Article Text

Abstract
Natural killer (NK) cells are innate lymphoid cells that have been increasingly recognised as important in lung allograft tolerance and immune defence. These cells evolved to recognise alterations in self through a diverse set of germline-encoded activating and inhibitory receptors and display a broad range of effector functions that play important roles in responding to infections, malignancies and allogeneic tissue. Here, we review NK cells, their diverse receptors and the mechanisms through which NK cells are postulated to mediate important lung transplant clinical outcomes. NK cells can promote tolerance, such as through the depletion of donor antigen-presenting cells. Alternatively, these cells can drive rejection through cytotoxic effects on allograft tissue recognised as ‘non-self’ or ‘stressed’, via killer cell immunoglobulin-like receptor (KIR) or NKG2D receptor ligation, respectively. NK cells likely mediate complement-independent antibody-mediated rejection of allografts though CD16A Fc receptor-dependent activation induced by graft-specific antibodies. Finally, NK cells play an important role in response to infections, particularly by mediating cytomegalovirus infection through the CD94/NKG2C receptor. Despite these sometimes-conflicting effects on allograft function, enumeration of NK cells may have an important role in diagnosing allograft dysfunction. While the effects of immunosuppression agents on NK cells may currently be largely unintentional, further understanding of NK cell biology in lung allograft recipients may allow these cells to serve as biomarkers of graft injury and as therapeutic targets.
- innate immunity
- lung transplantation
- lymphocyte biology
Statistics from Altmetric.com
Introduction
Natural killer (NK) cells are innate lymphoid cells increasingly recognised as important in immune responses to solid organ allografts.1 2 NK cells were discovered in the 1970s based on their ability to spontaneously lyse tumours and virus-infected cells in the absence of prior experience.3–5 Cells with NK-like properties are found in species as evolutionarily remote as the golden star tunicate Botryllus schlosseri, for which avoiding fusion with its distant relatives may provide a selective advantage.6 NK cells respond to non-self cells by direct cytotoxicity mediated by perforin, granzymes and tumor necrosis factor (TNF) family effector molecules and by the production of effector cytokines, such as gamma interferon (IFN γ ).7 8 Evidence for their importance includes the observations that humans lacking functional NK cells are subject to certain viral and bacterial infections.9 10 The receptors that NK cells use to identify infected cells can also distinguish between healthy, transformed, malignant and stressed cell populations.11
In contrast with T and B cells, whose specificity is determined by a T or B cell receptor that is diversified through somatic cell genetic recombination, NK cell functions are dependent on integration of signals derived from a range of activating and inhibitory receptors (figure 1).11 These germline-encoded receptors are stochastically expressed in numerous combinations on subsets of NK cells, rendering NK cells capable of responding to a broad range of targets. Human NK cells are identified by expression of CD56 and NKp46 and the absence of lineage-specific markers for T cells (CD3), B cells (CD19) and monocytes (CD14).12 NK cells comprise between 5% and 20% of the total peripheral blood lymphocyte population.12 Approximately 10% of resident lymphocytes in the mouse lung are NK cells.13 During pulmonary inflammation, additional NK cells traffic from peripheral blood or other sites to the lung.14 Within the lung, NK cell activation and proliferation are enhanced by interleukin (IL)-15 secreted by bronchial epithelial cells and suppressed by transforming growth factor (TGF-ß) secreted by alveolar macrophages.15

NK cell receptors and ligands. The NK cell activating and inhibitory receptors that are discussed within this review article are depicted with their associated ligands. HA, haemagglutinin; HLA, human leucocyte antigen; IL, interleukin; IgG, immunoglobulin G; KIR, killer cell immunoglobulin-like receptor; LAIR1, leukocyte-associated immunoglobulin-like receptor 1; MICA/B, MHC class I polypeptide-related sequence A and B; NK, natural killer; ULBP, UL16-binding proteins.
NK cell function and relevance in lung transplantation
NK cells are prevalent and play a critical role in host responses to infection, especially within the lungs. The ability to distinguish self from altered-self or missing-self, paired with potent and diverse effector abilities make NK cells particularly relevant to lung transplantation. A diverse array of receptors lead to multiple mechanisms by which NK cells could potentially modify clinical outcomes in lung allograft recipients (figure 2).
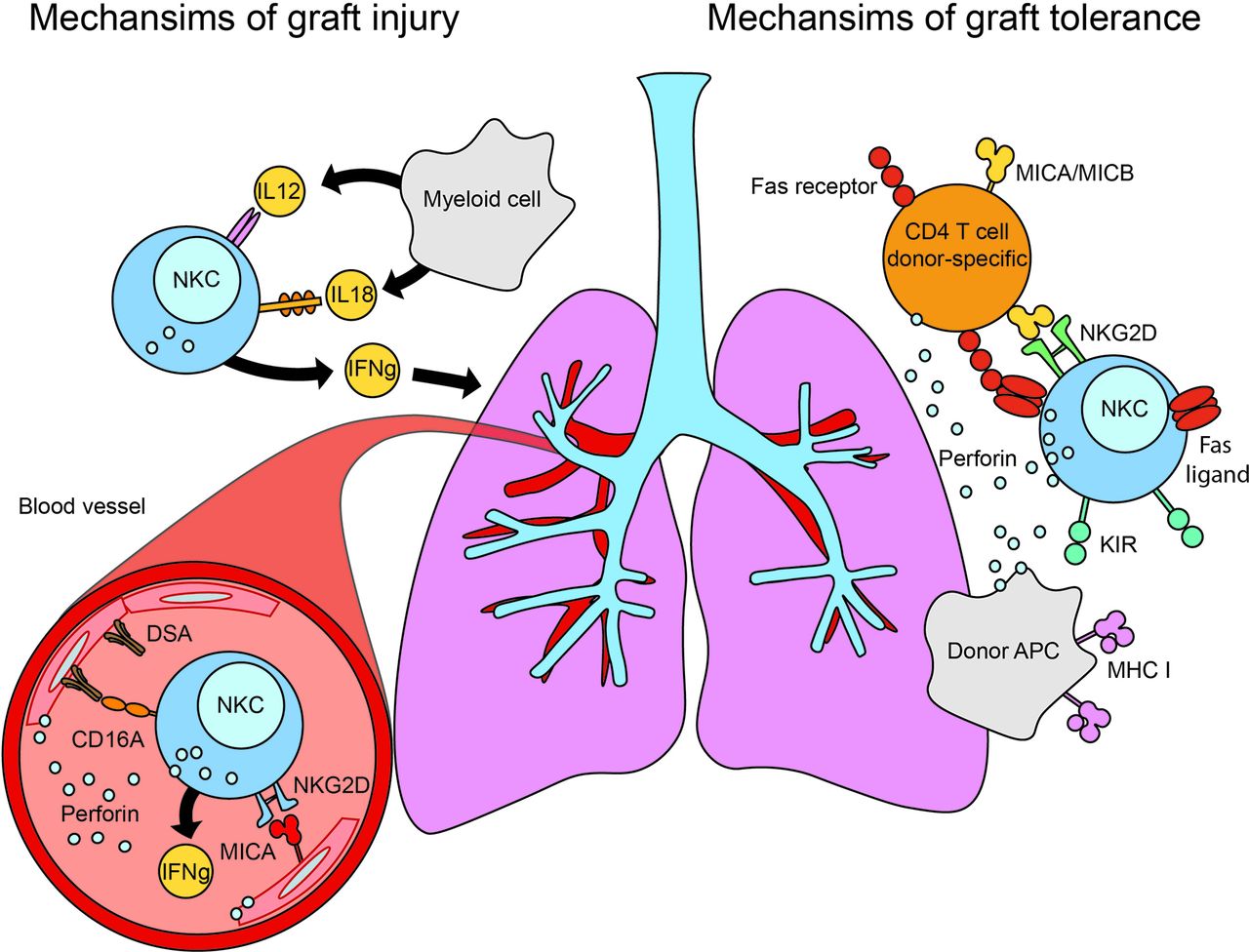
NK cells may mediate lung transplant outcomes. Overview of the potential roles that NK cells play in mediating lung allograft tolerance (right portion of figure) and lung allograft injury (left portion of figure). NK cells may promote allograft tolerance through detection of missing-self and direct cytotoxicity of donor APCs. Stressed donor-specific CD4+ T cells may also be eliminated through the NKG2D receptor on NK cells recognising MICA or other stress-induced ligands on activated T lymphocytes. Allograft injury may occur through several mechanisms, chief of which is perforin-dependent cell-mediated cytotoxicity, augmented by the production of proinflammatory cytokines such as IFN γ in response to IL-12 or IL-18. NK cells are also capable of antibody-dependent cell-mediated cytotoxicity by recognising antibody bound to the allograft (donor-specific antibodies (DSAs)) through the CD16A receptor on the NK cells. Finally, NK cells may also recognise and lyse stressed allograft cells (MICA or other NKG2D ligands on endovascular endothelial surface) through the NKG2D receptor on NK cells. APC, antigen-presenting cell; IFN γ , gamma interferon; IL, interleukin; KIR, killer cell immunoglobulin-like receptor; MHC, major histocompatibility complex; MICA/B, MHC class I chain-related sequence A and B; NKC, natural killer cell.
NK cells and their potential roles in promoting allograft tolerance
Much of NK cells’ ability to differentiate self from altered-self centres on the specificity of certain NK receptors to detect major histocompatibility complex (MHC) class I proteins on potential target cells.16 The NK cell repertoire of killer cell immunoglobulin-like receptors (KIRs) undergoes selection to tailor their recognition of host MHC class I ligands. NK cells are ‘licenced’ in the sense that recognition of self MHC class I molecules by inhibitory KIRs primes these cells to become more responsive towards cells that lack these MHC molecules, while potentially autoreactive NK cells lacking MHC class I receptors for self are rendered hyporesponsive.17 18 Viruses, particularly within the herpesviridae family, target and prevent expression of MHC molecules in the infected cells to avoid recognition by T cells,19 rendering these virus-infected, MHC class I-negative cells more susceptible to attack by NK cells.20
It has been postulated that a host-versus-graft NK cell interaction may promote tolerance in solid organ allograft recipients through depletion of donor antigen-presenting cells (APCs; figure 3).21 Direct allorecognition results when donor APCs display donor-specific MHC antigens and activate host alloreactive T cells.22 In turn, the frequency of these alloreactive T cells has been associated with kidney and lung allograft injury.23 24 In a mouse skin transplant model, graft-derived APCs were largely destroyed by donor NK cells. However, in the absence of NK cells, donor APCs survived and migrated to lymph nodes. Subsequently, alloreactive T cells were more frequent and resistant to costimulatory blockade, leading to decreased allograft survival.25 An analogous model of mouse orthotropic lung transplantation demonstrated improved graft survival from NK cell-mediated APC destruction.26
{kind=link}
{kind=link}
{kind=link}
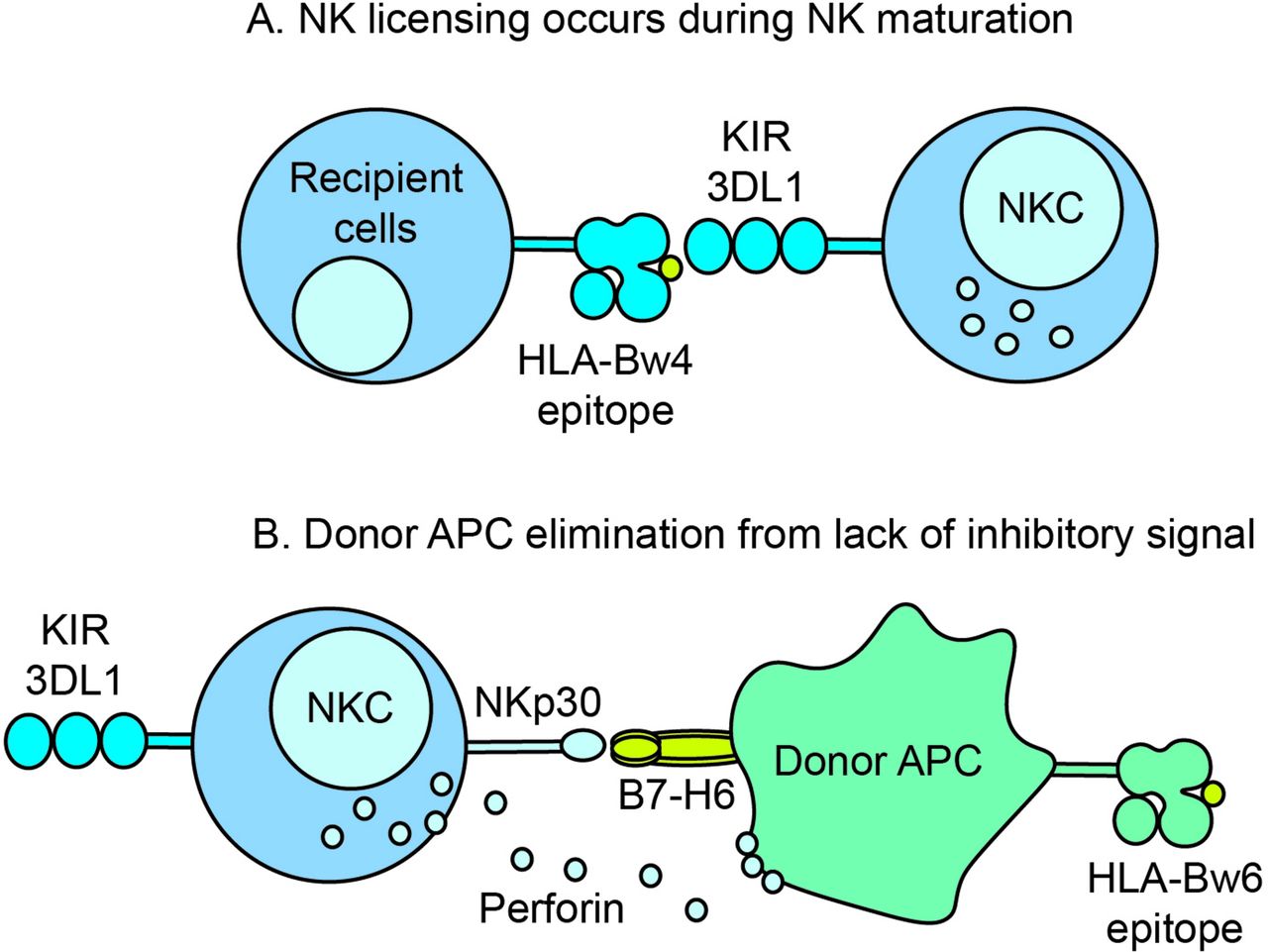
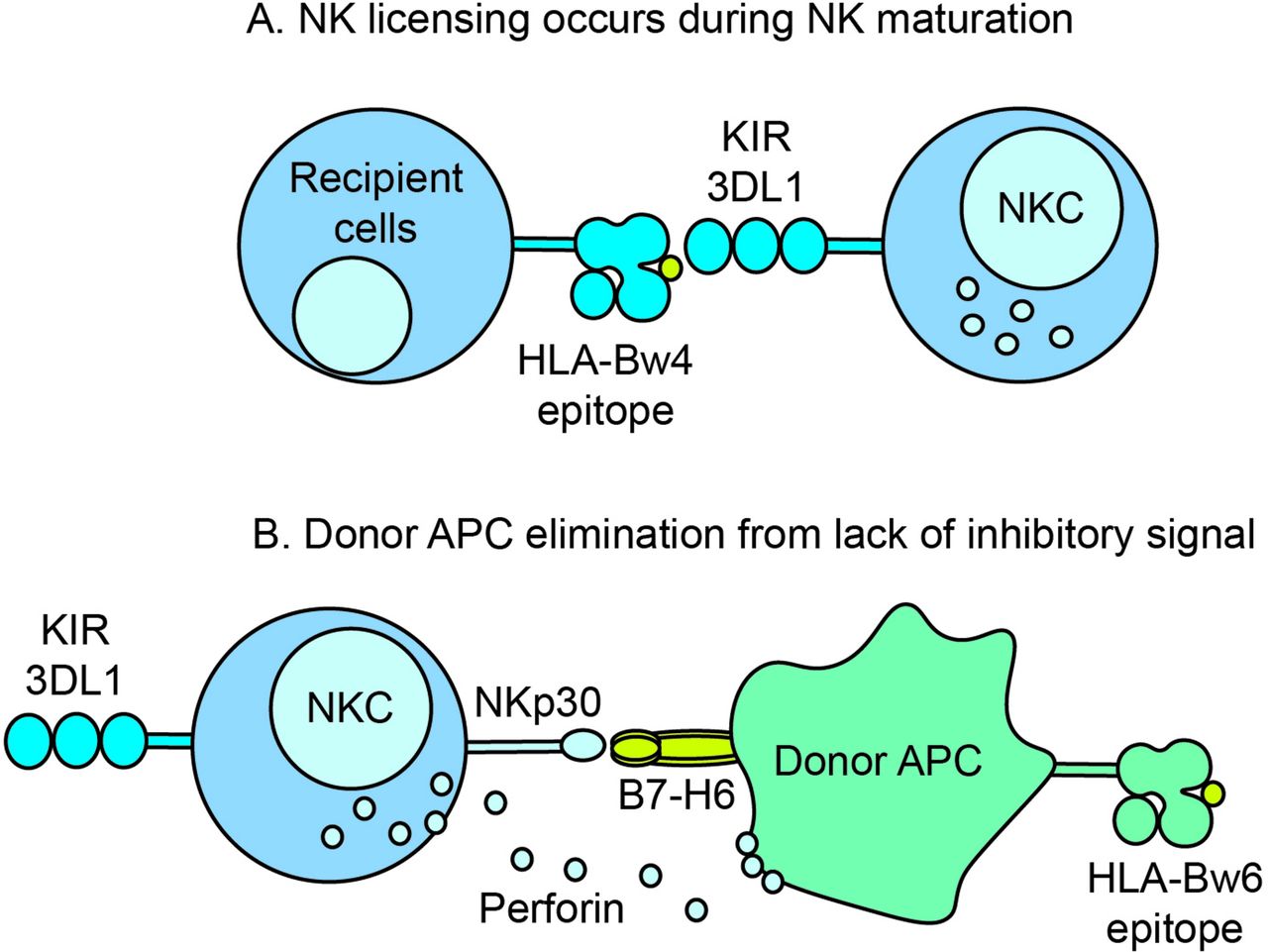
Possible mechanism for KIR licencing and destruction of donor antigen-presenting cells (APCs). (A) During NK cell maturation, inhibitory receptor KIR3DL1 is licenced by HLA antigens with the Bw4 epitope. (B) After transplantation, a KIR3DL1-Bw4 licenced recipient NK cell contacts a donor APC bearing Bw6 epitope. The absence of a KIR inhibitory signal triggers a host-versus-graft destruction of the recipient APC in conjunction with an activating receptor–ligand interaction such as NKp30 on NK cells and B7-H6 on APC. HLA, human leucocyte antigen; KIR, killer cell immunoglobulin-like receptors; NKC, natural killer cell.
In humans, the first evidence that NK cells in lung transplant are involved in allograft tolerance versus rejection stems from the observation that the KIR group A haplotype, which possesses more KIR genes encoding activating rather than inhibitory receptors, is a risk factor for chronic lung allograft dysfunction (CLAD).27 One strong MHC class I-inhibitory KIR licencing interaction occurs between KIR3DL1 and its ligands, human leucocyte antigen allotypes (HLA-A and HLA-B) containing the Bw4 epitope.28 KIR3DL1 does not recognise Bw6 HLA alleles. Subjects with KIR3DL1 and HLA-Bw4 who received HLA-Bw6 grafts, for whom a host-versus-graft effect could be present, had a decreased risk of CLAD or death, and decreased risk of early lymphocytic bronchitis.29 These findings suggest that the KIR3DL1 ‘licenced’ NK cells in the recipient may have depleted donor-derived HLA-Bw6+ APCs before they were able to induce an alloantigen-specific T cell response in the graft recipient.
NK cells might also promote graft tolerance through elimination of alloreactive T cells. Several mouse models of GVHD demonstrate that activated T cells might upregulate stress molecules that are recognised by activating NK receptors, leading to their elimination by host NK cells.30 In this setting, donor regulatory T cells were preserved relative to conventional CD4+ and CD8+ T cells, suggesting differential susceptibility to this effect,31 potentially because regulatory T cells secrete inhibitory factors such as TGF-ß that suppresses NK cell functions. How NK cells eliminate the activated donor T cells is not fully defined, but in one model system, NK cells have been shown to induce apoptosis of activated T cells via granzyme K.32
NK cells in antibody-mediated rejection (ABMR)
ABMR can arise from pre-existing donor-specific antibodies (DSAs), de novo DSA to HLA or MHC class I polypeptide-related sequence A and B (MICA/B) antigens, or autoantibodies to lung parenchymal self-antigens.33 34 Antibodies can bind to vascular endothelial cells and activate the classic complement pathway leading to cytotoxicity from membrane attack complex formation.35 Graft injury can also occur independent of the complement pathway, through antibody-dependent cell-mediated cytotoxicity (ADCC).36 ADCC relies on the bifunctional format of IgG antibodies. After the antigen-recognising fragment (Fab) binds to the surface of the target cell, the crystalline fragment (Fc) is free to interact with Fc receptors for IgG (FcγRs) on immune cells.37 There are several classes of FcγRs with multiple subclasses: the activating-only FcγRI (CD64) class on macrophages and neutrophils, the activating and inhibitory FcγRII (CD32) class on neutrophils and B cells and the low-affinity activating FcγRIIIa (CD16A) class present on NK cells and activated myeloid cells. Human NK cells are the only immune cells recognising Fc that generally do not coexpress an inhibitory FcγR, suggesting that they may be the predominant effector cell in ADCC.
The significance of ADCC to antibody responses has been demonstrated in the context of monoclonal antibody therapy and renal allograft rejection. The clinical efficacy of rituximab, a monoclonal antibody that recognises the CD20 antigen expressed on B cells, is correlated to NK cell frequency and function.38 In a mixed lymphocyte reaction-based assay, NK cells were shown to secrete IFNγ that was dependent on target-specific alloantibodies and could be inhibited by blocking the CD16A receptor.39 CD16A is encoded by the FCGR3A gene, for which a common genetic variant is a phenylalanine (F) for valine (V) substitution at position 158. When this polymorphism is present, the 158V homozygotes (VV) have significantly increased binding affinity for IgG compared with 158F homozygotes (FF).40 In a study of the efficacy of trastuzumab in HER-2/neu-positive metastatic breast cancer, subjects with low-affinity polymorphisms (158 FF) had worse progression-free survival.40 In lung transplant, a conference abstract reported lung allograft recipients with the high affinity 158 VV genotype had an increased risk for CLAD or death, although it is unknown if this reflected increased ABMR.41 Other studies in renal allograft recipients demonstrated NK cell-associated gene transcripts specific to CD16A were increased in renal ABMR.42
NK cell role in graft-specific cytotoxicity
In humans, NK cells have also been associated with lung allograft injury. A greater concentration of NK cells in bronchoalveolar lavage (BAL) fluid has been observed during acute cellular rejection, even though the NK cell percentage of total BAL lymphocytes decreased.43 In subjects with CLAD, NK cell peripheral blood frequencies were decreased, but these NK cells had a more activated phenotype.43 CLAD subjects also have higher numbers of NK cells in allograft transbronchial biopsy specimens.44 There are likely multiple mechanisms for the presence of NK cells in the lung during graft injury: NK cells may be bystanders and trafficking to the lungs in response to humoural or T cell-mediated inflammation or they could be causing direct graft injury from recognition of ‘missing-self’ or by NK cell surveillance of ‘stressed-self’ in allograft lung cells.
Allograft recipients with mismatched donor MHC KIR ligands may be at increased risk of later allograft cytotoxicity from recipient NK cells’ failure to recognise MHC class I molecules via inhibitory KIR leading to ‘missing-self’ cytotoxicity. While in lung transplantation donor HLA types that fail to bind to inhibitory KIR on the recipient’s NK cells have been associated with better outcomes, the opposite has been reported in the context of renal transplantation. In a study of 174 cadaveric renal allograft recipients, worse outcomes were seen in the absence of inhibitory NK cell interactions (either donor HLA-Bw4 with recipient KIR3DL1 or donor HLA-C2 group with recipient KIR2DL1).45 The reason for the difference between organ types is not entirely clear but could reflect the relative importance of inhibitory NK cell interactions in preventing ‘missing-self’ activation of NK cells in the context of DSAs or other NK cell activation signals from the renal allograft.45
In contrast with the KIR family of molecules that recognise missing-self through MHC class I ligands, the NKG2D receptor is activated in response to ‘stressed-self’ cells undergoing damage. MHC class I chain-related A and B (MICA and MICB) and UL-16-binding proteins 1–6 are NKG2D ligands absent or expressed at low levels in many healthy cells. Under stress from malignancy, infection or ischaemia-reperfusion injury, surface expression of these ligands increases.46 47 On binding these ligands, NKG2D-activated NK cells lyse the target cell. In a study of renal allograft recipients, elevated levels of NKG2D mRNA in biopsy specimens were associated with acute and chronic allograft rejection, and NKG2D+ cells were observed within the tubulointerstitial spaces.48 However, because CD8+ T cells and γδ T cells also express NKG2D and other NK cell transcripts, this ‘stressed-self’ response may not be entirely attributable to NK cells.49 Antibodies to allogeneic MICA proteins have been associated with both renal allograft rejection and CLAD, possibly through augmentation of complement-binding or increasing opsonisation and antigen presentation.50
In a mouse heterotopic tracheal transplant model, airway fibrosis was inhibited by depletion of NK cells. In this study, the mouse NKG2D ligand Rae-1 had increased expression in allografts, overexpression of Rae-1 resulted in severe luminal fibrosis even in the absence of T or B cells and an antibody blocking NKG2D prevented this luminal fibrosis.51 Notably, these findings contrast with the study by Jungraithmayr et al,26 where augmentation of NK cells by IL-15/IL-15Rα complex therapy before transplant resulted in depletion of APCs and protection from rejection pathology. Together these two studies suggest that magnitude, timing and context of NK cell responses may differentiate between protective versus destructive graft responses. Finally, there is evidence that allograft recipients can experience a protective effect from circulating soluble MICA. A study of heart allograft recipients found that high levels of soluble MICA were associated with preserved graft function, an effect attributed to internalisation of the NKG2D receptor.52
NK cells and cytomegalovirus (CMV) infection: the role of innate memory responses
Lung allograft recipients have higher rates of CMV infection relative to other solid organs, with significant CMV-associated complications despite effective antiviral treatment.53 54 CMV infection can induce proinflammatory cytokine release, drive antibody-mediated and cell-mediated cytotoxicity and enhance immune complex deposition.55–57 Cross-reactive antigens between CMV and the allograft might also induce heterologous immunity.58 NK cells are key effectors during CMV infection, where they eliminate CMV-infected cells through ADCC and direct cytotoxicity, as well as modulate T and B cells through the secretion of IFNγ.59 60
In humans, the CD94/NKG2C receptor identifies a largely CMV-specific NK cell population. For example, CMV-seropositive individuals have consistently higher proportions of NKG2C+ NK cells relative to CMV-naïve subjects.61 This NKG2C receptor belongs to the C-type lectin-like family of receptors expressed by T cells and NK cells. NKG2C covalently bonds with the CD94 glycoprotein and associates with the DAP12 signalling adapter, which contains immunoreceptor tyrosine-based activation motifs (ITAMs). Together, this complex recognises the invariant HLA-E protein as a ligand.62 63 How NKG2C+ NK cells recognise CMV-infected cells has not as yet been defined. Alterations in HLA-E stability and HLA-E-associated molecules presented during CMV infection might be recognised by NKG2C+ NK cells, but a dominant CMV peptide epitope directly presented by HLA-E seems unlikely.64
NKG2C+ NK cells expand following CMV reactivation or viraemia in solid organ transplant recipients and may contribute to control of CMV viraemia.65 Elevation in NKG2C+ NK cell frequency has been shown to persist for over a year following acute CMV infections. NKG2C+ NK cells were increased in recipients of haematopoietic stem cell (HSC) transplantation who reactivated CMV.66 These NKG2C+ NK cells were more mature, expressed CD57, had increased cytokine production on stimulation through CD16A and had increased function during CMV viraemia when arising from seropositive donors rather than seronegative donors, suggesting memory-like capabilities.65 66
In lung transplant subjects, we and others have observed increased frequencies of NKG2C+ NK cells in peripheral blood and BAL of subjects with CMV+ donors.67 68 Bayard et al further followed the subjects with CMV reactivation for 2 years and found higher frequencies of NKG2C+ NK cells in this group compared with those without CMV reactivation. Following reactivation, the NKG2C+ NK cell population continued to expand but apparently afforded no protection from recurrent viraemia.67 Furthermore, in a prospective study of 130 lung transplant recipients, we found that BAL NKG2C+ NK cells increased prior to CMV viraemia and that subjects with increased frequencies of BAL NKG2C+ NK cells were at increased risk for CLAD or death.68 A significant proportion of humans possesses a null allele of the gene (KLRC2) encoding NKG2C. In a study of 98 lung transplant recipients, subjects homozygous for the expressed allele of the KLRC2 gene had greater freedom from CMV viraemia and disease compared with subjects carrying the null allele.69 Together, these data suggest the NKG2C+ NK cell population, combined with T and B cell responses, provide essential but incomplete protection against CMV in lung transplant recipients and that quantification of this population may provide insight into the magnitude of ongoing CMV-related inflammation in the graft.
NK cells respond to other viral and bacterial infections
Lower respiratory tract viral, bacterial and fungal pathogens are all risk factors for CLAD.70 A primary function of NK cells is to respond quickly and specifically to infection. In addition to KIR and NKG2 receptor families, the natural cytotoxicity receptors (NCRs) are a family of activating receptors that are potentially important in tumour recognition and to viral infections. NKp46 and NKp44 are NK cell Ig-like transmembrane glycoproteins associated with the ITAM-bearing signalling adapters CD3ζ and FcεRIγfor NKp46 and DAP12 for NKp44.71 It has been reported that human NKp44 and human and mouse NKp46 recognise influenza viral haemagglutinin (HA) as a ligand. O-glycosylation of the sialic acid carrying residue threonine on NKp46 may be important for the recognition of HA,72 although it is unclear how sialic acid renders NKp46 preferentially able to bind HA.
The role of NK cells in limiting influenza infection is unclear. Mice deficient in the Ncr1 gene encoding NKp46 were reported susceptible to lethal influenza infection.73 However, in other studies and our unpublished observations, the total depletion of NK cells in mice had no effect on pulmonary influenza titres or influenza-associated morbidity or mortality, although an increase in NK cells was detected in the lung after influenza infection.74 In human paediatric subjects with severe influenza, peripheral blood CD56dim NK cell frequencies were decreased.75 A study examining NK cell surface markers in healthy controls and subjects with moderate-to-severe influenza found decreased frequencies of NKp46+NKp44− NK cells but increased frequencies of NKp46+NKp44+ NK cells in severe influenza cases.76 In lung transplant subjects, we found increased NK cells in BAL during pulmonary bacterial and viral infection, higher NK cell turnover and greater frequencies of mature NK cells defined by increased KIR and CD16A and decreased CD94/NKG2A (an inhibitory receptor for HLA-E) (unpublished). Together, these data suggest that NK cells undergo changes and likely traffic to the lung during influenza infection, but their contribution to controlling infection is ill defined.
Epstein-Barr virus (EBV) is another important transplant-associated infection. Allograft recipients are at increased risk for EBV-associated post-transplant lymphoproliferative disorder (PTLD) as the immunosuppression regimens necessary to prevent allograft rejection also suppress antiviral T cell immune surveillance. NK cells have been demonstrated to expand during acute EBV infection77 and may be more effective at killing EBV-infected cells when they express the NKG2A+KIR− phenotype using activating DNAM-1 and NKG2D receptors to mediate the recognition.78 Compared with cells from healthy non-transplanted and thoracic transplant controls, NK cells from thoracic organ transplant recipients with PTLD had decreased NKG2D and elevated PD-1 expression, potentially consistent with an exhausted phenotype.79
Historically, NK cells have been considered important in mediating responses to viral infections, but there is also evidence of their role in the host response to bacterial infections.15 Cystic fibrosis accounts for around 16% of lung transplants internationally, and Pseudomonas aeruginosa is a major infectious complication both pretransplantation and post-transplantation for this group.80 During in vivo and in vitro infection in mice, P. aeruginosa induces NKG2D ligands on pulmonary epithelial cells.81 In a mouse model of P. aeruginosa infection, increased expression of the NKG2D ligand Rae-1a increased pulmonary clearance of bacterial cells, cellular phagocytosis and survival.82 Furthermore, NK cells stimulated via NKG2D were shown to be a primary source of IFNγ in the lungs in response to infection.
NK cells and the impact of transplant-specific immunosuppression
Lung allograft acceptance is dependent on aggressive immunosuppression regimens, which can be divided into induction and maintenance therapies. Induction regimens often include corticosteroids, cell cycle inhibitors such as mycophenolic acid (MPA) and azathioprine and antibodies such as basilixumab or antithymocyte globulin. Modern maintenance immunosuppressant regimens typically contain a calcineurin inhibitor (CNI), most commonly tacrolimus, a cell cycle inhibitor, a corticosteroid and possibly an mammalian target of rapamycin (mTOR) inhibitor (rapamycin or everolimus). The effects of these immunosuppressants on T cells are well characterised, but their impacts on NK cells are less well defined.
Basiliximab, a blocking antihuman monoclonal antibody against the IL-2Rα chain (CD25), and alemtuzumab, a cell-depleting monoclonal antibody against CD52 found on mature lymphocytes, are used in human solid organ transplant induction regimens. One study showed that treatment with alemtuzumab or basiliximab reduced the absolute count of T lymphocytes up to 90-fold, with a smaller, fourfold reduction in NK cells, following treatment early after transplantation.83 However, these differences may derive from faster reconstitution of NK cells relative to T cells. Alternatively, basiliximab therapy might promote NK cell proliferation by freeing IL-2 from T cells that express the high-affinity CD25-containing IL-2 receptor.84 Evidence for this effect arises from studies in subjects treated with daclizumab, another blocking humanised monoclonal antibody against CD25. Daclizumab treatment significantly expanded the CD56bright NK cell population in vivo.85 In vitro studies showed increased NK cell proliferation and cytotoxicity, attributed to the increased availability of IL-2.86
The CNIs, cyclosporine and tacrolimus reduce production of IL-2 by inhibiting the transcription of nuclear factor of activated T cells within the calcineurin pathway. This pathway is present in NK cells, but there is mixed evidence as to the effect of CNIs on the function of NK cells. Some studies report that NK cells cultured in the presence of cyclosporine or tacrolimus may indeed retain their cytolytic activity, possibly because of storage of previously synthesised effector molecules, perforin and granzymes87; however, other studies show a decrease in the cytotoxic activity of NK cells with CNI.88 89 There is also mixed evidence as to the degree that NK cell degranulation is affected by inhibition of this pathway.83 90 In vivo, other immunosuppressants such as MPA or rapamycin may also affect NK cell function. Two studies show reduction in the expression of NKG2A, reduced expression of NCR and NKG2D receptors and diminished cytotoxicity against targets in subjects taking MPA or rapamycin.83 87
Treatment of NK cells with corticosteroids results in decreased expression of activating receptors and decreased granule exocytosis via inhibition of ERK1 and 2 signalling pathways. In vitro corticosteroid effect on NK cell survival, proliferation and cytotoxicity are additionally dependent on stimulation with either IL-2 or IL-15.91 Finally, CNIs, corticosteroids and MPA have all demonstrated reduced NK cell proliferation in vitro, mostly in a dose-dependent fashion.83 88 90
Summary: NK cell biology important to lung allograft outcomes, knowledge gaps and future directions
In summary, there is increasing evidence that NK cells play critical and sometimes opposing roles modifying outcomes following lung transplantation (table 1). NK cells can promote tolerance by targeting donor APCs and alloreactive T cells. Conversely, NK cells can augment rejection though recognition and elimination of ‘non-self’ and ‘stressed’ cells or enhancing ABMR.
NK cells and solid organ transplant outcomes
A major limitation in our understanding of the role of NK cells in lung transplant cohorts is the reliance on extrapolation from studies of mice and non-lung allograft recipients. Table 1 summarises the sources of evidence for proposed roles of NK cells in lung transplant. Findings in human lung transplant recipients may differ due to differences in immune activation, infection susceptibility and immunosuppressant use. For example, in lung transplant recipients, the magnitude of the positive association between long-term outcomes and KIR/HLA mismatches that promote APC destruction is less robust than what has been observed in HSC transplant cohorts, potentially because of counterbalancing injurious host-versus-graft NK cell effects or differences in T cell activation.92
In current clinical practice, enumeration of NK cells may be useful in distinguishing rejection from infection, while detection of elevated numbers of NKG2C+ NK cells might identify patients at risk for CMV infection. If constraints on donor matching could be reduced through strategies to maintain lungs ex vivo, matching to augment host-versus graft NK cell function and thereby reduce antigen presentation could be considered. At the same time, limiting NK cell-mediated graft injury could also be beneficial. Further study of the dynamics of NK cell function in lung allografts may lead to strategies to promote tolerogenic NK cell functions while reducing allograft toxicity.
References
Footnotes
Contributors All individuals who meet authorship criteria are listed as authors, and all authors certify that they have participated sufficiently in the work to take responsibility for the content, including participation in the concept, design, writing or revision of the manuscript.
Funding This study was supported by Career development award IK2CX001034 from the VA Office of Research and Development Clinical Sciences Research & Development Service.
Competing interests None declared.
Patient consent Not required.
Provenance and peer review Not commissioned; externally peer reviewed.