Article Text

Abstract
Introduction COPD is a chronic, progressive, inflammatory disease of the lungs and the third leading cause of death worldwide. The current knowledge of the pathophysiology of COPD is limited and novel insights in underlying disease mechanisms are urgently needed. Since there are clear parallels between ageing and COPD, we investigated genes underlying lung ageing in general and abnormal lung ageing in COPD.
Methods Whole genome mRNA profiling was performed on lung tissue samples (n=1197) and differential gene expression with increasing age was analysed using an adjusted linear regression model. Subsequent pathway analysis was performed using GeneNetwork and the gene-expression signature was compared with lung ageing in the Genotype-Tissue Expression (GTEx) project. In a subset of patients with COPD (n=311) and non-COPD controls (n=270), we performed an interaction analysis between age and COPD to identify genes differentially expressed with age in COPD compared with controls, followed by gene set enrichment pathway analysis.
Results We identified a strong gene-expression signature for lung ageing with 3509 differentially expressed genes, of which 33.5% were found nominal significant in the GTEx project. Interestingly, we found EDA2R as a strong candidate gene for lung ageing. The age*COPD interaction analysis revealed 69 genes significantly differentially expressed with age between COPD and controls.
Conclusions Our study indicates that processes related to lung development, cell-cell contacts, calcium signalling and immune responses are involved in lung ageing in general. Pathways related to extracellular matrix, mammalian target of rapamycin signalling, splicing of introns and exons and the ribosome complex are proposed to be involved in abnormal lung ageing in COPD.
- Copd ÀÜ Mechanisms
Statistics from Altmetric.com
Key messages
What is the key question?
Can we identify a lung tissue gene-expression signature that reflects lung ageing and abnormal lung ageing in COPD?
What is the bottom line?
We identified a strong gene-expression signature for lung ageing with EDA2R as a strong candidate gene for lung ageing and found that genes differentially expressed with increasing age between patients with COPD and non-COPD controls are involved in processes related to extracellular matrix organisation, mammalian target of rapamycin signalling, splicing of introns and exons, and the ribosome complex.
Why read on?
The data presented in this study enhances the current understanding of abnormal lung ageing in COPD by complementing the current data with genome-wide gene expression data specifically on lung tissue and provide several new candidate genes involved in lung ageing and abnormal lung ageing in COPD.
Introduction
COPD is a chronic and progressive, inflammatory lung disease, characterised by persistent airflow limitation.1 Although exposure to cigarette smoke is the main risk factor for COPD, its development also involves other environmental and occupational exposures.2 Patients with COPD suffer from severe respiratory symptoms, resulting in inability to work, invalidity and an overall worse quality of life. The prevalence of COPD is high, especially in the elderly, and will increase even further due to the ageing population, leading to a considerable economic and social burden.1 The current knowledge of the pathophysiology of COPD is limited and novel insights in the disease mechanisms are urgently needed.
A novel and interesting approach towards a better understanding of COPD is the accelerated lung ageing hypothesis.3 It has been postulated that acceleration of the normal ageing process is involved in the disease pathogenesis of several chronic degenerative diseases, including COPD.4 Ageing is defined by the progressive loss of physiological integrity, resulting in impaired function and increased risk of disease or death.3 5 While the normal ageing lung is characterised by a decline in lung function, loss of elasticity, alveolar enlargement and inflammation, the majority of these features are also observed in the lungs of patients with COPD.4 Moreover, several other markers of ageing such as telomere shortening, cellular senescence and stem cell exhaustion have shown to be increased in patients with COPD.6 Shortening of telomere length, as well as increased expression of the senescence marker p21 was demonstrated in circulating lymphocytes of patients with COPD.7–9 Furthermore, pulmonary fibroblasts of patients with COPD showed increased expression of senescence associated beta-galactosidase activity when compared with control fibroblasts.10 Thus, there are clear parallels between ageing and COPD and better understanding of the underlying mechanisms of lung ageing and accelerated lung ageing in COPD may provide highly warranted new insights into the pathogenesis of COPD.
Notwithstanding the fact that the evidence for a role of lung ageing in COPD is increasing, little is known about the genes underpinning lung ageing and accelerated lung ageing in COPD. Therefore, in this cross-sectional study we first investigated differences in gene expression associated with age in human lung tissue. Second, we generated a gene-expression lung tissue signature for accelerated lung ageing in subjects with COPD.
Material and methods
Lung tissue collection
Lung tissue was collected from patients undergoing lung resection, lung volume reduction or transplant surgery at one of the three participating sites in Groningen (The Netherlands), Québec (Canada) or Vancouver (Canada) in accordance with local ethical guidelines as previously described.11 12 If resection surgery was conducted for tumour removal, macroscopically normal lung tissue was taken far distant from the tumour. All samples were histologically checked for abnormalities using standard H&E staining.
For the gene-expression signature for lung ageing, we included a total of 1197 subjects from the database. The subjects were between 4 years and 85 years of age and subjects with unknown age were excluded as well as subjects with cystic fibrosis. For the age*COPD interaction analysis, a selection of 311 well defined current and ex-smoking (>5 pack-years) patients with COPD (FEV1/FVC ratio <70%) and 270 non-COPD controls (FEV1/FVC ratio >70%) was used.12
Gene expression arrays
Total RNA was extracted from lung tissue at Rosetta Inpharmatics Gene Expression Laboratory (Seattle, Washington, USA) for all three centres. mRNA profiling was performed using a custom-made Affymetrix HU133 array (GPL 10379) containing 751 controls probe sets and 51 627 non-control probe sets. Unfiltered gene expression was normalised with the Robust Multichip Average method with background subtraction implemented in the Affymetrix Power Tools software V.1.8.5.11 For the analyses, the log2(microarray intensity) values of the expression are used. The data are deposited at the Gene Expression Omnibus of the National Centre for Biotechnology Information with the accession number GSE23546.
Statistics
Linear regression analyses were performed using R software V.3.2.3.13 To adjust for strong expression differences between the samples due to unknown factors such as batch or technical variation, we used the common principal component approach.11 12 We calculated the principal components for each cohort separately and adjusted all our analyses for the principal components explaining more than 1% of the variation within each cohort. The potential confounders age, gender, smoking status and other diseases were excluded from the principal components calculation.
For the gene-expression signature for lung ageing, we additionally corrected for the potential confounders gender, smoking status (ie, never-smoking, current-smoking or ex-smoking) and disease if applicable (ie, COPD, non-COPD, unclassifiable COPD or non-COPD, alpha-1-antitrypsine deficiency, surgery for cancer, idiopathic pulmonary fibrosis, pulmonary hypertension or other diseases). For the age*COPD interaction analysis, we performed an age*COPD interaction analysis on the subset of the data set only including COPD and non-COPD controls on the three cohorts separately. This analysis was corrected for the potential confounders gender and smoking status (ie, current-smoking vs ex-smoking) and technical variance as described before. Equations for all analyses are written down in the online Supplement.
Supplementary file 1
For both of the analyses, we analysed the three cohorts separately and performed meta-analyses to identify genes that were differentially expressed with increasing age and behaved in the same direction across the three cohorts. To correct for multiple testing, the Benjamini-Hochberg false discovery rate (FDR) was applied.
Sensitivity analyses
The robustness of the gene-expression signature for lung ageing was tested in two subset analyses, that is, patients with COPD (n=311) and non-COPD controls (n=270). We compared the probe sets significantly associated with increasing age at nominal P value<0.05 in the same direction in the COPD and non-COPD control subsets with the 5732 probe sets significantly associated with increasing age in the whole data set.
To validate the gene-expression signature for lung ageing, we compared our signature with the lung ageing gene-expression signature identified in the Genotype-Tissue Expression (GTEx) project.14 Within this project, gene-expression signature in lung tissue was identified by modelling gene expression in a linear regression model using lung tissue of 119 subjects. From our differentially expressed genes (DEGs) with increasing age, ageing coefficients and P values of 3301 of these genes were available and the overlap in the same direction at nominal P value <0.05 was determined. In addition, we compared the associations between gene expression and age in our expression quantitative trait loci (eQTL) lung tissue database with adipose, artery, heart, lung, muscle, nerve, skin and thyroid tissues and blood from the GTEx for the top three most significant upregulated and downregulated genes with increasing age. Here, we used the age coefficient and the nominal P values for all the separate tissues as presented in the online supplement of the paper by Yang et al.14
The expression of EDA2R was confirmed in lung tissue from 21 ex-smoking non-COPD control subjects using quantitative PCR analysis.
GeneNetwork analysis
For the gene-expression signature for lung ageing, we used GeneNetwork, in which the biological processes are ranked with principal component analyses, to predict the enrichment of biological processes of the top 10% of the DEGs.15
Gene set enrichment analysis
For age*COPD interaction analysis we performed functional enrichment analysis with gene set enrichment analysis (GSEA) (V.7.0.670.1). Genes were ranked according to the strength of their t-statistics reflecting their interaction between age and COPD and subsequently analysed for enrichment in biological pathways listed in the Kyoto encyclopaedia of genes and genomes (KEGG).
Gene function prediction
To predict the function of the genes highlighted in the text, we used a combination of GeneNetwork and Genecards (www.genecards.org). GeneNetwork is built on an independent gene expression data set of 77 840 samples and predicts the function of genes based on known biological pathways available in the molecular signatures database MSigDB (http://www.broadinstitute.org).15
Results
Subject characteristics
An overview of demographical and clinical characteristics of the subjects included in this study are presented in table 1. A total of 1197 subjects were included in our analysis, with 357, 470 and 370 subjects in the Groningen, Quebec and Vancouver cohorts, respectively.
Subject characteristics
While there was a large range in age in all three cohorts, the median age of the cohorts was significantly different, possibly as a consequence of the lower age of the subjects in the Groningen cohort.
We did observe differences in smoking status and pack-years among the cohorts, with relatively more never smokers and lower number of pack-years in the Groningen cohort. Furthermore, the subjects from the Groningen cohort had a lower median lung function compared with the subjects in the cohorts from Quebec and Vancouver, and the above can be attributed to the fact that the Groningen cohort contained more subjects with very severe COPD undergoing lung transplantation. Based on the phenotypical differences between the cohorts described above, we analysed the cohorts separately, followed by a meta-analysis.
Gene-expression signature for lung ageing
Linear regression demonstrated a strong impact of age on lung tissue gene expression levels. Therefore, we restricted the FDR cut-off to <0.0001 and identified 5732 probe sets coding for 3509 significantly DEGs with age. With increasing age, 1980 genes were higher expressed, while 1529 genes were lower expressed with increasing age. Ectodysplasin A2 receptor (EDA2R), involved in the DNA damage response, was most significantly upregulated with age (FDR: 4.55×10-63), while microsomal glutathione S-transferase 1 (MGST1), involved in detoxification and the defence response to oxidative stress, was most significantly downregulated with age (FDR: 8.59×10-42). A complete list of these DEGs is provided in online supplementary table S1. A heatmap illustrating these results with the subjects ordered for each cohort separately based on their individual age is shown in figure 1A. To illustrate the expression pattern of the most significant DEGs with age, the individual gene expression of the top three genes most significantly upregulated (EDA2R, MAP4K1 and FRZB) and downregulated (MGST1, ZNF518B and ATP8A1) with age are shown in figure 1B–D and figure 1E–G, respectively.
Supplementary file 2

Differentially expressed genes upon increasing age in lung tissue. The heatmap shows probe sets coding for genes differentially expressed with increasing age at FDR<0.0001, for all the three cohorts from Groningen, Quebec and Vancouver. Each probe set is shown separately and for several genes more than one probe set was found to be significantly different. Relative increased gene expression is indicated in red, while relative decreased gene expression is indicated in blue. The top five genes upregulated and downregulated with increasing age are shown in the table next to the heatmap (1A). Log2 fold expression of the individual top three genes EDA2R, MAP4K1 and FRZB upregulated with increasing age (figure 1B–D). Log2 fold expression of the individual top three genes MGST1, ZNF518B and ATP8A1 downregulated with increasing age (figure 1E–G).
Next, we performed pathway analysis using GeneNetwork15 to identify enriched biological processes to get more insight in the relevance of the DEGs to the ageing individual. To this end, we selected the top 10% DEGs upregulated and downregulated with age. Genes upregulated with age were enriched for biological processes related to calcium signalling and immune responses, whereas genes downregulated with age were enriched for biological processes related to lung development and cell-cell contacts. An overview of the 25 most enriched biological processes for DEGs altered with age is shown in table 2, the complete list of all the enriched biological pathways is presented in online supplementary table S2.
Supplementary file 3
Top 25 enriched biological processes
Taken together, by the identification of 3509 DEGs with increasing age, we show a strong gene-expression signature for the ageing lung.
Sensitivity analysis of the lung ageing gene expression and validation of EDA2R
To assess if the gene-expression signature for lung ageing was robust, we performed a sensitivity analysis in the selected COPD and non-COPD control subjects. Of the 5732 probe sets significantly associated with increasing age in the whole data set, we found an overlap at nominal P-value <0.05 of 70% and 72%, respectively, in the COPD and non-COPD control subjects. A schematic overview of the comparison can be found in online supplementary figure S1a. Since the overlap is considerably higher than what can be expected by chance (2.5%), we are confident that our gene-expression signature of the ageing lung is robust.
Supplementary file 4
To further validate our gene-expression signature of the ageing lung, we determined the overlap with the lung ageing gene-expression signature identified in the GTEx project.14 From our signature, 3310 genes were present in the GTEx projects. By comparing these genes at nominal P value <0.05, we found an overlap of 33.5% in the same direction, which is 13 times more than what can be expected by chance. A schematic overview of the comparison can be found in online supplementary figure S1b.
Supplementary file 5
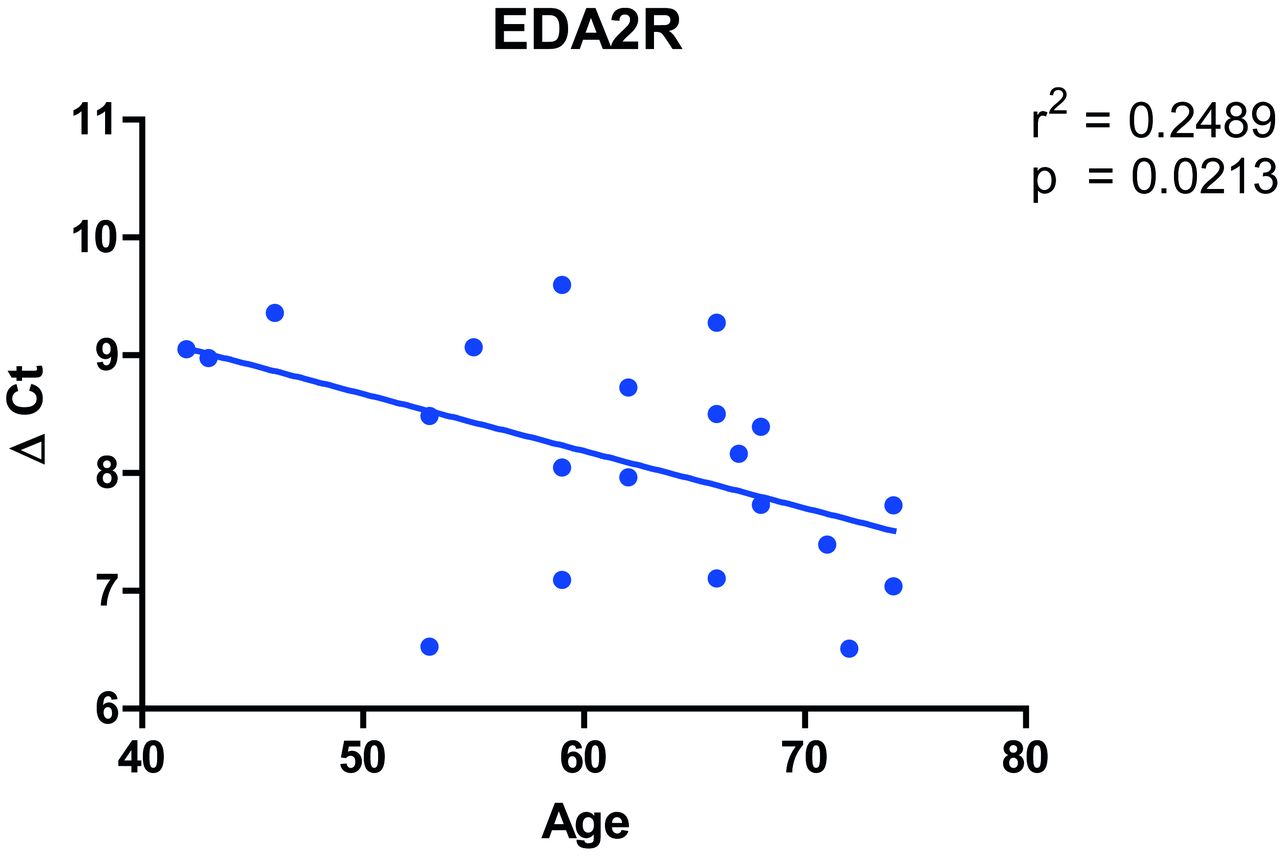
Of interest, the gene most upregulated with age in our analysis, EDA2R (figure 1B), was the second highest ranked gene among the genes upregulated with age in the GTEx project. Furthermore, in the sensitivity analyses in COPD and non-COPD subjects, EDA2R was the third highest ranked gene among the genes upregulated with age in both analyses. Moreover, quantitavie real time (qRT)-PCR validation of EDA2R in lung tissue of 21 ex-smoking non-COPD control subjects showed a significant association between increasing age and increased gene expression of EDA2R (figure 2). Based on these four analyses, EDA2R might be considered as a strong candidate gene for lung ageing in general.
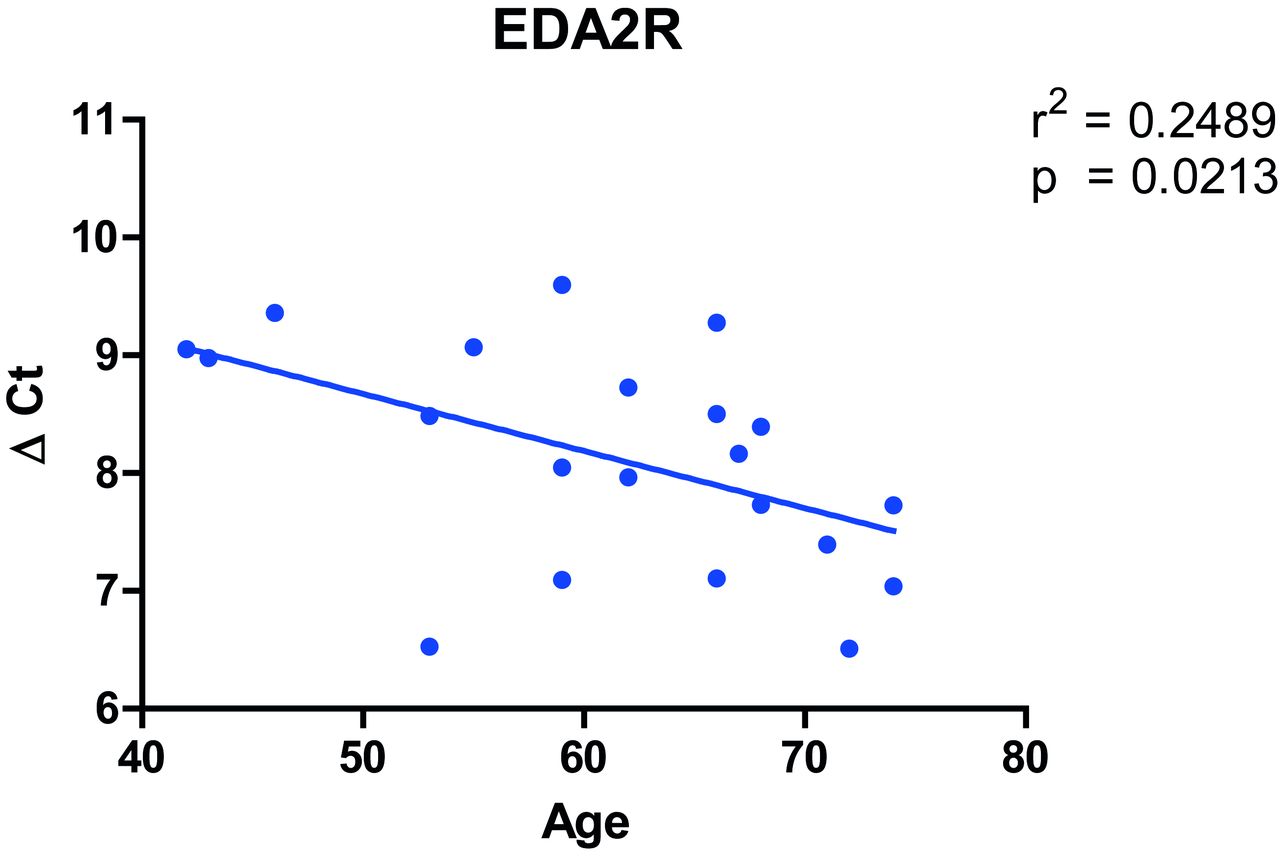
Association between gene expression of EDA2R and age in lung tissue. Age is displayed at the x axis and EDA2R gene expression relative to the housekeeping gene U6 ((ΔCt) on the y axis.
To further confirm our findings in the independent GTEx data set and to assess whether the associations between gene expression and age are specific for lung tissue, we compared the three most significant upregulated and downregulated genes with age from our lung tissue gene-expression signature with the available tissues within the GTEx project (online supplementary figure S2). The expression of the six genes was significantly associated with age in the same direction in lung tissue in the GTEx project. However, this association was not lung specific, since there were several significant associations in other tissues as well.
Genes differentially expressed with age in patients with COPD compared with non-COPD controls
While our previous analysis resulted in a gene-expression signature for lung ageing and identified EDA2R as a strong candidate gene for lung ageing, it did not discriminate between COPD and non-COPD. Since we hypothesise that COPD might be a disease of accelerated lung ageing, we were interested in genes whose expression changed differentially with age between individuals with and without COPD. To find these genes, we next assessed the interaction between age and COPD. An overview of demographical and clinical characteristics of the patients with COPD and non-COPD controls included in this subcohort are presented in online supplementary table S3 of the online supplement.
Supplementary file 6
Our age*COPD interaction analysis identified four DEGs with age in patients with COPD compared with non-COPD controls at an FDR cut-off of <0.05. Deoxyribonuclease I-like 3 (DNASE1L3), a gene involved in the complement activation cascade, and retinol binding protein 5 (RBP5), involved in the cellular binding of retinol, were the two DEGs that most significantly increased with age in COPD compared with non-COPD controls. KIAA1462, known to be involved in angiogenesis and blood vessel development, and G protein-coupled receptor 173 (GPR173), involved in tissue homoeostasis, were most significantly decreased with age in patients with COPD compared with non-COPD controls. When using a more lenient FDR cut-off of <0.25, we found 100 probe sets coding for 69 genes differentially expressed in association with age in patients with COPD compared with non-COPD controls (table 3).
Differentially expressed genes
We decided to use GSEA for the pathway analysis as this is based on the complete ranked gene list reflecting the interaction between age and COPD. GSEA analysis showed that genes that decreased more with age in COPD compared with control were significantly enriched in the extracellular matrix (ECM)-receptor interaction pathway (figure 3A). This indicates that the expression of ECM-related genes is more decreased with age in patients with COPD than in non-COPD controls. Of interest, the three collagen genes COL6A3, COL3A1 and COL4A1 were among the core enriched genes in this ECM-receptor interaction pathway (figure 3A). Genes that increased more with age in patients with COPD compared with non-COPD controls were significantly enriched for the mammalian target of rapamycin (mTOR) signalling pathway (figure 3B). In addition, genes involved in the splicing of introns and exons and genes that belong to the ribosome complex, including the core enriched genes FAU and RPL38, were also more increased with age in patients with COPD compared with non-COPD controls (figure 3C,D, respectively).

{kind=link}
![[SP4.jpg]](https://thorax.bmj.com/content/thoraxjnl/73/7/609/DC4/embed/inline-supplementary-material-4.jpg?download=true){kind=link}
![[SP5.jpg]](https://thorax.bmj.com/content/thoraxjnl/73/7/609/DC5/embed/inline-supplementary-material-5.jpg?download=true){kind=link}
{kind=link}
{kind=link}
Pathway enrichment among genes differentially expressed with age between patients with COPD and non-COPD controls. Gene set enrichment analysis shows enrichment of genes whose expression levels significantly change with age in patients with COPD compared with non-COPD controls. Genes are ranked from left to right based on their expression in the age*COPD interaction analysis in selected patients with COPD and non-COPD controls. The position of each vertical bar indicates the position of a gene whose expression is associated with the indicated pathway within the ranked list. The height of the bar represents the running gene enrichment analysis score. Log2 fold expression of genes differentially expressed with age between patients with COPD and non-COPD controls and core enriched in the subsequent pathway are presented right next to the indicated pathway. Patients with COPD are shown in red and non-COPD controls in blue. ECM receptor interaction pathway with the core enriched genes COL6A3, COL3A1 and COL4A1 was enriched among genes downregulated with age (A). mTOR signalling pathway (B), genes involved in the splicing of introns and exons (C) and genes belonging to the ribosome complex with the core enriched genes FAU and RPL38 (D) were enriched among genes upregulated with age. ECM, extracellular matrix; mTOR, mammalian target of rapamycin.
Discussion
In this study, we showed that age strongly affects gene expression in lung tissue. We identified a clear lung tissue gene-expression signature for the ageing lung with 3509 genes being differentially expressed. Genes upregulated with increasing age are enriched in pathways involved in calcium signalling and immune responses, while genes downregulated with increasing age are enriched in pathways involved in lung development and cell-cell contacts. Sensitivity analysis in a subset of patients with COPD and non-COPD controls and in the GTEx project data set demonstrates the robustness of our findings and points towards EDA2R as a strong candidate gene for lung ageing. Moreover, the association between EDA2R gene expression and age was validated with qRT-PCR in lung tissue. The final analysis in a subset of patients with COPD and non-COPD controls revealed potential roles for genes involved in ECM, mTOR signalling, splicing of introns and exons, and the ribosome complex in abnormal lung ageing in COPD.
Ageing has been a topic of research for decades and with the increasing prevalence of COPD in the elderly, the role of ageing in the pathogenesis of COPD has gained increasing interest.4 Recently, Lopez-Otin et al defined nine general hallmarks of ageing,5 including genomic instability, telomere attrition and cellular senescence. Several of these hallmarks have also been observed in COPD,7 9 10 supporting a role of abnormal ageing in COPD. However, most of these studies have been performed in blood leucocytes and data on actual lung tissue is limited. In addition, not much is known yet about the genes underpinning lung ageing in general and accelerated lung ageing in COPD. Therefore, our cross-sectional study on differential gene expression in lung tissue of patients associated with their age is a first attempt to overcome these knowledge gaps. We revealed that processes related to calcium signalling, immune responses, lung development and cell-cell contacts are common between the most significant DEGs in our analysis. The most abundant biological processes among the genes downregulated with age comprise development of the respiratory system and cell-cell contacts. The observations on developmental pathways are in line with the theory that ageing might be a result of developmental decay, stating that ageing occurs as a consequence of alterations in developmental pathways.16 For pathways related to cell-cell contacts, it has been shown that the permeability of the intestinal epithelium decreases with age in humans17 and decreased expression of tight junction molecules has been observed in aged rats.18 Interestingly, within our research group reduction of tight junction molecules has been demonstrated in epithelium of patients with COPD, linking this specific hallmark with COPD.19 The enriched biological processes among genes upregulated with age involve calcium signalling and immune response. Calcium is an important intracellular signal, functioning in many different cellular processes,20 however to our knowledge a role for calcium in COPD has not been described. Ageing, and in particular ageing in COPD, has been associated with immunosenescence (age-related decline in the function of the immune system) and inflammageing (immunosuppression and low-grade inflammation).21 22 Our results further support the contribution of immune regulatory processes to lung ageing by showing enrichment of these processes at gene-expression level. The sensitivity analyses showed the robustness of our lung ageing gene-expression signature, and revealed a considerably large overlap between our signature and the lung ageing gene-expression signature obtained in the GTEx project. The observation that EDA2R was highly significant in all the analyses and the validation of the association between expression of EDA2R and age in lung tissue suggest that EDA2R might function as a potential biomarker for lung ageing. In contrast to validation of the mRNA expression of EDA2R in lung tissue, it was not possible to reliably quantify EDA2R at the protein level due to weak and unspecific staining on both western blotting and immunohistochemistry. Evaluating the ageing signature of the other tissues from the GTEx project14 shows that EDA2R is also highly associated with ageing in adipose tissue, artery, heart, muscle and skin tissue. Therefore, EDA2R seems not to be specific for lung ageing. EDA2R belongs to the tumour necrosis factor receptor superfamily and has been associated with the Nuclear Factor Kappa B (NF-ĸB) and p53 signalling pathways.23 24 The function of EDA2R should be further explored before we can make any statement on EDA2R as a marker for (lung) ageing.
The second aim of our study was to identify key genes involved in abnormal lung ageing in COPD. Based on our GSEA analysis, we identified four significantly enriched KEGG pathways. The ECM receptor interaction pathway was the only significantly enriched pathway among genes downregulated with increasing age in COPD compared with controls. One of the main phenotypes of COPD is emphysema, in which breakdown of the alveolar walls causes decreased gas exchange in the alveoli.2 Loss of ECM, and in particular a lack of repair, plays an important role in emphysema. Our observation that the expression of genes involved in ECM is more decreased with age in patients with COPD compared with non-COPD controls is in line with emphysema development in COPD. Furthermore, three collagen genes COL6A3, COL3A1 and COL4A1 are core enriched in the ECM receptor interaction pathway. Interestingly, the expression of COL6A3 and COL3A1 has been shown to be decreased in aged lungs compared with adult lungs in an ageing mouse model.25 Since collagens belong to the main components of ECM,26 this further strengthens our observation that ECM modulation plays an important role in abnormal lung ageing in COPD. We found mTOR signalling, ribosome and spliceosome pathways significantly enriched among genes upregulated with age in patients with COPD compared with controls. mTOR is involved in cellular growth and metabolism and the mTOR complex has been established as a central, evolutionary conserved regulator of longevity.27 Moreover, it has been shown that inhibition of mTOR can reverse cellular senescence, one of the main hallmarks of ageing.28 The fact that one of the upstream activators of mTOR, phosphoinositide-3-kinase, is increased in peripheral blood mononuclear cells of patients with COPD29 30 gives interesting leads to further explore in COPD lungs. In contrast to the mTOR signalling pathway, not much is known yet about the specific contribution of the ribosome complex and splicing of introns and exons to COPD and ageing. While protein synthesis (ribosome) and the splicing of introns and exons (spliceosome) are indispensable biological processes, their exact role in accelerated lung ageing in COPD has to be further explored.
Notwithstanding the fact that we identified clear gene-expression signatures for lung ageing and accelerated lung ageing in COPD, we have to consider some limitations of our study. First of all, there are differences between the cohorts with respect to age, smoking status and lung function, which might be attributed to the higher number of patients with early onset, very severe COPD included in the Groningen cohort.31 Although we adjusted for intercohort differences by analysing the three cohorts separately followed by a meta-analysis, we cannot completely exclude some cohort-related skewing of the results. Second, the power of our age*COPD interaction analysis was limited as the analysis was performed merely on COPD and non-COPD control subjects for the three centres separately, followed by a meta-analysis. The power limitation is reflected by the relatively low number of significantly DEGs after correction for multiple testing. Nevertheless, our GSEA analysis on the age*COPD interaction takes the complete gene list into account and clearly identified enrichment of pathways relevant to ageing and COPD.
In conclusion, we identified a clear age-related lung tissue gene-expression signature and identified several genes for which a role can be envisaged in abnormal lung ageing in COPD. Based on these findings, we propose processes in control of lung development, cell-cell contacts, calcium signalling and the immune response as important processes in lung ageing in general. Furthermore, we suggest processes in control of ECM homoeostasis, mTOR signalling, splicing of introns and exons, and the ribosome complex to be involved in abnormal lung ageing in COPD.
References
Footnotes
MB and C-AB contributed equally.
Contributors MdV, AF, DSP, DDS, YB, DN, VG, WT, MvdB and C-AB were involved in the conception, design and/or supervision of research; MdV, AF, RRW and TVdJ analysed data; MdV, AF, DSP, WT, MvdB and C-AB interpreted results of analyses; MdV prepared figures and drafted the manuscript; AF, DSP, WT, MvdB and C-AB. critically reviewed and revised the manuscript; all authors read and approved the final version of the manuscript. The authors would like to thank Wierd Kooistra for his technical assistance with the EDA2RqRT-PCR validation.
Funding This work was supported by an UMCG Healthy Ageing Pilot grant. CAB is an active member of COST Action BM1201.
Competing interests None declared.
Ethics approval The ethics committees of the Institut Universitaire de Cardiologie et de Pneumologie de Québec, UBC-Providence Health Care Research Ethics Board and University Medical Center Groningen conform the Dutch national ethical and professional guidelines.
Provenance and peer review Not commissioned; externally peer reviewed.