Article Text

Abstract
To explore the role of extracellular vesicles (EVs) in chronic lung diseases.
EVs are emerging as mediators of intercellular communication and possible diagnostic markers of disease. EVs harbour cargo molecules including RNA, lipids and proteins that they transfer to recipient cells. EVs are intercellular communicators within the lung microenvironment. Due to their disease-specific cargoes, EVs have the promise to be all-in-one complex multimodal biomarkers. EVs also have potential as drug carriers in chronic lung disease.
Descriptive discussion of key studies of EVs as contributors to disease pathology, as biomarkers and as potential therapies with a focus on chronic obstructive pulmonary disorder (COPD), cystic fibrosis (CF), asthma, idiopathic pulmonary fibrosis and lung cancer.
We provide a broad overview of the roles of EV in chronic respiratory disease. Recent advances in profiling EVs have shown their potential as biomarker candidates. Further studies have provided insight into their disease pathology, particularly in inflammatory processes across a spectrum of lung diseases. EVs are on the horizon as new modes of drug delivery and as therapies themselves in cell-based therapeutics.
EVs are relatively untapped sources of information in the clinic that can help further detail the full translational nature of chronic lung disorders.
- lung cancer
- asthma
- idiopathic pulmonary fibrosis
- cystic fibrosis
- COPD exacerbations
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
What are EVs?
Extracellular vesicles (EVs) are lipid-membrane bound vesicles released from almost all cell types and an increasing abundance of literature has demonstrated the numerous roles EVs play in intercellular communication and disease pathogenesis.1 2 The cargo of EVs largely consists of miRNA, proteins and lipid molecules which they transfer to recipient cells.3 Chargaff and West first reported on the potential existence of EVs in 1946 describing the presence of ‘lipoproteins of very high particle weight’ in serum.4 Further studies described lipid‐rich particles originating from the granules of platelets that accelerate coagulation.5 6 In the 1980s, Harding et al 7 reported on multivesicular endosomes which fused with the plasma membrane of reticulocytes and released cargo into the extracellular space. Numerous studies were then undertaken to better understand the vesicle trafficking process being displayed in numerous cell types.8–10 Small vesicles emanating from the intracellular secretion pathway or multivesicular bodies became known as exosomes with larger particles shed from the plasma membrane described as microparticles.
The International Society for Extracellular Vesicles (ISEV) currently provides guidelines on categorising EVs. The minimal information for studies of extracellular vesicles (MISEV) paper published by ISEV summarises the difficulty in including biogenesis and origin as classifications for EV subtypes as consensus has not yet been reached on appropriate markers.11 Doyle and Wang make the argument that markers associated with vesicle formation should be used with nomenclature such as exosome and microvesicle.12 According to MISEV, there are three appropriate ways to subcategorise EVs (1) size alone, for example, small EVs, medium/large EVs (2) by markers, for example, CD81±EVs or (3) by known cellular origin, for example, from breast cancer cell culture.11
Table 1 below summarises the different EV subcategories encompassing their definition by size and biogenesis with indicative markers.An important caveat noted (*) is that there is cross sectionality in markers for vesicles which have similar exit strategies.
Guide on extracellular vesicle (EV) subcategories by size, formation and formation-associated markers2 11 12 147
Cellular origin of EVs is particularly important in the clinical setting. MISEV provides guidelines on marker proteins which are cell/tissue-specific, table 2 summarises these. The inclusion of cellular origin has expanded the EV field to include other large vesicles such as oncosomes. Oncosomes carry distinct protein cargo and are considered to be a separate functional class of tumour-derived vesicles.13
Markers used for cellular origin of extracellular vesicles11
In this review, we focus on EVs as both biological mediators of intercellular communication and biomarkers of disease, particularly in response to respiratory stressors in COPD, CF, asthma, idiopathic pulmonary fibrosis (IPF) and lung cancer.14
Methods of EV isolation and analysis
Isolation
EV isolation methods commonly exploit the physical or chemical properties of EVs. A challenge within EV research can be to obtain well-defined EV fractions of high quantity and purity to facilitate analytical characterisation. Table 3 summarises the most commonly used approaches and provides potential advantages/disadvantages of each.
Summary of commonly used extracellular vesicle (EV) isolation methods
One of the biggest difficulties of EV research is the relatively long sample isolation times and suffer from lower yield and purity.15 Therefore, combined isolation/analysis systems are gaining traction and are of particular benefit for use in the clinical setting.16 17 One such system is the NanoView which uses a ‘lab-on-a-chip’ set-up, with EV antibodies immobilised on silicon chips. The benefit of a system such as this is that; small volumes (35 µL) of unprocessed biological samples are used, without prior isolation.18 However, this system is limited, as prior knowledge of the target protein is required and limits exploratory characterisation.
Analysis
Characterisation of individual EVs can be achieved using different instruments. Nanoparticle tracking analysis (NTA) based on the illumination of nanoparticles in suspension with a laser beam is one of the most widely used methods of establishing the size and quantity of EVs in a sample.19 Resistive pulse sensing (RPS) is an increasingly common technique for determining the size and distribution of EVs.20 21 RPS allows for accurate size determination of single particles by measuring the change in resistivity as a particle of a certain size passes through a pore. However, the method is limited by the pore size range.21 Flow cytometry is another frequently used technique for single-particle analysis, particularly in clinical settings.22 Benefits of this technique over others, such as NTA, are that it incorporates a number of detection modes and arguably provides a more accurate analysis due to user-defined thresholds.23
Multifaceted ‘omics’ approaches which often include mass spectrometry (MS) (proteome, lipidome) and RNA sequencing analysis (transcriptome) have successfully enabled profiling of the EV in chronic lung diseases.24 25 RNA-Seq and microarrays are two of the most commonly used methods to sequence the transcripome.25 Specific studies analysing RNA profiles within EVs are highlighted in the disease sections. MS is a developing area used to characterise the protein and lipid content of EVs. Accurate protein identification and quantitation have dramatically advanced due to high-performance MS technologies.26 Bottom-up proteomics approaches which involve the proteolytic digestion of proteins before MS analysis have enabled the identification of thousands of proteins in EVs.27 A bottom-up proteomic strategy is highlighted in figure 1. Alternative strategies such as top-down proteomics, which sequences intact proteins without the need for proteolytic digestion and targeted proteomic approaches such as multiple reaction monitoring also hold promise in EV research.28 29
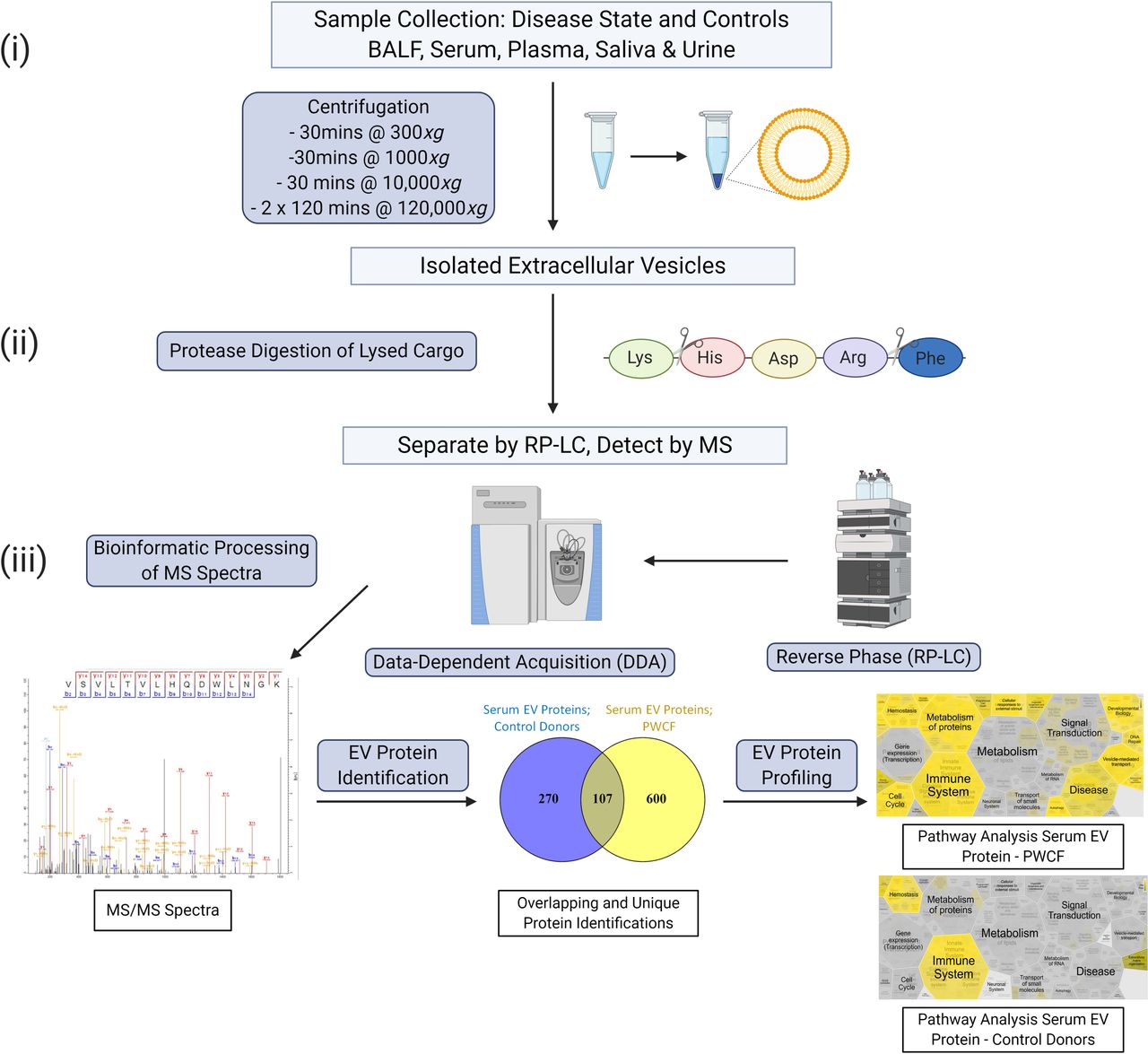
{kind=link}
Overview of EV isolation, digestion and reverse-phase liquid chromatography (RP-LC) mass spectrometry (MS) strategy. (i) Biofluids are collected and processed by standard and ultracentrifugation. (ii) The isolated EVs are lysed and digested with trypsin and the resulting peptides are then separated by RP-LC and analysed by MS. (iii) The MS scans generated are searched against databases using algorithms to produce qualitative and quantitative data. Bioinformatic tools enable identification of unique EV disease profiles. EV, extracellular vesicle.
Proteomic profiling of EVs has successfully identified markers of acute exacerbation in COPD and unique protein fingerprints at different ages in persons with CF (PWCF), demonstrating the potential of EVs as markers for disease severity.30 31 MS analysis of EVs has successfully identified differential proteomes in patients with lung adenocarcinoma (ADC).32 Additionally, EVs isolated from nasal lavage fluid of asthmatic donors exhibited unique proteomes compared with controls.33
Lipidomics involves the specific characterisation of the lipid profile of a sample. As EVs are encapsulated in a lipid bilayer, understanding their lipid content and their potential as biomarkers is of increasing interest in lung disease.34 35 Both targeted or global MS analysis of lipids can be performed using high-resolution mass analysers enabling the identification of lipid species by accurate mass.35 A study on the role of CF-EV derived sphingolipids in inflammation showing that the lipid profile of CF-EVs was significantly different compared with controls.36 Another study characterising the lipidome of EVs isolated from BALF from asthmatics and healthy controls identifying unique lipid EV profiles in patients with asthma.37
EVs as mediators of intercellular communication in the lung
It has been well established that EVs play a significant role in cellular communication.38 Cellular origin and microenvironment directly influence EVs’ effect on the cells they communicate with.39 EVs can bind to recipient cells by interactions with tetraspanin proteins, integrins (ITGs), immunoglobulins and proteoglycans.40 Once bound they can deliver their message by entering the cell or by ligand–receptor activation. Mechanistically it is thought that EVs can directly interact with receptors on the plasma membrane of cells which may help target them to certain cells. Hoshino et al 41 demonstrated tumour-derived exosomes could be directed to organ-specific cells by targeting the ITG, α6β.4, and concluded that exosomal ITGα6β4 activates the Src-S100A4 axis in lung fibroblasts during premetastatic niche formation. Our research demonstrated exosomes derived from bronchial epithelial cells caused activation of the receptor for advance glycation end-products (RAGE) receptor and mitogen activated protein kinase (MAPK) pathways in neutrophils.31
After entering the cell, the EV is endocytosed and releases its cargo by three hypothesised methods: by fusing with the plasma membrane, fusion with the endoplasmic reticulum, fusion with the endosome membrane or rupture of the endosome membrane.42 Cellular studies have traced the cellular uptake of labelled EVs with high-resolution microscopy to study the release of EV cargo. Joshi et al 42 demonstrated that after endocytosis of green flourescent protein (GFP) labelled EVs into HEK293 cells, internalised EVs fused with the membrane of endosomes/lysosomes resulting in EV cargo exposure to the cell cytosol. A study by Tian et al 43 investigated the fate of EVs following endocytosis. By simultaneously labelling the EV membrane and cargo the authors demonstrated that inner EV localised proteins were trapped within the lysosome.43 The authors hypothesised that as the EVs originated from the same cell type as the recipient cell the abundance of existing proteins within the cell resulted in express expulsion.43 Additionally, the proinflammatory environment within the lung can subject EVs to degradation by proteases and lipases.44 Degradation of the EV can release free ligands which can bind to cell surfaces in the localised area, inducing a signal.44
The lung is a unique organ providing defence and against viral and bacterial pathogens. Defence against respiratory pathogens requires communication between structural and immune cells. The lung airway epithelium acts as both a physical barrier and stimulates innate and adaptive immune mechanisms important for maintaining lung homeostasis.45 EVs play a role in maintaining the lungs unique microenvironment’s homeostasis through intercellular communication. Mast cell-derived EVs have been shown to exhibit TGF-β on their surface and to induce epithelial–mesenchymal transition in airway epithelial cells within the lung.46 The presence of cytokine suppressor molecules in CD8α+CD11c+ lung EVs and alveolar macrophage-derived EVs contributed to regulating immune balance in the respiratory tract in asthma and COPD models.44 47 48 A role for EVs in proinflammatory signalling is described further in this review and has been the topic of other reviews and studies.37 49 50
The source of EVs within the lung has a significant contribution to the message they carry. Endogenous sources of EVs in the respiratory tract include epithelial cells and haematopoietic cells such as eosinophils, monocytes, macrophages and neutrophils. However, this is not an exhaustive list; EVs can be shed from endothelial cells. Zhao et al 51 demonstrated damaged pulmonary artery endothelial cells release an abundance of EVs.39 Bronchial epithelial cells are a primary source of reactive proinflammatory EVs in the lungs as they are most frequently exposed to insult and injury.49 EVs can also be released from bacterial sources. Exogenous bacterial EVs, expressed by pathogens in the lung, can create optimum growing conditions, encourage pathogen proliferation and prevent host immune responses52
EVs as biomarkers in lung disease
EVs can be found and isolated from most biological fluids in clinical disease cohorts. In respiratory research, bronchial lavage fluid (BALF) and blood are some of the more frequently studied sources of EVs.53 In chronic lung inflammatory disease, stress-induced EVs are correlated to pathogenesis.54 Examples of such studies are outlined within the individual disease sections. Chronic pulmonary disorders are among the most frequent causes of death, claiming the lives of at least four million people annually around the world.55 56 Shared characteristics of chronic lung disease are progressive reduced lung function and inflammation. Clinical symptoms include cough, chest tightness, shortness of breath and mucus production. Acute exacerbations of chronic lung disease57 are often triggered by infection and contribute to disease progression and disease-associated mortality.58
There is a knowledge gap in the management of chronic lung diseases, including early identification, diagnostic biomarkers, and treatments.58 59 Biomarkers can be used as indicators of the presence and severity of a disease. They can help us understand the cause and progression of a disease, achieve early prognosis or predict treatment outcome. The search for useful biomarkers in the clinic can be difficult, with many biomarkers failing at verification and validation stages before they enter clinical practice.59 Many studies have been limited to examining the relationship between individual biomarkers and cross-sectional outcomes.60 However, recent investigations demonstrated that using multiple biomarkers can be more predictive of cross-sectional and longitudinal COPD outcomes.61
Due to EV cargoes’ disease-specific nature, these intercellular communicators can be an all-in-one complex multimodal biomarker.62 Changes in the number and content of EVs have been reported during exacerbation in COPD, asthma and CF.31 63 64
EVs as therapeutic tools in lung disease
EVs have the potential to be novel drug delivery vehicles. Liposomal drug delivery systems, such as doxorubicin, a Food and Drug Adminstration (FDA)-approved chemotherapeutic, are highly effective in reducing the toxic effects of the drugs they carry.65 However, liposome-based drug delivery has drawbacks, often the liposome is cleared from the system before reaching the target cell and can elicit adverse immune responses.66 Therefore, EVs have come to the forefront as they are naturally occurring molecular carriers. Biologically derived EVs are less likely to elicit an allergic immune response or be prematurely cleared and are considered to have a better safety profile than synthetic carriers.67 Cell and gene-based therapies are of particular interest in inherited disorders such as CF 68 A difficulty in effective RNA therapy is the relative instability of the nucleotides in circulation.69 EVs are often chosen as delivery systems for RNA due to the aforementioned stability and role in communication.69 However, EV mediated drug delivery can come with inherent difficulties due to the heterogeneity of biologic production.70 Mesenchymal stem cell (MSC) derived EVs have shown potential in the area of regenerative medicine.71 Studies using MSC-EVs in chronic lung disease are discussed further in this review. In the vaccine arena, research has been conducted using bacterial EVs in vaccine development. One such study by Olaya-Abril et al used EVs from Streptococcus pneumoniae to inoculate mice and conferred protection from pneumococcal infection.72
EVs in respiratory disease
COPD
COPD is a group of inflammatory lung diseases characterised by emphysema, bronchitis and obstructive airway.73 It is the third most frequent cause of death globally55 and EVs have been demonstrated to be functionally significant in COPD.74 They have shown biomarker potential, capable of distinguishing between stress-induced states and periods of exacerbation.75 They have also been associated with the inflammatory response in COPD airways.76 Mechanistic in vitro studies of EV roles in fibroblast accumulation in COPD have provided valuable insights into the pathophysiology of this disease. Fujita et al 77 investigated the contribution of EVs in airway remodelling in COPD. They found that EVs carrying miR-210 could suppress autophagy responses and induce myofibroblast differentiation suggesting a potential therapeutic target.77 Similarly, miR-21, derived from bronchial epithelial EVs, was shown to mediate myofibroblast differentiation and contribute to airway remodelling.78 Genschmer et al 79 studied the enzymatic cargo of polymorphonuclear neutrophil derived EVs and found they transport highly potent neutrophil elastase (NE). NE can degrade the extracellular matrix (ECM), causing significant damage and the hallmarks of COPD.79
There have been several studies on the proinflammatory role of EVs in COPD. A study by Kim et al 80 demonstrated that EVs from Escherichia coli could induce emphysema via IL-17A-mediated neutrophilic inflammation. Tan et al grouped patients according to the global initiative for COPD (GOLD) criteria for COPD exacerbations, they observed that elevated levels of circulating EVs in patients with COPD are associated with markers of systemic inflammation including C-reactive protein (CRP), sTNFR1 and IL-6.75 However, in a prospective study of patients with COPD, Takahashi et al 81 found that there was a negative correlation between increased circulating EVs and forced expiry volume (FEV)1 levels. An earlier study by Takahashi et al 63 showed increased levels of circulating endothelial microparticles in patients with COPD correlated with a decrease in FEV1/FVC ratio percentage.
There have been numerous studies into using EVs as biomarkers for COPD progression. Several studies have examined differential miRNA and protein expression after smoking.82 One such study by Sundar et al 83 performed differential expression analysis on miRNAs derived from the EVs of smokers, patients with COPD and non-smokers to investigate the potential unique and common nucleotides between the groups. In a comparison of non-smokers versus COPD groups, they found miR199a-5p was upregulated and a previous study by Mizuno et al 84 had shown that there was a negative correlation between miR199a-5p levels and lung function (FEV1)). Héliot et al 85 conducted a study on the effects tobacco smoke had on EVs’ miRNA content. This study found that while EV diameter and concentration showed no variation between smokers and non-smokers, there was a significant downregulation of the let-7 family of miRNA.85 Though this study used clinical samples there was no comparison drawn between the clinical measurements of the effects of cigarette smoking and let-7 miRNA.
Carpi et al 86 investigated the differential expression of skeletal muscle-specific miRNA cargo in EVs derived from serum of patients with COPD. Patients were grouped into four groups (A, B, C and D) relevant to disease severity by GOLD criteria. This study found that key miRNAs were upregulated in GOLD group D compared with other groups with clinical characteristics of dyspnoea, COPD assessment test, FEV1 and FEV1 predicted percentage all being significantly different in this group.86
Cystic fibrosis
CF is a life shortening disease caused by dysfunction in the cystic fibrosis transmembrane receptor resulting in impaired mucociliary clearance, inflammation and recurrent bacterial infection.87 A role for EVs as modulators of inflammatory cell activation in CF is emerging. An early study by Porro et al 88 demonstrated that microparticles obtained from CF sputum are proinflammatory when injected into the murine lung and illicit a strong neutrophil response. Analysis of EVs isolated from BALF of PWCF and primarily ciliary dyskinesia demonstrated that disease state EVs contained higher levels of proteins involved in leucocyte chemotaxis than controls.89 We demonstrated that CF epithelial-derived EVs were enriched with inflammatory markers such as S100 A12 that could regulate neutrophil migration.31
Our group and others have explored the biomarker potential of EVs in CF. Zulueta et al showed that the lipid profile of CF-EVs was significantly different than controls.36 Enhanced ceramide production leads to the release of EVs that export proinflammatory ceramide to the recipient cells, maintaining the unresolved inflammatory status of CF.36 Our group performed a study on EVs from BALF of four different age groups of PWCF and CF cell lines.31 We identified unique protein fingerprints and pathways between CF and control groups. Significant changes in EV number and protein content in CF adults undergoing exacerbation relative to controls were also observed.31 We also investigated the correlation between clinical markers such as IL-8 and NE found in BALF and protein expression in EVs from PWCF. A significant correlation between IL-8 levels and 29 EV proteins was found.
The therapeutic potential of lung MSC EVs was investigated as a potential anti-inflammatory treatment in CF.90 Using an in vitro model, Zulueta et al found that MSC EVs downregulated the expression of proinflammatory cytokines IL-1β and IL-6.90
Asthma
Asthma is a chronic inflammatory disease of the airways with a complex pathophysiology. There are multiple endotypes of asthma making the development of diagnostic and therapeutic tools a challenge.91 EVs play a role in the pathology and progression of asthma by eliciting proinflammatory responses.92 EVs isolated from nasal lavage fluid of asthmatic donors exhibited chemotaxis of monocytes, natural killer cells and neutrophils.33 Profiling of EVs from BALF of patients with asthma identified proteins involved in toll-like receptor (TLR) signalling, orchestrating the inflammatory response.89 A study by Kim et al showed EVs from Staphylococcus aureus stimulated neutrophilic inflammation via TLR signalling.93 The functional effects EVs have in asthma has been recently investigated in several studies. Hough et al characterised the lipidome of EVs isolated from BALF of asthmatics and healthy controls, identifying unique lipid EV profiles in patients with asthma concluding that the altered lipidome could drive chronic inflammation.37 Hough et al also found positive correlations between plasma eosinophilia and EV particle count and IgE titre and particle count.37
EVs from nasal polyps were found to have increased proteins associated with cell proliferation and thus remodelling of mucosal membranes.94 Treatment of human bronchial epithelial (HBE) cells with proinflammatory cytokines, specific to two subsets of asthma, T2 and T17 (T2 treated IL-4 and IL-13, T17 treated IL-17A and TNFα) increased EV release.95 EVs isolated from a murine model of asthma were found to have increased levels of immune cell-derived miRNA, such as miR-223 and miR-142a.96 Interestingly, compared with controls, the dominant cell type origin of EVs changed from largely epithelial cells to haematopoietic cells on allergen exposure.96
The use of EV miRNA cargo for potential biomarkers for pulmonary diseases has been discussed here concerning other conditions. Levänen et al 97 investigated the miRNA content of BALF EVs from patients with asthma. They found that there was a significant difference between asthmatic miRNA and that of controls and a significant correlation to EV miRNA and decreased FEV1. Similarly to the Héliot study of COPD, Levänen found that the let-7 family of nucleotides were among the most significantly downregulated in asthmatics.97 Mendes et al 98 used breath condensate to investigate a potential miRNA based platform for asthmatic endotyping. Let-7 and other miRNAs were shown in exhalate, in sufficient quantity and with a significant difference, to enable potential phenotyping of asthmatic children.98 Duarte et al 64 conducted one of the first investigations of EVs as biomarkers for asthma. They found that EVs levels were significantly increased in asthmatics and found strong but not significant indications of inflammatory markers.64 In the study by Ax et al, 95 several genes and proteins were differentially expressed in the two models compared with controls. In a recent communication, Bahmer et al 99 indicated that miR-122-5p had potential to differentiate endotypes of asthma. They isolated miRNA from the plasma EVs of people with mild‐to‐moderate or severe eosinophilic asthma and healthy controls and found that counts of miR-122-5p positively correlated with blood eosinophils, neutrophils and lymphocytes.99 Investigating the same miRNA in sputum EVs from patients with asthma and healthy controls showed promising indications of an increase in asthamtics.99
Idiopathic pulmonary fibrosis
IPF is a progressive lung disease that occurs due to increased fibrosis of lung tissue in response to chronic injury of the epithelium.100 Increased fibroblast activity and ECM accumulation in the lung are characteristics of the disease.101
Cellular senescence is a feature of the epithelial response to IPF.102 EVs released from senescent cells have been linked to reduced reparative stem cell activity.103 As intercellular communicators, EVs recruit fibroblasts to the ECM of the IPF lung. Chanda et al 104 showed that fibroblast EVs had high ECM protein levels, including fibronectin. Lacy et al 105 demonstrated that normal lung EVs are enriched in antifibrotic prostaglandin and inhibit myofibroblast differentiation. They demonstrated that reduction of antifibrotic prostaglandin could trigger an exacerbation of IPF.105 Parimon et al conducted a study of IPF phenotype mouse lung EV’s profibrotic qualities.100 Using RNA-seq technology, they found that TGF-β and Wnt signalling pathways were upregulated in mouse lungs inoculated with IPF EVs.100 Martin-Medina et al 106 investigated the clinical relevance of EV-mediated WNT5A expression in IPF fibrogenesis. They found increased levels of WNT5A in EVs of people with IPF compared with non-IPF controls and a correlation between EV-associated CD81 and WNT5A.106
Biomarker discovery in IPF is a clinically important field due to the disease’s multiple origins.101 Makiguchi et al tracked the expression of serum EV miR-21-5 across 41 patients with IPF, over 30 months.107 They observed that baseline levels of miR-21-5 could predict mortality in IPF over the 30-month period and could be used as an effective indicator of disease exacerbation.107 They found that there was a significant correlation between the levels of EV miR-21-5p and the rate of decline in predicted vital capacity of the lung over 6 months.108 Carleo et al 109 investigated potential biomarkers for familial IPF. They showed protein associated with clathrin-coated vesicles were part of a group of proteins upregulated in familial IPF.109 Njock et al have recently published two clinical studies investigating the role of EV-derived miRNA in the progression of IPF.110 111 Analysing the miRNA content of sputum-derived exosomes, the authors found seven upregulated miRNAs, and found a negative correlation between miR-142-3p and diffusing capacity of the lungs for carbon monoxide/alveolar volume.111 A further study by the group showed strong indications that upregulated miR-142-3p, derived from sputum and plasma exosomes of patients with IPF, were of macrophage origin.110 Guiot et al 110 also found that over expression of miR-142-3p could reduce expression of TGFβ-R1 and ultimately have an antifibrotic effect.
Using EV associated proteins for targeted therapies is relatively understudied. Marchetti et al 108 developed antiplasmalemma vesicle-associated protein (PV1) antibodies to enhance therapeutic delivery to the lungs. The PV1 antibody was conjugated to an antifibrotic small molecule and showed increased localisation of the small molecule to the lung and kidneys.108 Wan et al 112 investigated bone marrow stem cell (BMSC) derived EVs as a potential treatment for IPF. They found that BMSC EVs with over-expressed miR‐29b‐3p could inhibit fibroblast activity.112 Mansouri et al 113 demonstrated the ability of MSC-EVs to alter the phenotype of bone-marrow monocyte, aiming to elicit the same anti-inflammatory phenotype in lung monocytes as a treatment for IPF. Dinh et al 114 used treated murine models of IPF with lung spheroid cell (LSC)-derived exosomes. This study found that LSC-exos were enriched with miRNAs largely conserved in stem cells and promoted lung repair in the murine IPF model.114
Lung cancer
Lung cancer is the leading cause of cancer death worldwide.115 Although traditionally not considered a chronic disease, advances in biopharmaceuticals and surgical techniques for lung cancer treatment have created cohorts of long-term survivors.116 It is well established that EVs have numerous roles in lung cancer, including carcinogenesis, as biomarkers, as treatments and in treatment resistance.117 In the seminal paper by Fabbri et al, the authors investigated the prometastatic functionality of lung cancer-derived EV mRNA.118 They showed that mRNA could function as a ligand to TLR and induce inflammatory cells to create a metastatic niche.118 More recent studies have shown the significant and wide-ranging impact cancer EV-derived mRNA has on promoting cancer progression in the lung.119–121 Explorative approaches for integrating liquid biopsy into managing common cancer types have been developed, including 32 studies focusing on EVs.122 Here, we concentrate on EV studies that identified biomarkers between control and disease states and targets contributing to lung cancer progression.
Profiling EV proteins from sera in patients with early and advanced non-small cell lung carcinoma (NSCLC) identified fibronectin as a potential marker for clinical use.123 Qiao et al identified several EV targets in metastatic lung cancer, functionally important in driving metastasis, including hepatocyte growth factor.124 Salivary exosomes from patients with lung cancer were isolated, and several candidate diagnostic biomarker proteins such as SPARCL1, ENO1, IQGAP1 and BPIFA, all of which have roles in the survival and proliferation of lung cancer.125 EVs derived from patient with lung ADC serum were analysed and compared with healthy controls.32 Four proteins could distinguish ADC from controls, SRGN, TPM3, THBS1 and HUWE1 are primarily vesicle-associated and support EVs as enrichment sources for biomarkers.32 Cazzoli et al used EV microRNA to differentiate between ADC, granuloma and healthy controls.126 Comparing the small RNA cargo of ADC, NSCLC and control derived serum EVs showed differential expression of sRNA between the groups.127 Interestingly, comparison of sRNA from NSCLC cell lysate to serum EVs showed that miR-451a and miR-122-5p were significantly downregulated in lysate but significantly upregulated in EVs.127 A study of serum exosomes from metastatic and non-metastatic NSCLC showed that lipopolysaccharide-binding protein was a highly sensitive and specific marker for metastasis.128 Exosomes, isolated from pleural exudate of patients with lung cancer, contained protumour survival factors.129 The study found that exosomes from patients with lung cancer contained γ-glutamyl transpeptidase 1, which converts exogenous leucotrienes to a protumorigenic form.129 Choi et al recently investigated the correlation between circulating levels of EVs in pulmonary and peripheral blood and stages of lung cancer.130 They found that EVs were increased 10-fold in the peripheral blood of patients with lung cancer and 19-fold in the pulmonary blood, further they found a significant correlation between advancing cancer stages and EV counts in pulmonary blood.130
Research into effective and safe cancer therapies is a continually growing field, as is research into keeping therapies potent and reducing resistance. Ma et al conducted a study into the role EVs have in chemo-resistance.131 Increased EV miR-425-3p was found in serum of patients with NSCLC during chemotherapeutic treatment.131 In vitro, chemoresistant EVs induced a resistant phenotype in untreated cells, miR-425-3p drove this process.131 There have been numerous studies in EV associated resistance, and they are reviewed in greater depth elsewhere.132–134
As discussed previously, EVs can be drug carriers. Garofalo et al combined an oncolytic adenovirus and a chemotherapeutic within ADC EVs.135 Though the treatment showed accumulation in both the liver and lungs, a subsequent study by the group posited that an intravenous administration was preferential to intraperitoneal and could reduce accumulation.136 Similar to other respiratory diseases, cell-based therapies are also a viable therapeutic route in lung cancer.137 138
The future of EVs and EV profiling in clinical respiratory medicine
There is a role for EVs in both clinical research and diagnosis. As evidenced in this review, EVs can provide new insight into the pathology of respiratory disease. Their role as cellular communicators is key to their contribution to pathogenesis. They coordinate ECM remodelling in COPD and IPF.74 104 In CF, they induce chemotaxis of leukocytes.31 They influence TLR signalling in asthma and cancer, orchestrating inflammatory responses and preparing metastatic sites.89 118
Profiling of EVs in the clinical arena could lead to a refined diagnosis for individual patients, directing the right therapies to the right patients.139 Response to treatments, particularly personalised treatments, often give insight into disease processes. The availability of clinical cohorts now receiving personalised treatments for respiratory diseases presents a unique opportunity to gain a greater understanding of EVs. Novel model systems such as organoids are being employed in the drive towards personalised medicine and offer opportunities in more physiologically relevant in vitro states to understand the functional role of EVs.140 However, challenges remain in the analysis of EVs within clinical studies. To date, studies of EVs in lung diseases have recruited modest numbers of patients. More large scale, prospective, multinational studies are required to precisely define the benefits of EV profiling in specific lung disorders. Where applicable the clinical parameters in EV studies discussed here have been stated above. However, further correlations between EV count or content and the clinical indicators of lung disease need to be performed. Additionally, the field lacks in clinical studies with the many investigations performed ex vivo, in vitro and using in vivo murine models.
A further issue is that pulmonary diseases are physiologically complex and occur on a spectrum of severity, which adds an additional layer of difficulty.62 Identifying robust biomarkers that can distinguish between exacerbated and steady states will be beneficial. Though caution is warranted, as the field of EV research grows rapidly, there is a tendency to focus on target cargo molecules while the broader landscape goes under-reported. Studies cited here have achieved this to some extent, but there is still much to be understood. Advances in omics technologies such as the advent of high-end mass spectrometers and new RNA sequencing technologies will enable deeper profiling of EVs enabling greater discovery of EV biomarkers.141 142 Identification of novel EV markers will also contribute towards better classification of EVs subpopulations addressing the challenge of EV heterogeneity.
Standardisation of EV methods is essential for the field to move forward and positively impact patient care. However, until there is consensus on isolation protocols analysts are required to balance the advantages and disadvantages, as discussed in table 3, of each method. Transparent reporting and centralising knowledge in EV research (EV-TRACK) platform is a collective effort to bring standardisation to EV studies by ‘scoring’ a study based on the techniques used in isolation and characterisation.143 Continued guidelines from organisations, such as ISEV, and evolution of platforms such as EV TRACK, will increase uniformity and improve standardisation.11 143–146 There are inherent difficulties in EV isolation that will require more than standardisation to resolve namely the large amounts of sample required to yield ‘useable’ quantities of EVs and the limitations of acquiring fresh samples in a longitudinal study. Though many groups, including our own, have gained valuable information from EV derived from deep-frozen samples the effects of freeze-thaw on the lipid-membrane are not fully understood.
The complexity and variety of chronic respiratory diseases call for more effective targeted therapies. MSC EVs have shown promise in reversing inflammatory environments of chronic lung disease.90 112 EVs have been shown to safe, effective carriers of drugs due to their specific cell targeting properties and stability. Although EVs have not elicited allergic responses in studies to date further investigations are needed to examine their immunogenicity profiles and potential to generate autoantibodies which could reduce the effectiveness of therapy.67
The future for EV biology is bright and where the advent of new EV technologies will provide greater insights into different disease states in respiratory medicine ranging from early identification and diagnosis, to the prediction of exacerbation and effects of disease-modifying treatments.
Ethics statements
Footnotes
Contributors The article was conceived by JAC, AT, PMN and SCD. AT and JAC were responsible for the initial draft preparation and revision. All authors were involved in reviewing each draft and approving the final manuscript.
Funding This work was supported by The National Children’s Research Centre under Project Grant No C/17/3.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.