Article Text

Abstract
Idiopathic pulmonary fibrosis (IPF) is a progressive and fatal lung disease with unclear aetiology and poorly understood pathophysiology. Although plasma levels of circulating cell-free DNA (ccf-DNA) and metabolomic changes have been reported in IPF, the associations between ccf-DNA levels and metabolic derangements in lung fibrosis are unclear. Here, we demonstrate that ccf-double-stranded DNA (dsDNA) is increased in patients with IPF with rapid progression of disease compared with slow progressors and healthy controls and that ccf-dsDNA associates with amino acid metabolism, energy metabolism and lipid metabolism pathways in patients with IPF.
- idiopathic pulmonary fibrosis
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
Idiopathic pulmonary fibrosis (IPF) is a chronic, progressive lung disease characterized by epithelial cell damage, proliferation and activation of fibroblasts and extracellular matrix accumulation which portends a poor prognosis with an estimated mean survival of 2–5 years from time of diagnosis.1 While the etiology of disease progression and exacerbation is unknown, there is growing evidence that double-stranded DNA (dsDNA) may play an important role in IPF pathogenesis.2 Also, we have recently reported that absent in melanoma 2 (AIM2) inflammasome, a multiprotein complex that recognises dsDNA, plays a critical role in IPF pathogenesis and that AIM2 is regulated by glucose metabolism.3 To advance the knowledge of the role of dsDNA in IPF pathogenesis, we examined plasma levels of circulating cell-free dsDNA (ccf-dsDNA) in patients with IPF and identified metabolites and metabolic pathways associated with ccf-dsDNA levels.
Results
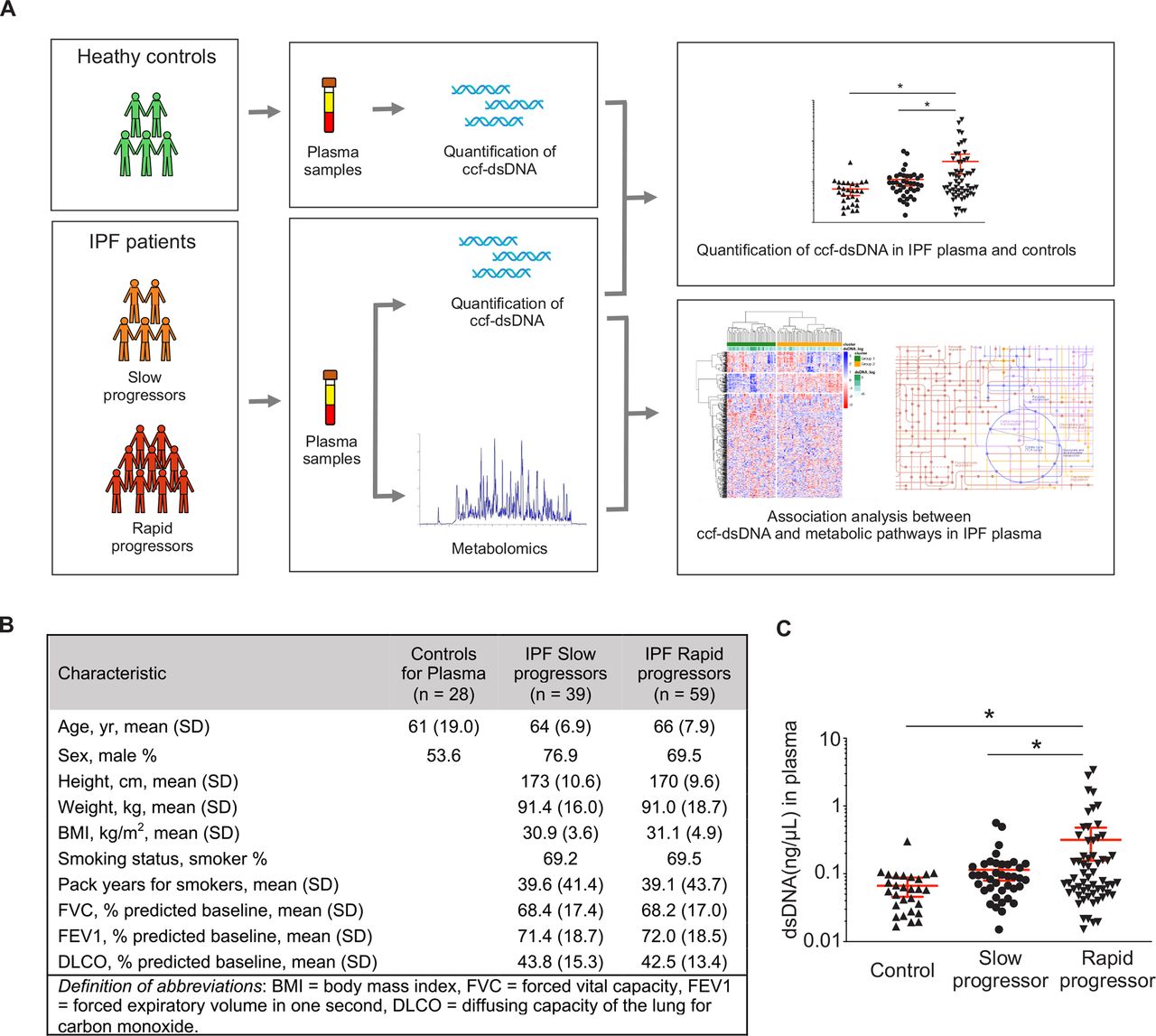
Subjects in this study were patients with IPF from the Correlating Outcomes with Biochemical Markers to Estimate Time to Progression in Idiopathic Pulmonary Fibrosis (COMET-IPF) trial, a multicenter prospective observational study correlating biomarkers with IPF disease progression.4 Rapid progressors were defined as patients who met any part of a composite outcome (death, acute exacerbation of IPF, relative decline in forced vital capacity of at least 10% or carbon monoxide transfer factor of 15%) during the 80-week follow-up. Healthy subjects served as controls (figure 1A). Demographics and clinical characteristics of study subjects are summarised in figure 1B and online supplemental table 1.

Study design and the circulating cell-free double-stranded DNA (ccf-dsDNA) levels in plasma from patients with IPF. (A) Schematic diagram representing overall study design. (B) Demographics and baseline characteristics of patients with IPF and controls. (C) Quantification of ccf-dsDNA concentration of plasma from patients with IPF and controls. Data are mean±SEM. *p< 0.05 using analysis of variance (ANOVA). IPF, idiopathic pulmonary fibrosis.
To investigate the potential role of ccf-dsDNA in IPF, ccf-dsDNA was isolated from plasma of 98 patients with IPF (39 slow progressors, 59 rapid progressors) and 28 healthy controls (figure 1B). Plasma concentrations of ccf-dsDNA were higher in rapid progressor patients relative to those in slow progressors and healthy controls (figure 1C). ccf-dsDNA levels were not significantly different between the slow progressor and control groups.
Untargeted metabolomics assays run on 799 metabolites revealed that 73 metabolites were positively correlated with ccf-dsDNA concentration, while 59 were negatively correlated (figure 2A). Clear clustering of metabolites by ccf-dsDNA levels was noted. From this analysis, we identified two distinct groups of metabolites (group 1 and group 2). Interestingly, the levels of ccf-dsDNA of group 1 were significantly higher than group 2. Differentially changed metabolites were annotated mapped to KEGG pathways. The 15 pathways (10 amino acid pathways, 2 energy pathways, 2 nucleic acid pathways, 1 lipid pathway) with the highest number of correlating metabolites are highlighted in figure 2B. The complete list of significantly associated pathways is shown in online supplemental table 2 and online supplemental figure 1.
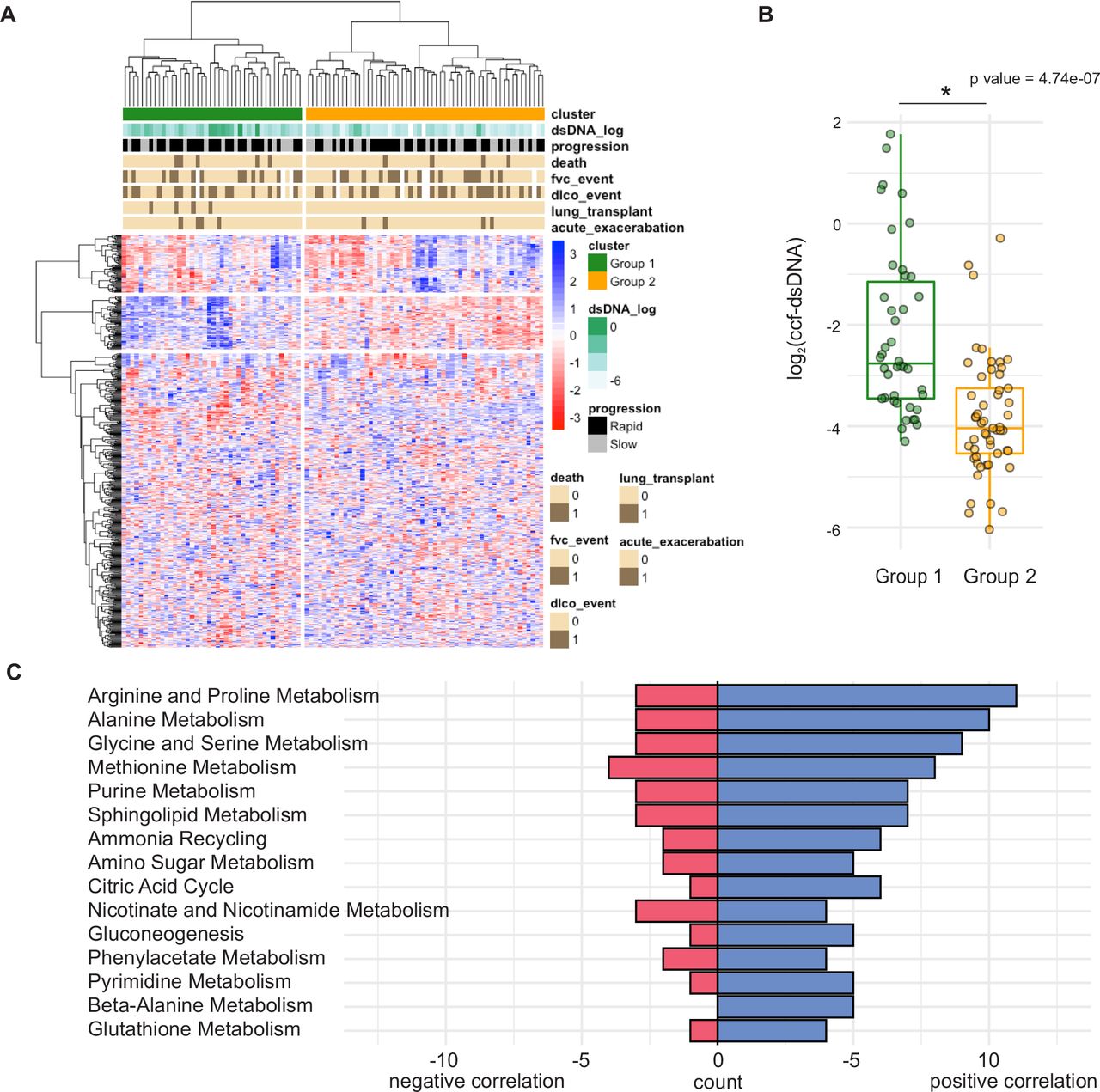
(A) Hierarchical clustering of Kendall’s Tau coefficients calculated for correlations between abundances of dsDNA concentration (rows) and metabolites (columns) in the pairwise fashion, using data from n=98 IPF plasma samples. (B) ccf-dsDNA concentration in group 1 and group 2 defined by the hierarchical clustering. *p<0.05 by Student’s t-test. (C) Metabolome KEGG enrichment analysis of plasma from patients with IPF. Count=number of hits per pathway, p value<0.05. ccf-dsDNA, cell-free double-stranded DNA; IPF, idiopathic pulmonary fibrosis; KEGG, Kyoto Encyclopedia of Genes and Genomes.
There was notable correlation between amino acid metabolism and ccf-dsDNA, especially in the alanine/aspartate/glutamate, arginine, glutathione and taurine/hypotaurine metabolic pathways. Additionally, several lipid metabolic pathways were associated with ccf-dsDNA levels. Key lipid pathways included sphingolipid synthetic and fatty acid metabolic pathway. ccf-dsDNA was also associated with energy metabolism pathways such as glycolysis and tricarboxylic acid (TCA) cycle.
Amino acid metabolism
In addition, specific metabolites pertinent to amino acid pathways were found to correlate with ccf-dsDNA levels (figure 3A). Significant associations existed between ccf-dsDNA and metabolites of the arginine/proline pathway. Arginine is one of the essential amino acids in animal cells, serving as a precursor for the synthesis not only of proteins but also of nitric oxide, urea, polyamines, proline, glutamate and creatine.5 In our study, arginine and creatinine were negatively correlated with ccf-dsDNA, whereas glutamate, spermidine and ornithine were positively correlated. Of note, ornithine converts to proline and hydroxyproline for collagen synthesis during fibrogenesis, while spermidine has been shown to serve as an antioxidant in murine fibrosis models.6

{kind=link}
{kind=link}
{kind=link}
The linear regression graph showing significant metabolites in selected pathways using univariate linear regression. Amino acid metabolism pathways (A), energy metabolism pathways (B) and sphingolipid metabolism pathway (C) are associated with ccf-dsDNA levels. ccf-dsDNA, cell-free double-stranded DNA.
We also found associations between ccf-dsDNA and metabolites of the glutathione pathway. L-cysteinyl-glycine, L-glutamate, 5-oxoproline, spermidine and L-ornithine were positively correlated with ccf-dsDNA, while L-gamma-glutamyl-L amino acid was negatively correlated with ccf-dsDNA (figure 3A). Glutathione, the most abundant antioxidant, regulates gene expression, DNA and protein synthesis, cell proliferation and apoptosis, signal transduction, cytokine production and protein glutathionylation. A prior study showed that cysteine, glycine and glutamate, components of glutathione synthesis, are increased in IPF lung compared with controls,7 consistent with our current observations in plasma.
Like glutathione, taurine is an antioxidant that has diverse cytoprotective function including regulation of antioxidation, energy metabolism, endoplasmic reticulum (ER) stress, neuromodulation, quality control and calcium homeostasis.8 We found that taurine/hypotaurine metabolism is significantly associated with ccf-dsDNA in IPF plasma. Specifically, hypotaurine, taurine, pyruvate, L-alanine, 2-oxo-glutarate and L-glutamate were positively correlated with ccf-dsDNA levels, whereas 3-sulfino-L-alanine was negatively correlated with ccf-dsDNA (figure 3A). This is an important finding as taurine/hypotaurine metabolism has not previously been identified as being altered in lung fibrosis in either animal or human metabolomics studies.
Energy metabolism ccf-dsDNA levels were also found to be associate with energy metabolism pathways (figure 3B), specifically glycolysis and TCA cycle pathways. Glycolysis is the first step in the breakdown of glucose to extract energy for cellular metabolism. In our study, glucose levels negatively correlated with ccf-dsDNA level, while lactate, 3-phosphoglycerate and pyruvate, later-stage glycolysis metabolites, were positively correlated with ccf-dsDNA. The glycolytic byproducts measured in plasma in our study may represent increase in glycolytic activity in IPF lungs, which has previously shown to be a key mediator of lung fibrosis.9 ccf-dsDNA levels were also associated with the TCA cycle pathway which, along with glycolysis, is the key pathway for cellular energy production. We found significant positive correlations between ccf-dsDNA and succinate, fumarate, malate, 2-oxo-glutamate and pyruvate. Conversely, citrate and aconitate were negatively correlated with ccf-dsDNA. These findings in plasma complement data from prior studies showing that TCA cycle metabolites are increased in IPF lung tissue.10
Lipid metabolism
Finally, our study identified significant changes in metabolites involved in sphingolipid metabolism, a bioactive lipid pathway that plays pivotal roles in various cellular processes including inflammation, apoptosis and fibrosis. In our patients with IPF, sphinganine, phosphoethanolamine, sphingosine and sphinganine-1-phosphate were positively correlated with ccf-dsDNA levels (figure 3C). This is an interesting finding in light of a previous study showing a reduction of sphingolipid metabolism in IPF lung.10 The opposing phenomenon observed between plasma and lung tissue may possibly be explained by a surplus of plasma lipid metabolites arising from an underutilisation of these metabolites in lung tissue.
Conclusion
We found that ccf-dsDNA levels are elevated in patients with IPF who are rapid progressors compared with healthy controls and slow progressors and are furthermore associated with alterations in multiple central metabolic pathways. This serves as an interesting complement to prior observations that these same metabolic pathways are significantly changed in IPF lung tissue. The source of ccf-dsDNA in patients with IPF is unknown. Most existing data are from patients with cancer whose ccf-DNA appears to come from different sources, including the tumour cells that circulate in peripheral blood in addition to normal cell types such as haematopoietic and stromal cells.11 In patients with IPF, a recent study showed that mitochondria may be a major source of ccf-dsDNA.12
The association between ccDNA and various metabolites is central to our study. Although we did not perform mechanistic studies in our paper, there is evidence to suggest that metabolic reprogramming regulates dsDNA-dependent-immune responses via cytosolic DNA sensors such as AIM2 inflammasome and cGAS (GMP–AMP synthase)–STING (stimulator of interferon genes). As an example, glucose metabolism, sphingolipid metabolism and fatty acid metabolism all contribute to dsDNA-dependent AIM2 inflammasome activation.13 Evidence linking cGAS–STING signalling and metabolic dysregulation is scarce, which suggests that further investigation is warranted.
In sum, our results suggest ccf-dsDNA both as a potential biomarker for IPF disease progression and severity and also as a mediator of IPF disease progression by modulation of specific metabolic pathways. While no causal relationship can be determined from this observational study, further research may better elucidate the role of ccf-dsDNA and metabolic derangements both as possible biomarkers and as targets for therapeutics in IPF.
Ethics statements
Patient consent for publication
Ethics approval
Subjects included in this analysis were a subset of patients who had participated in a prospective observational study correlating biomarkers with disease progression (clinicaltrials.gov, clinical trials ID no. NCT01071707) (Correlating Outcomes with biochemical Markers to Estimate Time progression in Idiopathic Pulmonary Fibrosis–COMET). Written informed consent was obtained from the individual(s) for that was included in this article.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
WW and MB are joint first authors.
Twitter @mu2bu
Correction notice This article has been corrected since it was published Online First. Competing interests statement has been updated.
Contributors Conception and design: WW, HS-D and SJC; analysis and interpretation: WW, MB, JK, FJM, AMKC, HS-D and SJC and drafting the manuscript for important intellectual content: WW, MB, JK, FJM, BM, AMKC, HS-D and SJC.
Funding HWS-D was supported by grants R01AG052530 and R01AG056699 and SJC was supported by grant K08HL138285.
Competing interests FJM reports personal fees for advisory board work, non-financial support for travel and other from AstraZeneca, other (steering committee work and publication) from Abvie, Boehringer Ingelheim, BristolMyersSquibb, Bridge Biotherapeutics, CsL Behring, DevPro, Genentech, IQVIA, Sanofi, Shionogi, twoXAR, and Veracyte. FJM has served on Steering Committees for ILD studies supported by Afferent/Merck, Bayer, Biogen, NHLBI, Nitto, Novartis, Patara/Respivant, ProMedior/Roche, and Veracyte. FJM has participated in DSMB for studies supported by Biogen and Boehringer Ingelheim. FJM has been supported for study related travel by Boehringer Ingelheim, CsL Behring and Patara/Respivant. FJM has presented nonpromotional presentations for the Academy for Continuing Healthcare Learning, the American College of Chest Physicians, Boehringer Ingelheim, Brooklyn Methodist Hospital, National Association of Managed Care Physicians, Paradigm Communications, PeerView, United Therapeutics, and Vindico. AMKC is a cofounder and equity stockholder for Proterris, which develops therapeutic uses for carbon monoxide. AMKC has a use patent on CO and in COPD. All other authors have no conflicts of interest.
Provenance and peer review Not commissioned; externally peer reviewed.