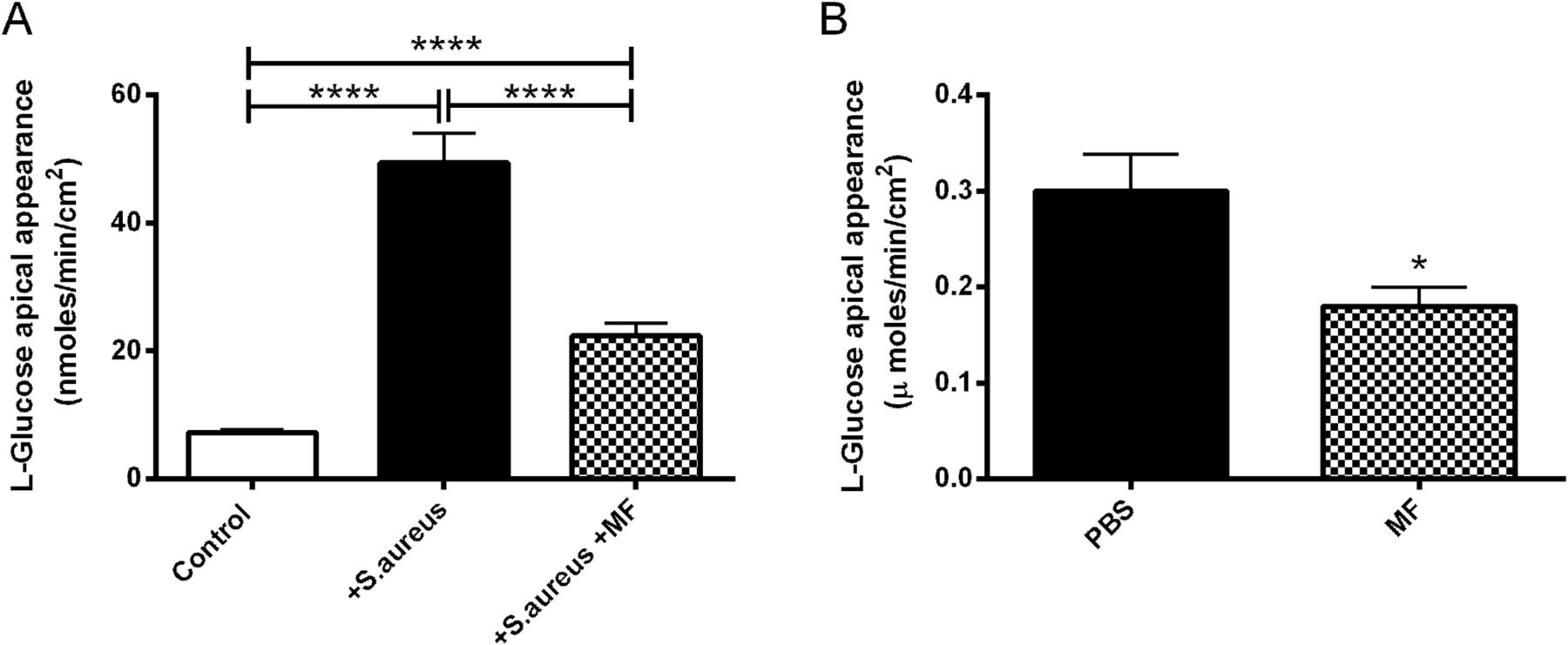
The decrease in RT evoked by apical S aureus gave rise to an increase in the rate of basolateral-to-apical paracellular L-glucose flux, from 7.3±0.5 to 49.3±4.6 nmoles/min/cm2 (10 mM basolateral glucose; p<0.0001; n=8; figure 8A). Metformin significantly impaired the rise in L-glucose flux (p<0.0001; n=8; figure 8A). Similarly, metformin treatment significantly reduced the L-glucose flux across infected ex vivo mouse tracheas by 40±5% (p<0.05; n=3; figure 8B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Metformin (MF) reverses the effect of Staphylococcus aureus infection on paracellular glucose flux across airway epithelium. H441 monolayers with and without 1 mM metformin pretreatment, were infected with S aureus. (A) Paracellular glucose flux measured by adding radiolabelled l-glucose to the basolateral surface and monitoring its appearance at the apical surface, n=8. (B) Transepithelial l-glucose flux across ex vivo murine tracheas from S aureus-infected mice pretreated with phosphate-buffered saline (PBS) or metformin, n=3. *p<0.05, ****p<0.0001.
Discussion
We have shown for the first time that the S aureus load was increased after inoculation into the airways of hyperglycaemic db/db mice compared with mice with normal blood glucose concentrations. These findings were replicated in an in vitro human airway epithelial–S aureus co-culture model, where elevation of basolateral glucose promoted apical growth of S aureus in a dose-dependent manner. We used S aureus 8425-4 in our experiments as it is the standard laboratory strain of S aureus for genetic and virulence studies. It belongs to an important lineage of virulent S aureus and methicillin-resistant S aureus (MRSA) isolates.30 Glucose also promoted the apical growth of JE2, a strain closely related to the common community-associated MRSA clone circulating in the USA (USA300), indicating that the effect is not strain-specific.31 In addition, elevation of basolateral glucose promoted the growth of unrelated, but glucose-using, bacteria P aeruginosa (PA01), indicating relevance of this effect to other respiratory pathogens.
In humans, cross-sectional studies showed an association of hyperglycaemia with respiratory S aureus infection. People with diabetes mellitus have increased risk of nasal S aureus colonisation.3 Intubated patients in intensive care with acute hyperglycaemia have increased S aureus isolated from bronchial aspirates.1 ,2 People admitted to hospital with COPD exacerbations who have hyperglycaemia are more likely to have S aureus on sputum culture than those with normoglycaemia.1 In CF, S aureus is an important pathogen, with 22% of adults (≥16 years) having chronic persistent infection.32 In a cohort study, 8–21-year-old patients with CF and persistent MRSA infection were more likely to have related diabetes mellitus than those who were MRSA negative.33 Our experimental findings support a direct causative effect of hyperglycaemia underlying this association. Hyperglycaemia preceded bacterial growth both in vivo and in vitro. The effects of elevated basolateral glucose on bacterial growth could be replicated by direct application of glucose to the apical compartment in vitro. Effects were not seen when bacteria were cultured in apical secretions from cells cultured under different glycaemic conditions, after normalisation for glucose concentrations, indicating that the effects were not mediated by changes in other factors, such as mucin secretion, metabolic products or protein glycation.34
Addition of S aureus to the luminal surface of the epithelium in vitro and in vivo induced a reduction in RT and an increase in basolateral-to-apical glucose flux. This is consistent with our previous observations where proinflammatory conditions decreased RT and increased glucose flux,12 and S aureus increased epithelial permeability in mouse trachea.35 These data therefore support a model whereby hyperglycaemia and the effect of S aureus on RT increase transepithelial glucose flux in vitro and in vivo. As S aureus uses glucose as a nutrient source, we propose that these changes promote its growth by increasing nutrient availability.5
A key finding of this study was that, metformin (40 mg/kg in mice; 30 μM in cell culture) attenuated glucose-induced S aureus growth both in vivo and in vitro, at clinically-relevant doses and concentrations (in humans, the usual metformin dose is about 15–30 mg/kg/day and plasma concentrations are ∼5–20 μM36 ,37).
Metformin appears to prevent apical/luminal growth of S aureus through an effect on epithelial permeability, with restoration of RT and reduced glucose flux in the presence of S aureus. We propose that this action, at least in part, inhibits S aureus growth by limiting apical glucose availability. How metformin increases RT in the airway epithelium has not been described. In renal epithelial cells, metformin directly modified epithelial tight junctions via activation of AMPK.18 ,19 Our observation that compound C (an AMPK inhibitor) prevented the elevation of RT by metformin in H441 monolayers provides evidence for a similar pathway in airway epithelium. Metformin could indirectly modify epithelial permeability by suppressing the action of proinflammatory cytokines.38 However, metformin increased RT in the absence of proinflammatory agents or S aureus. Furthermore, although metformin prevented S aureus-induced proinflammatory cytokine secretion in vitro, we were unable to determine a significant effect on the inflammatory response in vivo, even though we detected a decrease in glucose flux across ex vivo trachea with metformin.
Metformin thus appears to reduce bacterial growth in the airway by reducing epithelial permeability and limiting glucose flux into the airway. We were unable to confirm that metformin reduced luminal glucose concentrations in mouse airway. Measurement of glucose in mouse ASL is extremely challenging and BAL mostly samples distal lung fluid, rather than airway secretions. Glucose homeostasis differs considerably between the airways and distal lung.39 Hence, BAL fluid may not provide insights into processes occurring in the proximal airways. Additionally, epithelial damage during infection or BAL fluid collection may cause glucose leak into ASL from blood and interstitial fluid, leading to contamination of samples. Rapid glucose use by bacterial and inflammatory cells may also affect ASL glucose concentrations.
Metformin had no direct effect on S aureus growth and critically, the in vivo effect was independent of any observed changes in blood glucose concentration. Metformin was shown to have blood glucose-lowering effects in db/db mice,40 but our treatment protocol was much milder (40 mg/kg compared with 300 mg/kg) and shorter (2 days compared with 2 weeks).40 Metformin similarly did not affect basolateral glucose concentration in our in vitro studies (data not shown).
Metformin reduced S aureus numbers in db/db mice but not in WT mice, providing further evidence that metformin did not inhibit bacterial growth via alternative effects on—for example, airway ion transport or mucus secretion in vivo. Additionally, metformin did not increase airway epithelial glucose uptake, in contrast to its role in skeletal muscle where it promotes glucose uptake.41
In conclusion, S aureus growth in the airways is promoted by hyperglycaemia, and clinically-relevant concentrations of metformin significantly reduced this glucose-dependent growth. Metformin decreased transepithelial glucose permeability, at least in part, via activation of AMPK. We propose that this restricts glucose availability to S aureus, limiting bacterial growth. As bacterial resistance to antimicrobial increases, and there are few new agents in development, there is increasing need for alternative anti-infective strategies. This new action of metformin, in addition to its blood glucose lowering effects, may be of therapeutic benefit in the prevention of respiratory infection, particularly with glucose-using pathogens such as S aureus and in diseases such as COPD and CF where hyperglycaemia is associated with accelerated pulmonary decline. We are now undertaking a clinical trial of metformin for COPD exacerbations (ISRCTN66148745), focusing on the clinical utility of this drug for respiratory infection.
Key messages
-
What is the key question?
-
How does hyperglycaemia promote respiratory infection with Staphylococcus aureus?
-
What is the bottom line?
-
We show that elevation of blood glucose in vivo and basolateral glucose in vitro promotes the growth of S aureus in the airway and is associated with increased glucose flux across the airway epithelium. These data provide the first mechanistic insights into the association between hyperglycaemia and increased risk of respiratory S aureus infection in patients with acute illness or chronic lung disease.
-
Why read on?
-
We also show that pretreatment with the antidiabetic drug metformin limits the growth of S aureus in the lung. We provide new evidence that metformin reduces glucose flux into the lung by reducing transepithelial permeability, independently of its known blood glucose-lowering effects. This highlights a new potential therapeutic benefit of metformin in the prevention of lung infection, particularly in patients with respiratory disease and hyperglycaemia.
Acknowledgments
The authors would like to thank Dr Malcolm Brodlie, Newcastle University, UK for providing human bronchial epithelial cells and Professor Alain Filloux, Imperial College London, UK, for providing P aeruginosa.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figure
Footnotes
-
Contributors DLB, EHB and JPG conceived and designed the experiments. JPG performed the in vitro and ex vivo experiments and analysed the data. JPG and DBL wrote the paper. JST designed and performed the in vivo experiments, analysed the data and contributed to writing and editing the manuscript. SN and SG contributed to the in vitro experiments. JAL and GMK provided bacteria and microbial reagents.
-
Funding This work was supported by the Wellcome Trust (grant No: 088304/Z/09/Z) an enterprise grant from St George's University of London and a Wellcome Trust ISSF grant to Imperial College London.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval All animal work was approved by a local ethical review board, as described by the Animals (Scientific Procedures) Act 1986, and the work was licensed by the UK Home Office. Bronchial brushings, from which human bronchial epithelial cells were derived, were performed with ethical approval from the local research ethics committee, REC reference number: 2001/179.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Open Access This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 3.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/3.0/