Article Text

Abstract
Respiratory syncytial virus (RSV) is the most common single cause of respiratory hospitalisation of infants and is the second largest cause of lower respiratory infection mortality worldwide. In adults, RSV is an under-recognised cause of deterioration in health, particularly in frail elderly persons. Infection rates typically rise in late autumn and early winter causing bronchiolitis in infants, common colds in adults and insidious respiratory illness in the elderly. Virus detection methods optimised for use in children have low detection rate in adults, highlighting the need for better diagnostic tests. There are many vaccines under development, mostly based on the surface glycoprotein F which exists in two conformations (prefusion and postfusion). Much of the neutralising antibody appears to be to the prefusion form. Vaccines being developed include live attenuated, subunit, particle based and live vectored agents. Different vaccine strategies may be appropriate for different target populations: at-risk infants, school-age children, adult caregivers and the elderly. Antiviral drugs are in clinical trial and may find a place in disease management. RSV disease is one of the major remaining common tractable challenges in infectious diseases and the era of vaccines and antivirals for RSV is on the near horizon.
- viral infection
- innate immunity
- paediatric lung disaese
- copd exacerbations
Statistics from Altmetric.com
Introduction
Respiratory syncytial virus (RSV) is the most common single cause of respiratory hospitalisation of infants and is the second biggest cause of lower respiratory infection mortality worldwide.1 In resource-rich countries, RSV causes relatively few deaths among otherwise healthy children but places a great strain on hospital resources in winter seasons. In healthy adults, RSV causes common colds reinfecting with apparent ease; in frail elderly persons, it causes insidious deteriorations of respiratory health with high mortality, much of which may be undiagnosed. Despite intensive research since the 1960s, vaccines and specific therapies remain unavailable. Furthermore, using burden-adjusted research intensity, RSV research is underfunded and has seen a decline in spending during a period when influenza research has been progressively better funded.2 In this review, we examine the epidemiology, immunology and contemporary research landscape of RSV disease in both children and in adults.
Epidemiology
In the developed world, RSV is the single largest cause of hospitalisation in under-5s, with 33 500 admissions in the UK, and most children infected by 2 years of age.3 4 Outbreaks of RSV disease occur each winter in temperate regions, normally beginning in autumn and early winter in Europe and North America.3 5 National data from the UK shows that infections typically start to appear around week 40 (September) and peak between week 46 and 52 (November–early December).6 In summer, there are few cases; for example, in week 18, the number of reported cases in England is typically approaching zero. This seasonality is mirrored in the USA where the median week of onset ranged from week 46 to week 3 (November–January).6 7 There has been a rise in the proportion of infants admitted to hospital with RSV from around 2% in 1998 to 5% today.4 This may be due to more infants surviving at early gestational ages and/or a lower threshold for admission of infants with respiratory distress.
In affluent countries, deaths due to RSV infection are very rare in otherwise healthy children. Mortality rates are greatest in the first few months of life and decrease as childhood progresses.8 However, findings from the Pneumonia Etiology Research for Child Health PERCH) study implicate RSV in development of pneumonia in the under-5s. In this study of children with pneumonia, RSV was found in 31.1% of all cases, and three times more common than the next most predominant pathogen.9 In a Spanish study, mortality was estimated to be 6.19 per 100 000 in children under 1 year of age, decreasing by 50% each year until a plateau around 4 years of age (0.79 in 1–2 years of age; 0.32 in 2–3 years; 0.19 in 3–4 years and 0.19 in children over 4 years).8
Most of the global RSV-associated child mortality occurs in low-income countries with 99% of deaths occurring in these countries, most in children under 6 months of age.10 Globally, there are an estimated 33.1 million cases of RSV-associated acute lower respiratory tract infection (LRTI) per year, with 3.2 million hospital admissions and 59 600 hospital inpatient deaths.10 When including RSV infections outside the hospital setting, mortality estimates almost double to 118 200.10 Furthermore, at least 28% of RSV-associated mortality occurred in children with severe comorbidities (such as congenital heart disease) in low-income countries.11 Despite this, health inequality means that only 24% of children in low-income and lower middle-income countries have access to paediatric intensive care, an important reason for the differences in mortality.11
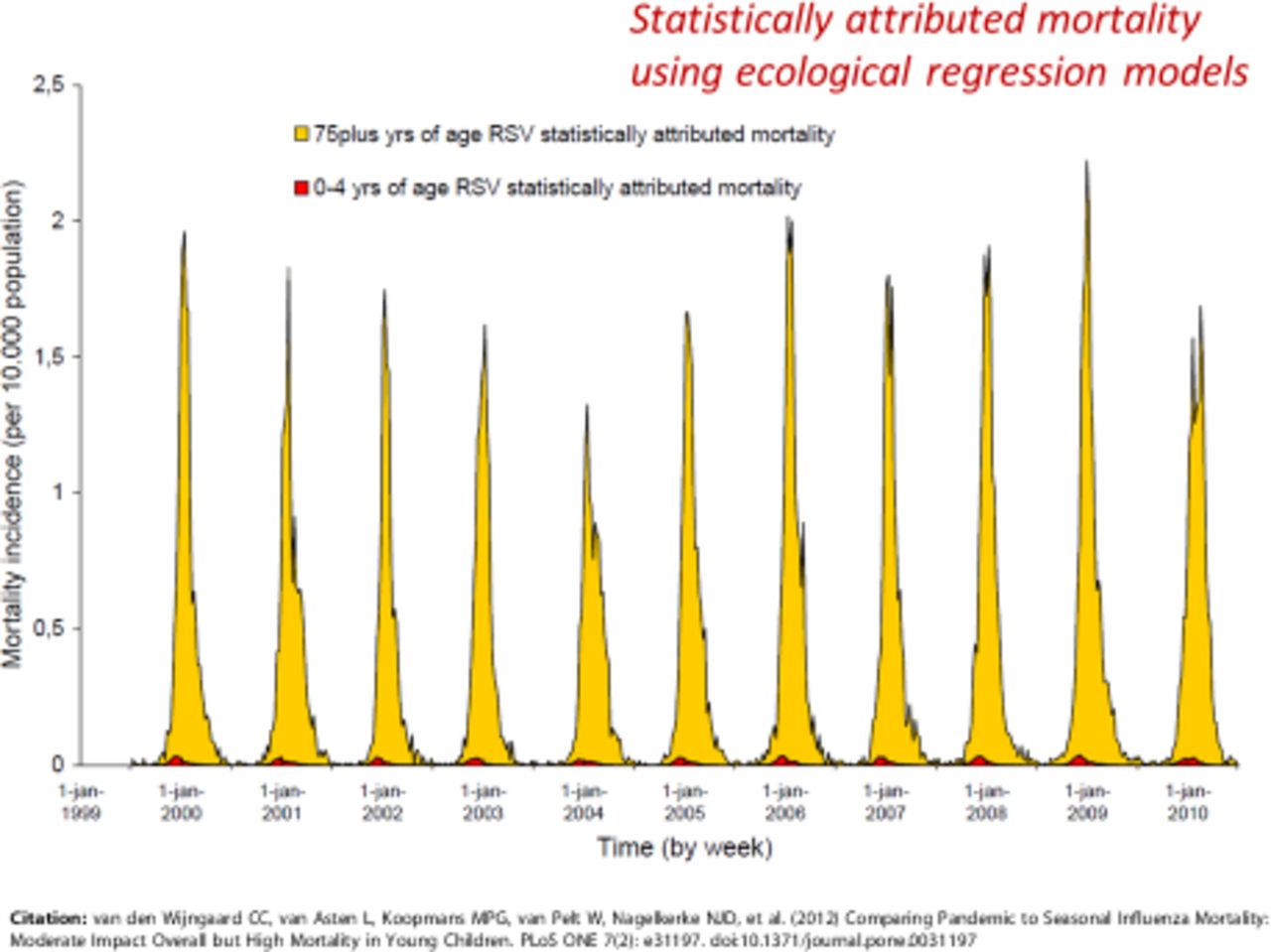
In adults, there are 8482 deaths per year attributable to RSV in the UK, with 93% of those occurring in adults aged more than 65 years.12 Deaths due to RSV respiratory disease increase after age 49, rising from 4.2% of all respiratory disease deaths in adults aged 18–49 years to 5.9% in adults aged 50–64 years, 5.7% in elders aged 64–74 years and again reaching 5.9% in elders aged more than 75 years.12 Mortality in these groups also rises from 1 per 100 000 in adults aged 18–49 years to 155 per 100 000 in elders aged over 75 years.12 It is important to note that RSV infection may be greatly underdiagnosed in adults due to delayed and insidious disease and the relatively low viral load (especially if only upper respiratory tract samples are tested).13 Epidemiological studies show a much larger number of inferred RSV-associated deaths than those that might be established by direct testing and viral detection.11 In a study from the Netherlands using statistical methodology to estimate seasonal mortality of the total Dutch population of 16.5 million, RSV-attributable mortality could be inferred in different age groups.14 Projected RSV-attributable deaths in the paediatric age group were very rare indeed compared with RSV-attributable deaths in older adults (figure 1); based on data from Wijngaard et al 14 with permission.

Estimated respiratory syncytial virus (RSV)-attributable mortality in the Netherlands according to age. Excess specific mortality in children aged 0–4 years (red) compared with adults aged 75 years and over (yellow) over 11 successive winters (1999/2000 to 2010/2011). Figure based on data from Wijngaard et al,14 with permission.
Risk factors
Age remains the biggest risk factor for bronchiolitis, young children having small-diameter airways, impaired respiratory capacity and low respiratory reserve.15 Risk is greatest at 1 month of age and decreases thereafter.16 Exposure to tobacco smoke and lack of breastfeeding have important additional effects though conclusions are limited by small sample size in these studies.17 Risk factors such as male sex, prematurity, congenital heart disease and underlying pulmonary disease are also significant.18 19 Another strong risk factor is the presence of older siblings.20 Older siblings are a greater source of spreading infection than adults as they spend significant time in nurseries and schools where RSV can spread with ease.
In elderly persons, the greatest risk factors are pulmonary disease (especially COPD) and functional disability as measured by activities of daily living.21 Interestingly, coronary artery disease and diabetes (both risk factors for severe influenza) are not associated with increased risk of RSV severe disease.21 RSV is associated with significant morbidity and mortality in care home residents, with 12% of all adult RSV admissions occurring in this group and with a mortality rate of 38% compared with 3% in patients admitted from the community.22
Clinical diagnosis
Children
Viral bronchiolitis is one of the most common infant viral illnesses; RSV infection causing about 70% of cases. Bronchiolitis typically develops after an initial prodrome of nasal congestion, cough and coryza that can last for up to 3 days.23 24 Subsequently, low-grade fever, wheezing, crepitations on auscultation and signs of increased respiratory effort develop, such as nasal flaring, chest wall retraction and tachypnoea.23 In very young infants under 6 weeks of age, apnoea associated with RSV infection is an important indication for hospital admission.25 In most children, however, reduced oral intake and hydration status are the main indications for admission. Other reasons for hospital admission include hypoxia and respiratory failure. RSV infection is also associated with otitis media.26
In hospitalised infants, RSV infection is associated with pulmonary infiltrates/atelectasis (42.8%), otitis media (25.3%), hyperinflation (20.8%), respiratory failure (14%), hyperkalemia (10.1%, defined as K+ >6.0), apnoea (8.8%) and bacterial pneumonia (7.6%). Rarer complications included anaemia (6%), sepsis (5.9%), seizures (1.8%) and meningitis (0.2%, coinfection with S treptococcus pneumoniae).27
Adults
In young adults, RSV typically manifests as upper respiratory tract infections with mild to moderate symptoms, only very rarely causing severe disease.13 However, severe complications may follow in frail elderly people with respiratory or cardiac comorbidity.13 Fever occurs in approximately 50% of cases but rarely reaches >38°C; cough occurs in over 90% and wheeze in around 40%. Wheeze in the absence of a history of asthma or other lung disease raises the suspicion of RSV infection.13 Compared with infants, adults are much less likely to test positive for RSV, present later, have lower viral titres and have lower yields on routine diagnostic testing.28 This leads to great difficulty in ascertainment of RSV as the original cause of deteriorating health in frail elderly persons, similar to the elusive cat called Macavity described by TS Eliot in Old Possum’s book of practical cats (box 1).
Adults are less likely to test positive for RSV, drawing similarities with the elusive cat called Macavity from TS Eliot's Old Possum's book of practical cats.
He’s outwardly respectable (they say he cheats at cards).
And his footprints are not found in any file of Scotland Yard’s.
And when the larder’s looted, or the jewel-case is rifled.
Or when the milk is missing, or another Peke’s been stifled.
Or the greenhouse glass is broken, and the trellis past repair.
Ay, there’s the wonder of the thing! Macavity's not there!
From TS Eliot, Old Possum’s Book of Practical cats, Faber and Faber, 1939.
In the elderly, RSV can also lead to viral-associated pathology with a different profile of complications to infants. One study in New York state found RSV associated with over 10% of LRTIs, 11.4% of COPD exacerbations, 7.2% of asthma exacerbations and 5.4% of congestive heart failure exacerbations.22
Viral diagnosis
RSV isolation from tissue culture was originally the gold standard for RSV diagnosis, but may take several days to report ; the cost and reliability of processing the sample have led to this method being largely superseded.29 Antigen detection assays such as direct immunofluorescence assays, enzyme immunosorbent assays, optical and chromatographic immunoassays have become popular,29 but despite their convenience they are inappropriate for diagnostic use in older children and adults with a history of previous infection. In such cases, the antigen load is much lower; sensitivity ranges from 72% to 94% and specificity 95%–100%, in children up to 32 months of age, but sensitivity is 0%–25% in older children and adults.30 Nucleic acid assays, such as real-time PCR, have superior sensitivity and specificity compared with antigen detection assays and tissue culture, but adults are still less likely to have a positive result.31 While sensitivity is greatly reduced in older patients, they remain the most sensitive detection method available.30
Virology of RSV
Virus structure
RSV is a single-stranded negative sense virus, genome being held within a nucleocapsid surrounded by a lipoprotein envelope.32 The viral RNA encodes 10 genes encoding 11 proteins.32 The major surface glycoprotein (G) facilitates virus attachment and the fusion (F) protein mediates virus–cell fusion and cell–cell fusion into syncytia.32 The third envelope protein is small hydrophobic (SH) protein. Other RSV proteins include matrix (M) proteins M2-1 and M2-2 which regulate transcription, nucleoprotein (N), phosphoprotein (P) and large nucleoprotein (L)—RNA polymerase, which are located in the nucleocapsid. The non-structural (NS) proteins, NS1 and NS2, have immunomodulatory functions.32 The structure of the virus and genome order is shown diagrammatically in figure 2, from Battles and McLellan.33
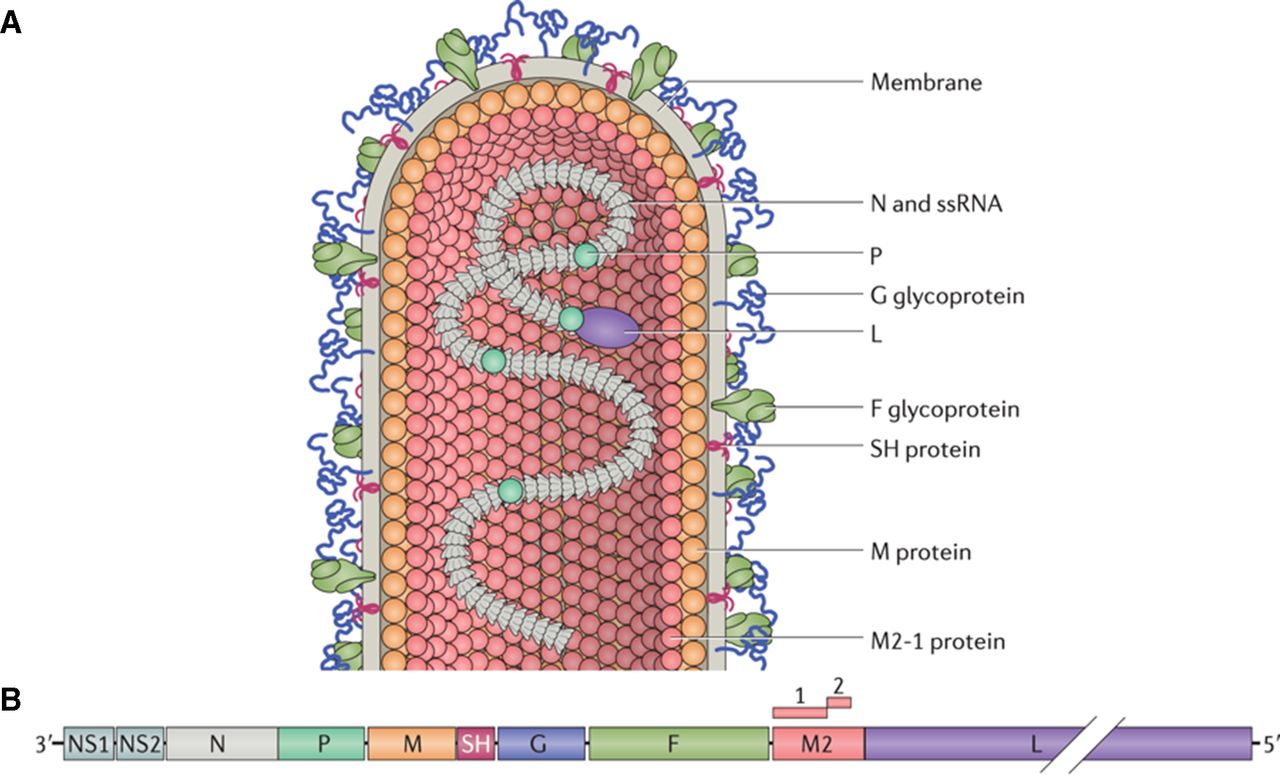
Structure of RSV. (A) The infectious form of the virus is filamentous, the structure bearing attachment G and F proteins embedded in the viral membrane. The M protein lies underneath the viral membrane. The L and the P are associated with viral RNA. (B) The viral genome contains 10 genes encoding 11 proteins (M2 gene encoding the M2-1 and M2-2). The most abundantly transcribed genes are those at the 3′ end, encoding NS protein 1 (NS1) and NS2, which inhibit apoptosis and antiviral responses. Source file: F, fusion protein; G, glycoprotein; L, large nucleoprotein; M, matrix protein; NS, non-structural protein; P, phosphoprotein; RSV, respiratory syncytial virus; ssRNA, single-stranded RNA (reproduced with permission).33
Recent knowledge of the structure and conformational changes of the envelope glycoprotein F has had a major impact on vaccine development. RSV-F exists in several forms, the prefusion form (preF) undergoes a conformational change after binding its target, allowing insertion of the F protein into the host cell. It again changes into a more stable and elongated form (postF) where the virus and host membranes are fused.34 In man, the most neutralising antibody is directed against the Ø and V antigenic sites on RSV PreF, as measured by Luminex-based assays34 and antibodies directed against RSV-F prevent epithelial cell binding.35 36 PreF vaccines aim to conserve these highly immunogenic sites.34
Karron et al, using RSV mutants, showed that changes in F protein alone affect virus penetration, protein assembly and subsequent virus release.37 Like RSV-G, the F protein has the capacity to bind to glycosaminoglycans (such as immobilised heparin), but its capacity is far less than that of G protein, and its exact ligand remains unclear.38 39 Intercellular adhesion molecule 1 (ICAM-1) has been proposed as a ligand as monoclonal antibodies directed against ICAM-1 significantly reduced RSV infection.40 Nucleolin has been identified as a potential ligand.41
Hallek et al demonstrated the ability of RSV-G to bind proteoglycans (GAGs) present on cell surfaces and thus facilitate virus attachment.42 Its role as a low-affinity receptor was confirmed when RSV-cell binding was shown to occur in its absence.43 While various GAGs have been identified, the protein ligand for RSV-G has not been identified either. The most promising candidate is CX3CR1, which is expressed on human airway epithelial cells (AECs).44 45
In addition, RSV-G also modulates the host immune response in various ways; the presence of RSV-G leads to a reduction in CX3CR1+ T cell migration to the lung and in Class I restricted interferon (IFN)-ϒ+ T cells specific for RSV.46 47 More dramatically though, RSV-G reduces the number of DX5+ natural killer (NK) cells (an early response cell in viral infections), neutrophils and CD11b-expressing cells (a regulator of leucocyte adhesion).48 G protein attenuates type I IFN responses via toll-like receptors on epithelial cells and plasmacytoid dendritic cells (pDCs) and also reduces IFN-ϒ production from T cells via the same mechanism.49 RSV-G has been targeted as a vaccine antigen, particularly as primary RSV infection generates strong anti-G antibody responses; preclinical trials have shown promising neutralising antibody responses.36 Phase II trials are also underway using MVA-BN-RSV, a vector model incorporating various proteins including RSV-G.
The SH protein forms viroporins once inside the target cell which permeabilise host cell membranes to facilitate viral spread.50 SH protein also has various immunomodulatory features. SH blocks tumour necrosis factor-α signalling via nuclear factor kappa light chain enhancer of activated B cells (NF-κB) and thus inhibits apoptosis of virus infected cells.51 In addition to promoting apoptosis, NF-κB is a master regulator of many proinflammatory cytokines in antigen-presenting cells as well as promoting antigen presentation through upregulation of transporter associated with antigen processing (TAP1), major histocompatibility complex class I (MHC-I), CD40 and CD86.52 Pollock et al showed that the SH protein attenuates production of NF-κB dependent cytokines and that SH protein also leads to reduced numbers of CD3+ IFN-ϒ+ cells.53 SH is harder to detect on the virion compared with F and G proteins but was recently incorporated into a vaccine targeting infected cells rather than virions, via Fcϒ receptors.54 This vaccine candidate, DPX-RSV, is currently undergoing phase I trials.
NS1 and NS2, the non-structural proteins of RSV, play a key role in virus immune evasion, primarily through inhibition of type 1 IFNs and attenuated signalling in RSV-infected cells. NS1 directly binds interferon regulatory factor 3 and prevents it binding to the IFN-β promoter region, while both NS1 and NS2 both increase STAT2 proteasome-mediated degradation leading to weakened IFN responses.55 56 Such attenuated immune responses have significant consequences for RSV disease (box 2).
RSV interference with the host immune response
Non-structural (NS) proteins
NS1 disrupts interferon regulatory factor 3 binding to the interferon-β promoter.
NS2 protein binds retinoic acid-inducible gene I, blocking innate signalling.
NS1/2 enhance degradation of STAT2, terminating innate response.
NS1/2 inhibits conventional dendritic cell (DC) maturation, inhibiting antigen-presenting cell functions.
Surface glycoproteins
Glycoprotein binds to CX3CR1 on plasmacytoid DC/cilliated cells.
Secreted glycoprotein acts as a decoy for antibody.
Fusion protein binds to toll-like receptor 4, possibly causing innate desensitisation.
Internal proteins
Nucleoprotein disrupts the synapse between CD4 and CD8 cells.
Host response
Pathogenesis
Wheezing, hypoxaemia and increased respiratory effort are a consequence of the intense inflammatory infiltrate recruited to the respiratory tract in response to RSV infection. The inflammatory cell infiltrate is associated with copious mucus production, oedema and shedding of AECs, all of which contribute to critical narrowing of the small airways. Previously, hypotheses about disease severity have related it to an inappropriately intense immune response; however, new evidence challenges this classical view. A recent study by Thwaites et al showed that severe RSV bronchiolitis in infants was associated with a paradoxically reduced viral load, IFN-γ, CCL5 and type I IFN gene expression but increased levels of interleukin (IL)-17a and mucin gene expression.57
Innate immune response
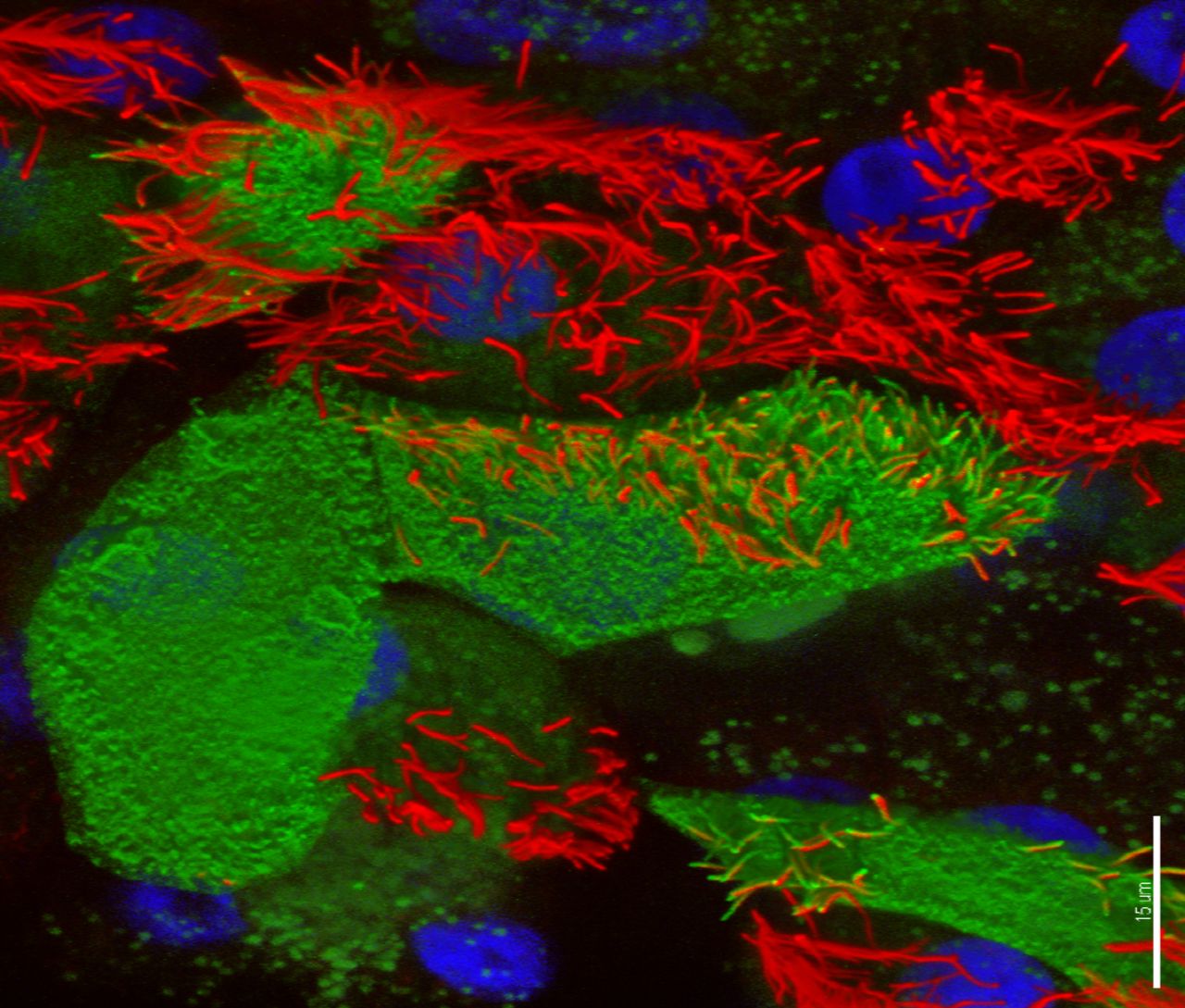
RSV primary infects ciliated AECs (figure 3).58 The epithelium is protected by a thick mucus layer containing mucin and sialic acid compounds; bronchial epithelial cells (and A549 human alveolar basal epithelial adenocarcinoma cells) specifically express the mucin MUC5AC.59 Also important are cathelicidin and other host defence peptides which impede RSV invasion by directly acting on the virus envelope. The airway epithelium produces chemokines including the CXC chemokines inducible protein (IP)-10 and IL-8 which recruit neutrophils to the airway lumen, in large numbers, shortly after infection.60 AECs are also an important early source of type 1 IFNs, which upregulate MHC-I and drive type 1 T helper cell (Th1) responses. RSV-NS1 and NS2 interact with AECs after cell invasion to block such IFN responses and mitigate virus clearance.61
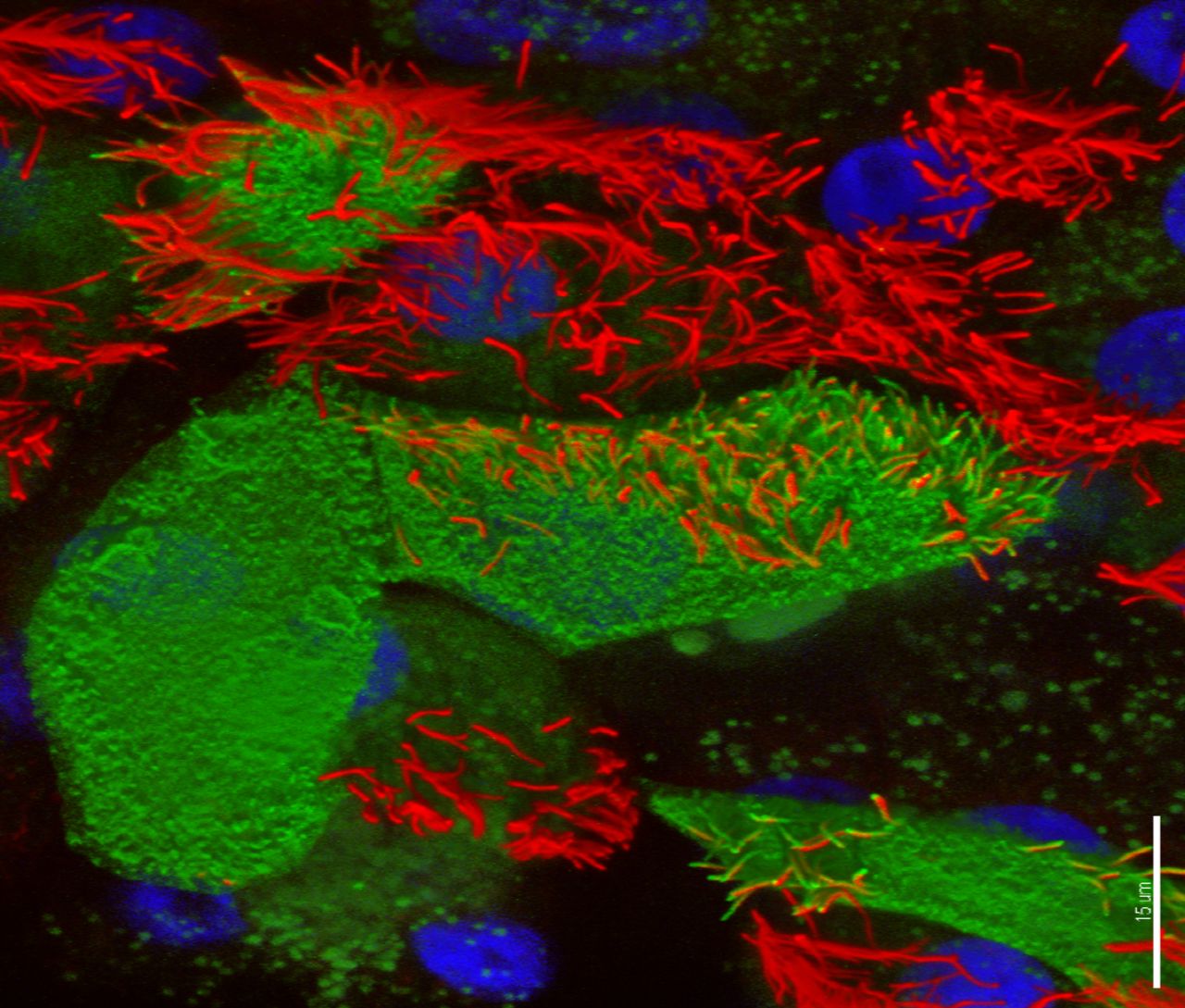
Confocal image of respiratory syncytial virus (RSV)-infected human nasal ciliated epithelial cells in vitro. At 72 hours postinfection, viral antigens (stained green, fluorescein isothiocyanate) are seen on the surface of the cell and the full length of some the cilial shafts. Antibodies against acetylated tubulin were used to detect the axonemal microtubules (AlexaFluor 594; red); nuclei are blue. Three-dimensional reconstruction of all channels was performed with Imaris (Bitplane AG, Zurich, Switzerland) blend filter. Scale bar 15 um. Image kindly provided by Dr Claire Smith, UCL GOS Institute of Child Health, University College London, UK.
Other aspects of innate immune responses include alveolar macrophages, DCs and innate lymphoid cells (ILCs), which release IFN-ϒ, IL-1β and IL-18, further promoting generalised inflammation.62 These proinflammatory cytokines lead to recruitment of conventional DCs, and later CD8+ T cells and Th1 CD4+ T cells as well as immunoglobulin (Ig)G and IgA producing and primed B cells.
The pDCs are another source of INF-α/β (type 1 IFNs) which elicit antiviral responses.63 Whereas adult pDCs have been shown to recognise intracellular virus RNA via Rig-I-like receptors and subsequently produce type 1 IFNs, this function in infant pDCs, and even up to 5 years of age, is greatly reduced.64
Neutrophils, recruited by the AEC-derived CXC chemokines, IP-10 and IL-8, and CCL2 and CCL4 are the most prevalent inflammatory cell in the airways of RSV-infected infants, but their role in host defence is not clear.60 Group 1 ILCs contain the cytotoxic NK cell which is an important source of IFN-ϒ and drives the initial antiviral responses in RSV infection. NK cells may even play a role in controlling immunopathology, as two studies have found that the number of NK cells in the lung and blood were inversely proportional to the severity of RSV infection.65 66
Polymorphisms of the innate immune system are associated with increased susceptibility to RSV infection, but mutations in IL-4-Rα and IL-8 are also implicated.67–69 Polymorphisms in MX1 (encoding the MX1 protein which blocks primary transcription of viral RNA) are associated with severe RSV infection,70 as is Marco (an innate immune system scavenger receptor); Marco−/− mice show enhanced neutrophil and monocyte recruitment during RSV infection.71 While many genes have minor effects on RSV disease, there is currently no evidence of a strong genetic component.
Adaptive immune response
High antibody titres of both IgA and IgG are associated with partial protection in adults and may prevent RSV-associated LRTI, but antibody levels wane rapidly after episodes of reinfection.72 After RSV exposure, both IgG and IgA responses are seen; such B cells are both T cell dependent and independent. In infants, follicular DC structures are not well developed, so B cell activation relies more on T cell-independent pathways. In addition, neonatal B cells have low expression of transmembrane activator and calcium modulator and cyclophilin ligand interactor (receptor for B cell activating factor (BAFF) and A PRoliferation-Inducing Ligand (APRIL)), CD86, CD80 and CD40, and lack the alpha chain of the IL-4 receptor and so respond poorly to stimulation.73–75
Extrafollicular B cell activation also leads to IgA+ and IgG+ memory B cell formation and this process relies on stimulation by protein ligands BAFF and APRIL.76 One study found that infant bronchial epithelial cells had high expression of BAFF during RSV infection, suggesting a role for airway epithelium in supporting lung B cell responses.76 Even with F-specific IgG present, adults may still be reinfected with RSV, highlighting a prominent role for IgA.77 78 Singleton et al found that nasal associated lymphoid tissue IgA+ B cells declined rapidly postinfection, but these B cells confer protective immunity.79 In human challenge trials, individuals resistant to RSV have significantly higher levels of nasal RSV-F-specific IgA.80
RSV prevention
Intravenous polyclonal immunoglobulin (given as a monthly infusion) is partially effective as prophylaxis, but this has now been replaced by palivizumab,81 a humanised mouse monoclonal antibody that neutralises RSV and is effective prophylactically. It is administered as a monthly intramuscular injection during the RSV season and binds F protein, preventing infection of both RSV A and B.82 It reduces hospitalisation due to RSV, but its limited efficacy and cost have restricted its indications to the most vulnerable infants.83 A new RSV-F monoclonal antibody (MEDI8897) with a longer half-life than palivizumab finished phase IIb trials in February 2019.84 Monoclonal antibodies against RSV-G (eg, 3A5 and 5H6) reduce viral load in mice but have not been used clinically.85 Antibody therapy is not of proven value therapeutically in infected persons.
RSV vaccines are not yet available but are expected to be of use not only in young children at risk, school-age children who act as disease vectors, adult caregivers and the elderly but also those with chronic cardiopulmonary conditions. In children, the most vulnerable are neonates to 6 months of age, with many infants therefore at risk before the routine immunisation schedule begins. One strategy would be to vaccinate pregnant mothers in the second or third trimester so that transplacental transfer of protective antibodies can take place and delay the median peak of RSV disease.86
Historically, formalin-inactivated RSV vaccines (FI-RSV) led to higher rates of admission during subsequent natural RSV infections, and at least two vaccinated infants died from vaccine-augmented disease.87 Enhanced pathology may in part be due to immune complex deposition in the lungs leading to complement activation causing enhanced immunopathology and bronchopneumonia.88 FI-RSV induced led to high levels of RSV-F antibodies but because these were not neutralising probably allowed virus-specific T cells (including Th2 cells) to cause an exuberant cellular immune reaction in the lung.89
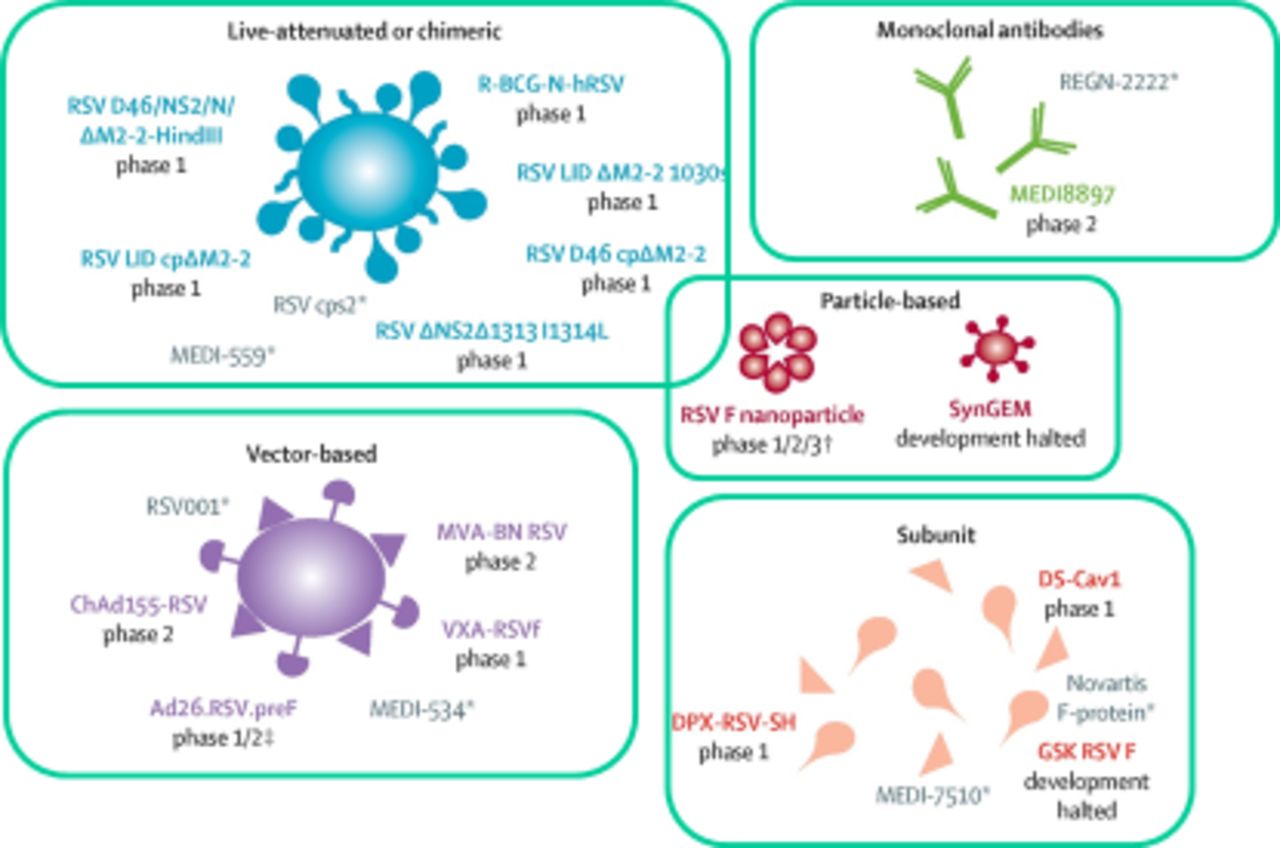
In order to induce neutralising antibody, many current vaccine programmes (figure 4, adapted from Mazur et al 90) are now focussed on the use of prefusion RSV-F or live attenuated vaccines that can be administered intranasally; these have the advantage of inducing mucosal antibody. At the time of writing, there are six live attenuated vaccines that are in phase I trials and four more in the preclinical stage of development.91 Live replicating agents are likely to be of most use in recipients under 2 years of age; replication being reduced in older populations with prior RSV exposure.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Respiratory syncytial virus (RSV) vaccine candidates and monoclonal antibodies in development.
There are currently five subunit vaccines in phase I trials and one in phase II trials clinical trial (four targeting preF, one targeting G and one targeting SH).91 Other vaccine candidates include particle-based and heterologous vector-based vaccines. Of the particle-based candidates, an RSV-F nanoparticle vaccine is currently in phase I, II and III trials; PREPARE phase III trial in pregnant women92; RESOLVE phase II trial in adults,93 showing some apparent benefit in patients with COPD and a third trial in infants which has just concluded phase I.91 94 A novel mucosal particle-based vaccine (SynGEM, Mucosis BV) induces RSV-F antibodies but requires additional optimisation (PMID: 30753101 DOI: 10.1164/rccm.201810-1921OC). Finally, there are currently four vector-based vaccines in phase I/II using a variety of RSV proteins91 (table 1).
Current vaccine pipeline as of December 2018
Maternal vaccination has considerable potential advantages, but the RSV-F nanoparticle approach (Novavax) has limited apparent efficacy; the GSK RSV-F (GSK) is a subunit vaccine and the RSV-F DS-Cav1 (National Institutes of Health/National Institute of Allergy and Infectious Diseases/Vaccine Research Center) is also a subunit vaccine.91 In February 2019, Novavax reported that their maternal vaccination approach did not meet primary objective of prevention of medically significant RSV LRTI in infants but showed partial efficacy against secondary objective (RSV LRTI hospitalisation) and reduced the frequency of severe disease among offspring.95 For elderly at-risk patients, a variety of particle based, vector based and subunit vaccines are in the pipeline; MVA-BN RSV is a notable vector-based candidate and DPX-RSV-protein is a subunit vaccine.91
Antiviral therapy
There are multiple antiviral therapies in development which centre around two main approaches: ‘entry as target’ and ‘non-entry as target’. Entry as target therapies block virus fusion via RSV-F and include palivizumab (a monoclonal antibody which binds RSV-F) and fusion inhibitors such as presatovir (allosteric inhibitors of RSV-F). Although effective in prevention, palivizumab has no therapeutic effect in acute RSV infection.96 Fusion inhibitors have gained the most attention with presatovir the most recent. Presatovir has undergone five phase II trials but has limited impact on viral load and had no effect on clinical features of RSV disease; clinical trials have been halted due to safety concerns.97 Other fusion inhibitors include MDT-637 (entered phase IIa trials but currently on hold), JNJ-2408068 (on hold due to safety concerns),98 TMC353121 (showed virus reduction during in vivo trials),99 BMS-433771 (reduced virus levels in mice studies when given prior to infection, suggesting use as a prophylactic agent),100 BTA-C585 (virus mRNA reduction in mice studies), P13 and C15 (awaiting results). Newer strategies include modifying the host rather than targeting the virus directly, with focus on RSV-binding molecules such as CX3CR1 (fractalkine) and nucleolin, though more work is required to assess safety and efficacy.101
‘Non-entry as target therapies’ interfere with the RSV polymerase which is responsible for viral RNA replication. The RSV polymerase is composed of an N-RNA template which alongside L and P proteins forms the polymerase itself, and this complex utilises M2-1 as a transcription elongation factor for efficient processing of RNA. ‘Non-entry as target therapies’ are separated into five different classes targeting different proteins of the polymerase: L-protein inhibitors (eg, ribavirin), N-protein targeting inhibitors (eg, RSV604), N–P protein–protein interaction targets and M2-1 protein targets.
ALS-008176 is an L-protein inhibitor, and clinical trials in adults showed both virus level reduction and symptom improvement,102 but clinical trials have been halted due to safety concerns. BI-D (another L-protein inhibitor) has shown virus reduction in mice studies but is yet to start human trials.103 RSV604 is an N-protein inhibitor but showed no reduction in viral load during human trials.104 Ribavirin (L-protein inhibitor) is currently the only approved antiviral agent, but its efficacy is largely disputed and has no clear impact on mortality105 and essentially is no longer used in the UK. Future antiviral agents could be used prophylactically in high-risk infants or adults as shown by the promising data on BMS-433771 fusion inhibitor.
Markers of severity
Studies are underway to identify new markers of severity, for example, by measuring gene expression profiles of CD4+ T cells. One study found that genes associated with neutrophil activation and inflammation were expressed more strongly in severe RSV disease; there seems to be a correlation between prolactin signalling and severity of infection, possibly explained by prolactin inhibiting Th1 responses which may contribute to pathology.17 Transcriptomic work also reveals an overexpression of SOCS genes which regulate T cell differentiation. SOCS2 and SOCS3 lead to Th2 and Th17 differentiation, and both, alongside SOCS5 are raised in severe RSV infection.17 106 107 Such new markers of disease severity may offer greater insight into which patients are likely to experience more severe disease and thus be candidates for more intensive management.
Conclusion
Despite decades of intensive research, much remains to be discovered regarding the host response to RSV infection. The virus’s apparent immunomodulatory adaptations have so far evaded the efforts of vaccinologists and the attempts of clinical scientists to specifically ameliorate its clinical manifestations. However, RSV is now square in the crosswires of many research teams and highly capable groups as one of the major remaining tractable challenges in infectious diseases, and there is renewed optimism that a new generation of vaccines and antivirals is on the near horizon. Clinical studies will reveal what part they have to play in prevention and treatment of disease, both in children and in vulnerable adults.
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.↵
- 91.↵
- 92.↵
- 93.↵
- 94.↵
- 95.↵
- 96.↵
- 97.↵
- 98.↵
- 99.↵
- 100.↵
- 101.↵
- 102.↵
- 103.↵
- 104.↵
- 105.↵
- 106.↵
- 107.↵
- 108.
Footnotes
Contributors All authors contributed to the conception, literature review, writing and revision of this manuscript.This work was supported by NIHR.
Funding This work was supported by NIHR.
Competing interests None declared.
Patient consent for publication Not required.
Provenance and peer review Commissioned; externally peer reviewed.