Article Text

Abstract
Background As immune regulatory and effector cells, monocytes play an important role in the blood–extracorporeal circuit contact-related acute lung injury in patients undergoing cardiopulmonary bypass (CPB). However, circulating monocytes are phenotypically and functionally heterogeneous, so we characterised how immature monocytes affect acute lung injury induced by CPB.
Methods The identification and dynamic changes in monocyte subsets were monitored by flow cytometry in patients undergoing CPB and in a rat model of CPB. The differentiation and migration of monocyte subsets were explored by in vitro cultures and adoptive transfer in the CPB rat model.
Results We observed a dramatic increase of two monocyte subsets in the peripheral blood of patients undergoing CPB, involving tumour necrosis factor (TNF)-α-producing, mature intermediate CD14highCD16+ monocytes and a novel immature CD14lowCD16− subset. The immature CD14lowCD16− monocytes possessed limited ability for TNF-α production, and failed to suppress T-cell proliferation mediated by T-cell receptor signalling. However, these immature cells were highly proliferative and could differentiate into TNF-α producing, mature CD14highCD16+ monocytes. In the rat model of CPB, we further demonstrated that CPB induced migration of immature monocytes into the lungs, either from the bone marrow or from the spleen. Moreover, we confirmed the hypothesis that immature subsets could contribute to CPB-induced acute lung injury by giving rise to TNF-α producing descendants.
Conclusions The immature CD14lowCD16− monocytes might contribute to blood-circuit contact-induced acute lung injury by generating TNF-α-producing, mature monocytes. New strategies based on monocyte manipulation could be a promising therapeutic approach for minimising CPB-related lung injury.
- Macrophage Biology
- ARDS
- Innate Immunity
Statistics from Altmetric.com
Key messages
What is the key question?
How do different subsets of monocytes contribute to cardiopulmonary bypass induced acute lung injury or pulmonary dysfunction?
What is the bottom line?
Human monocytes are phenotypically and functionally heterogeneous, and play different roles in the inflammatory response.
Why read on?
We demonstrated that the immature CD14lowCD16− subset generates downstream inflammatory CD14highCD16+ descendants, which might provide novel targets for perioperative lung protection by manipulating monocyte subsets at different checkpoints.
Introduction
Cardiopulmonary bypass (CPB) has been widely used clinically to support temporarily total heart and/or lung function during cardiac surgery.1 ,2 Despite development of the techniques, CPB exposes the blood to the large synthetic, non-endothelial cell surface of the extracorporeal circuit. Such blood-circuit contact may trigger severe systemic and local inflammatory responses, which subsequently can cause a number of clinical complications.3 ,4 It is acknowledged that almost all patients undergoing CPB have different degrees of acute lung injury (ALI) or pulmonary dysfunction.4 ,5 About 2% of adult patients develop acute respiratory distress syndrome (ARDS) or even acute respiratory failure with a high risk of mortality (15–72%).6 ,7
Multiple factors have been demonstrated to be involved in CPB-induced ALI. Among these factors, monocytes and macrophages have drawn considerable attention due to their important roles in immune responses and ALI,8 ,9 because systemic inflammatory responses can trigger monocyte trafficking into injured organs.10 ,11 After being recruited to sites of tissue injury and inflammation, monocytes can regulate local immune responses by acting as a key source of proinflammatory cytokines and functioning as precursors for macrophages and dendritic cells.12 ,13
Recently, monocyte heterogeneity prompted us to investigate the pathogenesis of CPB-induced lung injury. Early studies demonstrated that human circulating monocytes are phenotypically and functionally heterogeneous, and can be subdivided into CD14highCD16− classic inflammatory monocytes and CD14+CD16+ resident subsets.12 ,14–17 Other studies reported that among CD14+CD16+ monocytes, a subset with a CD14highCD16+ phenotype (termed as intermediate monocytes) was more closely correlated with the pathogenesis of many inflammatory diseases, exhibited a high capacity for production of tumour necrosis factor (TNF)-α, and regulated immune responses.18 ,19 CD14lowCD16+ monocytes (named non-classical monocytes) are smaller and less granular, and express higher levels of the C-X-C chemokine receptor (CX3CR1),15 patrol the vascular endothelium, and are involved in the innate surveillance of local tissues.15 ,20
The nomenclature for monocyte subsets was mainly based on studies of circulating monocytes.21 ,22 Recently, the bone marrow and the spleen have been identified as two critical reservoirs of monocytes. A large fraction of immature monocytes in the reservoirs are immature cells with a CD14+/lowCD16− phenotype,23 which might be released into the peripheral blood under stress conditions. In the present study, we determined whether and how these immature monocytes affect CPB-induced ALI. We found that although blood-circuit contact could cause transient sequestration of monocytes, the immature CD14lowCD16− subset, and TNF-α-producing CD14highCD16+ monocytes, dramatically increased in the peripheral blood of patients with CPB. We further applied a rat model of CPB, and hypothesised that immature monocytes might migrate from their reservoirs into the lungs after the CPB, and contribute to CPB-induced ALI by generating mature TNF-α-secreting descendants.
Materials and methods
Patients
Fifty patients (24 men and 26 women, aged 37–71 years) undergoing cardiac surgery with CPB were enrolled at Beijing Anzhen Hospital, Capital Medical University. All human blood samples were collected after written informed consent was obtained, and the study conformed to the tenets of the Declaration of Helsinki. For the analyses of monocyte subset kinetic changes, blood samples from patients were collected into EDTA vacuum tubes after the onset of anaesthesia (pre), and on postoperative days 0, 1, 3, 5 and 7 after CPB. Detailed information of all patients, including primary diseases and indications, is described in the online supplementary materials.
Supplemental material
CPB model
Adult male Sprague–Dawley rats (350–400 g) were obtained from the Vital River Laboratories (Beijing, China). The rat CPB model was established using a dedicated small animal ventilator, extracorporeal circulation machine, and pipelines. The model is described in detail in the online supplementary material.
Flow cytometry analysis and cell sorting
Cells from peripheral blood, bone marrow, spleen, lung or bronchoalveolar lavage fluid (BALF) were stained with the indicated markers, and monocyte subsets were identified and detected by flow cytometry. Monocyte subsets from patients and rats were sorted by FACS Aria II (BD, Palo Alto, California, USA). Detailed information is described in the online supplementary materials.
Carboxyfluorescein-diacetate succinimidyl ester staining and cell proliferation assay
T-cell proliferation was determined by measuring changes in fluorescence using carboxyfluorescein-diacetate succinimidyl ester (CFSE) labelling, as described previously.24
TNF-α RNA and protein measurements
The TNF-α mRNA level of different monocyte subsets from patients or rats was determined using real-time PCR. The secretion of TNF-α of human or rat monocyte subsets was detected using an ELISA or Cytometric Bead Array (CBA). Detailed descriptions of these procedures are provided in the online supplementary material.
In vitro labelling of CD43low monocytes and adoptive transfer
Rat CD43low monocytes were sorted with flow cytometry (FACS Aria II), then labelled with dioctadecyl tetramethylindotricarbocyanine iodide (DiR; Life Technologies, Carlsbad, California, USA). The DiR+ monocytes were injected into healthy rats through the vena orbitalis posterior plexus 1 hour before CPB.
In vivo labelling of CD43high monocytes with fluorescent latex microspheres
Anaesthetised rats were intravenously injected with Flouresbrite Plain 0.5 μm microspheres (2.5% solids w/v; Polyscience, Niles, Illinois, USA) through the left femoral vein to label CD43high monocytes, referring to the methods for labelling mouse Ly6chigh subsets.25 After 24 hours, the CPB rat model was created.
Statistical analysis
Data are reported as mean±SEM of values obtained from at least three independent experiments. Significant Differences between healthy control and CPB models, or between CPB models and the splenectomy CPB models were compared using Student's t test. Analysis of variance (ANOVA) was applied to analyse the expression differences of surface and functional molecules, and TNF-α protein or RNA expression of different monocyte subsets. During the follow-up of patients and the rat model, because the subjects in the study were observed at different times, and to determine the contribution of individual subjects and times to the difference across cell subtype kinetic changes, the repeated measurement data analysis of variance methodology was used and was followed by post hoc Bonferroni analyses. If the data were not symmetric, the Greenhouse–Geisser correction was used. p<0.05 was considered statistically significant.
Results
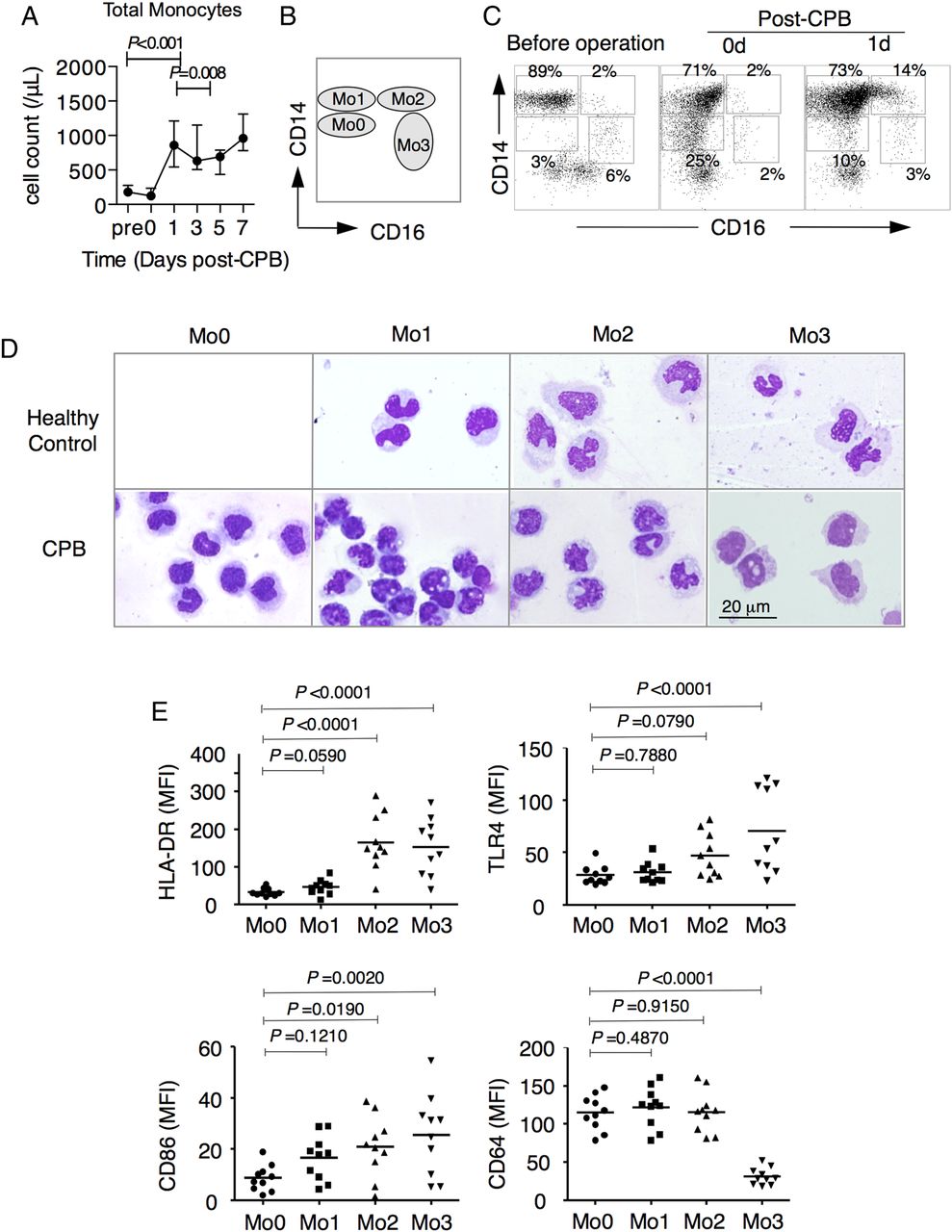
Emergence of CD14lowCD16− immature Mo0 monocytes after CPB
We found that the monocytes rapidly and dramatically increased within one day after CPB in patients undergoing CPB (figure 1A and online supplementary figure S1). To monitor the changes in monocyte subsets, blood monocytes were subdivided into three subsets: CD14highCD16− (Mo1), CD14highCD16+ (Mo2) and CD14lowCD16+ (Mo3), as previously described24 (figure 1B). Importantly, we noticed that a subset of monocytes with a CD14lowCD16− phenotype (named as Mo0, figure 1B,C) rapidly emerged in the peripheral blood of patients undergoing CPB (from 3% preoperatively to 25% at 0 days post CPB). Compared with Mo1 monocytes, Mo0 monocytes were more morphologically immature (figure 1D) and expressed similar levels of HLA-DR, toll-like receptor (TLR4), and CD64 as expressed by the Mo1 subsets, but lower levels of CD86 (figure 1E and online supplementary figure S2). However, compared with the Mo2 and Mo3 subsets, the Mo0 monocytes expressed lower levels of HLA-DR and CD86 (figure 1E and online supplementary figure S2).

Identification of human blood CD14lowCD16− immature monocytes induced by cardiopulmonary bypass (CPB). (A) Dynamic changes of total monocyte numbers. The peripheral blood from patients receiving CPB was collected at the indicated times, and total monocytes were analysed by flow cytometry combined with a Trucount Tube. The repeated measurement data analysis of variance (ANOVA) methodology was applied and was followed by post hoc Bonferroni analysis. Data are expressed as the median and IQR for each time point of 10 individuals and are representative of 10 individuals. (B and C) Ideograph (B) and representative flow cytometry (C) analyses of monocytes. The fresh peripheral blood from the patients was collected, stained with specific antibodies, and then lysed with red blood cell lysing buffer. According to the expression pattern of CD14 and CD16, human monocytes were divided into CD14lowCD16− (Mo0), CD14highCD16− (Mo1), CD14highCD16+ (Mo2), and CD14lowCD16+ (Mo3) subsets. (D) Morphology of peripheral monocyte subsets (Wright–Giemsa staining) from healthy controls (upper panel) and patients undergoing CPB (lower panel). Three or four monocyte subsets were sorted by FACS Aria II and Cytospin preparation of different monocytes for Wright–Giemsa staining. Magnification, 1000×, scale bar, 20 μm. (E) The mean fluorescence intensity (MFI) of HLA-DR, TLR-4, CD86 and CD64 in four monocyte subsets after 1 day from patients receiving CPB was analysed by flow cytometry. Mean values are indicated (n=10). ANOVA was applied to analyse the expression difference of surface and functional molecules of different monocyte subsets.
Immature Mo0 monocytes gave rise to mature, cytokine-secreting Mo2 monocytes
Given that Mo0 monocytes were immature, we tested whether the differentiation process was blocked. We found that about 10% of Mo0 monocytes from patients with CPB expressed nuclear protein Ki67, which was significantly higher than other subsets (figure 2A,B). This observation indicated that this population was more proliferative than other subsets. Next, we performed in vitro culture of whole blood samples from patients 1 day after CPB and pre CPB, and observed a rapid decrease in the percentage of Mo0 monocytes, along with a dramatically increased percentage of Mo2 monocytes within 16 h (figure 2C,D; online supplementary figure S3A,B). In addition to the above, in vitro differentiation results, we found a decreased percentage of Mo0 monocytes in the peripheral blood of patients at 1 day after cessation of CPB. Meanwhile, consistent with a previous report of patients with haemodialysis,26 we found a dramatic 8–10-fold increase (2.22%±0.23% vs 18.98%±2.24%; p<0.0001; figure 2E) in the number of Mo2 monocytes (figure 2E), which might reflect in vivo differentiation of Mo0 subsets into Mo2 monocytes. The kinetic change of percentage and numbers of different monocyte subsets for each patient was shown and the same trend was observed in all patients (see online supplementary figure S4A,B). The expression of HLA-DR on Mo0 and Mo1 monocytes decreased after CPB and gradually returned at 7 days post CPB (see online supplementary figure S5A,B).

Proliferation and differentiation of CD14lowCD16− immature monocytes. (A) Representative flow cytometry analyses combined with intracellular staining of Ki-67 expression in different monocyte subsets at 1 day after cardiopulmonary bypass (CPB). (B) The percentage of Ki-67-positive cells among Mo0, Mo1, Mo2 and Mo3 subsets from patients at 1 day post CPB. Analysis of variance (ANOVA) was used to analyse the differences among these groups (n=10). (C and D) The whole peripheral blood samples from patients at 1 day post CPB were cultured for indicated times using an in vitro differentiation assay. Representative scatter diagrams of flow cytometry (C) and kinetic changes of the percentages of different monocyte subsets (D) are shown from 10 individuals. The repeated measurement data ANOVA methodology was applied and was followed by post hoc Bonferroni analysis. Repeated measures ANOVA showed that the main effect of the percentage of different monocyte subsets on the kinetic change of monocyte subsets was significant (p<0.001). Post hoc comparisons indicated that the percentages of Mo0 and Mo1 significantly decreased (Mo0: 0–8 hours, p=0.014; 0–16 hours, p<0.001; 0–24 hours, p<0.001; 8–16 hours, p=0.007; for Mo1: 0–8 hours, p=0.028; 0–16 hours, p<0.001; 0–24 hours, p<0.001; 8–16 hours, p<0.001; and 8–24 hours, p<0.001), and the percentages of Mo2 gradually increased (0–8 hours, p<0.001; 0–16 hours, p<0.001; 0–24 hours, p<0.001; 8–16 hours, p<0.001; and 16–24 hours, p<0.001). Data are expressed as the mean±SEM of 10 individuals for each time point and are representative of 10 independent experiments. (E) The dynamic changes of percentage and numbers (n=10) of circulating monocyte subsets in the patients before CPB and post CPB at indicated times. The peripheral blood from patients receiving CPB was collected at the indicated times, and monocyte subsets were analysed by flow cytometry combined with a Trucount Tube. Data are expressed as the median and IQR of indicated individuals for each time point. The repeated measurement data ANOVA methodology was applied and followed by post hoc analysis. The repeated measures ANOVA did not find a significant effect of time on the kinetic changes of different monocytes (p=1.000). (F) Expression of tumour necrosis factor (TNF)-α protein by Mo0, Mo1 and Mo2 subsets was determined using Cytometric Bead Array (n=5). (G) Expression levels of TNF-α mRNA in monocyte subsets of patients were analysed with real-time PCR. Data are shown as mean±SEM of five individuals and are representative of five independent experiments.
Notably, CBA and RT-PCR analyses detected higher levels of TNF-α protein and mRNA in Mo2 monocytes (figure 2F,G), which suggested that immature Mo0 monocytes might contribute to CPB-induced inflammatory responses by generating mature, cytokine-secreting Mo2 monocytes.
CPB induced accumulation of immature and mature monocytes into the lungs
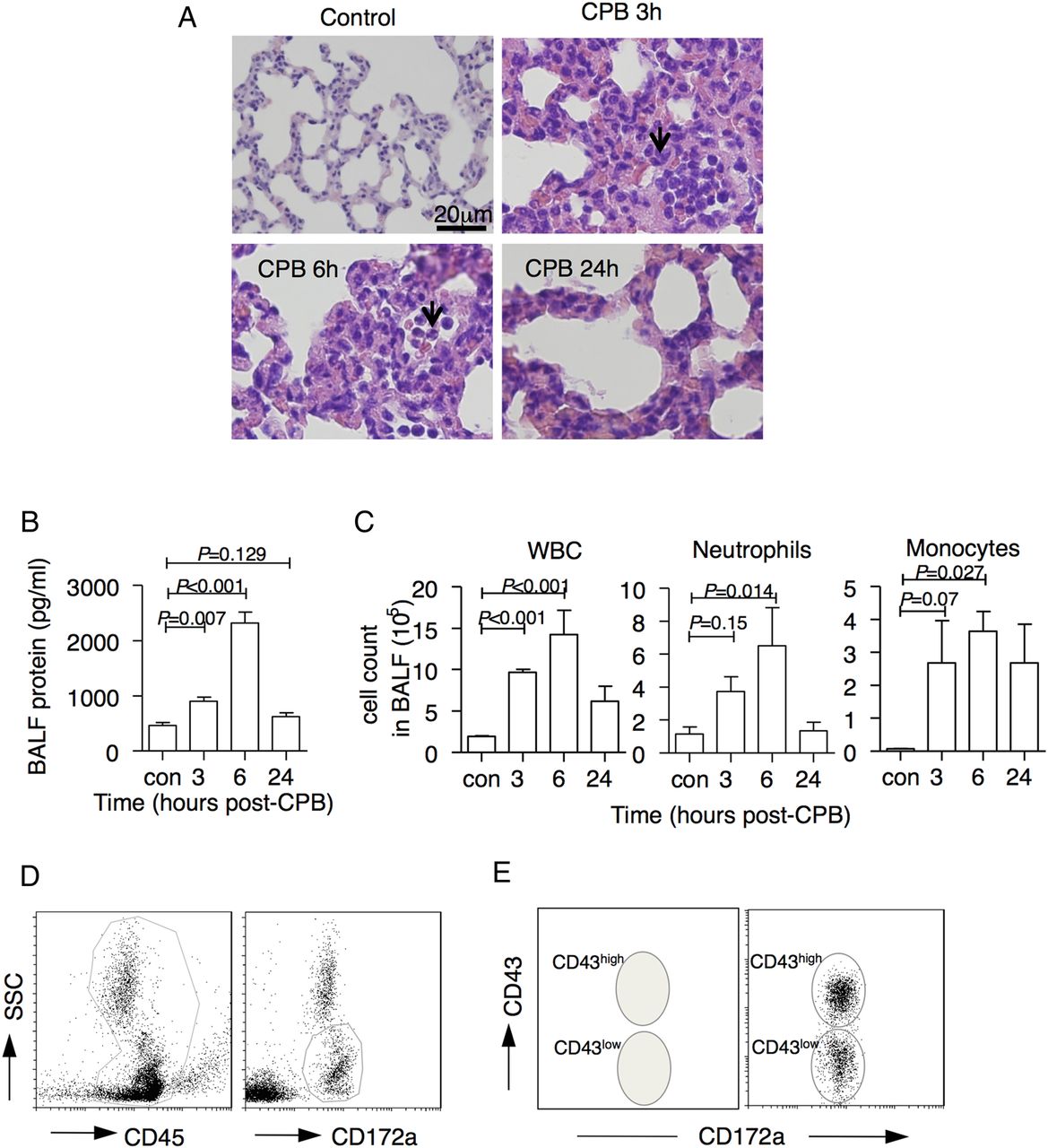
Because it is very difficult to investigate whether blood-circuit, contact-induced migration of immature monocytes occurred from their reservoirs into the lungs in patients with CPB, we used a rat CPB model. In this model, histological analyses showed that leukocytes were entrapped in the lung capillaries (figure 3A). We also observed an increase in total proteins and elevated cell counts of whole leukocytes, neutrophils and monocytes in BALF with a peak at 6 hours post CPB (figure 3B,C), which indicated that CPB caused ALI in these animals.


Cardiopulmonary bypass (CPB) induced acute lung injury in rats, and identification of rat monocyte subsets. (A) Representative histological image (H&E staining) of lung sections from the control group, and the 6 hours and 24 hours post-CPB groups. Magnification, 400×, bar, 20 μm. (B) Total protein concentration in bronchoalveolar lavage fluid (BALF) from rats with CPB was analysed using the Pierce BCA Protein Assay Kit (n=3 at the indicated times). (C) White blood cell (WBC), neutrophils and monocytes count in BALF at indicated times from rats with CPB were analysed by flow cytometry combined with haematometry (n=3 at the indicated points). (D and E) Representative flow cytometric profiles of rat monocyte subsets in the peripheral blood as defined by the expression of CD172a and CD43. Rat CD172a+ monocytes were divided into CD43low and CD43high monocytes. (F) Cytospin preparations of sorted CD43low and CD43high monocytes from the bone marrow, the peripheral blood, the spleen, and the lungs of healthy rats after Wright–Giemsa staining. Magnification, 1000×, scale bar, 20 μm. (G) Distribution percentages of CD43low and CD43high monocytes in bone marrow, peripheral blood, spleen, lung tissues, and BALF of normal rats (n=5). (H) A total of 2×104 CD43low and CD43high monocytes in the peripheral blood, lung tissues, and alveolar lavage fluid from model rats at 3 hours post CPB were sorted, cultured with lipopolysaccharide (1 pg/mL) in medium (1640 RPMI with 10% FBS) for 3 hours, and the supernatant was collected and analysed using a rat ELISA kit (n=5). (I) Expression levels of tumour necrosis factor (TNF)-α mRNA in sorted CD43low and CD43high monocytes in the peripheral blood, lung tissues, and alveolar lavage fluid from control rats or model rats at 3 hours post CPB were measured by real-time PCR (n=3).
According to previous studies,27 ,28 rat monocytes were subdivided into immature CD172a+CD43low (CD43low) and relatively mature CD172a+CD43high (CD43high) subsets (figure 3D–F, Wright–Giemsa staining). In healthy rats, immature CD43low monocytes accounted for ∼90% of monocytes in the bone marrow, while monocytes in the peripheral blood, spleen, lungs and BALF contained higher frequencies of CD43high monocytes (figure 3G). ELISA and real-time PCR analysis detected higher levels of TNF-α protein and mRNA in mature CD43high monocytes from peripheral blood and the lungs of CPB rats (figure 3H,I), which suggested that mature CD43high monocytes, rather than immature CD43low monocytes, might be a major source of TNF-α in response to blood circuit contact.
Consistent with the results from patients undergoing CPB, the frequency of the immature CD43low subset dramatically increased after the CPB operation and gradually returned to normal levels after 24 hours (figure 4A). Furthermore, we observed a rapid and transient increase in immature CD43low monocytes in the lung tissues, with a peak at 3 hours post CPB (figure 4B and online supplementary figure S6). In BALF, the absolute numbers of CD43high monocytes increased and reached a plateau at 3 hours after ceasing CPB (figure 4B). Thus, immature and mature monocytes might migrate into the lungs in response to CPB challenge.

Dynamic post-cardiopulmonary bypass (CPB) changes of monocyte subsets in rats. (A) Dynamic changes of the percentages of CD43low and CD43high subsets in the peripheral blood before (pre: control rat) and post CPB at indicated times. (B) Dynamic changes of absolute cell counts of monocyte subsets in lung tissue and bronchoalveolar lavage fluid (BALF) at the indicated times (n=3). (C) Dynamic changes of absolute cell counts of monocyte subsets in bone marrow (BM) and spleen at the indicated times (n=3). (D) Cell counts of CD43low and CD43high subsets in the lung and BALF at 3 hours and/or 6 hours post CPB in control rats, control sham rats, and CPB-treated rats with or without splenectomy (n=3). Data are expressed as the mean±SEM of three animals and are representative of three independent experiments. (E) Alveolar protein concentration in the lungs of rats with or without splenectomy at 6 hours post CPB was analysed by the BCA protein assay kit (n=3). (F) Respective H&E staining of the lungs of the rats with or without splenectomy at 6 hours post CPB (n=3). Magnification, 400×; scale bar, 20 μm.
Migration of CD43low and CD43high monocytes from the bone marrow and spleen in rats with CPB
It has been reported that in addition to the bone marrow, the spleen is also a reservoir for monocytes,11 ,29–31 Within 3–6 hours after ceasing CPB, the numbers of CD43low monocytes declined in the bone marrow and spleen to 50–60% of the baseline levels (figure 4C), which further suggests that immature monocytes were mobilised and migrated from bone marrow and spleen. By contrast, a significant decrease in CD43high monocytes was only observed in the spleen, and not in the bone marrow (figure 4C), which indicated that the majority of mature monocytes mobilised into the lungs were from the spleen.
To further confirm the notion, we performed CPB on rats with splenectomy. The number of CD43low monocytes in the lung tissues decreased by ∼20%, while the number of CD43high monocytes in BALF decreased more than 70% (figure 4D). There was no significant difference in monocyte subsets between the sham-operated control rats and healthy control rats (figure 4D). Thus, bone marrow might be the major source of CD43low monocytes, while the spleen might be involved in releasing CD43low and CD43high subsets, but predominantly CD43high. Consistent with a decreased number of monocytes in the lungs, ALI was alleviated in rats with splenectomy, as shown by a decreased protein concentration of BALF and less leukocyte infiltration into lung tissues (figure 4E,F).
CD43low monocytes migrated and contributed to CD43high monocytes in the lungs
Similar to human Mo0 monocytes, we found that rat CD43low monocytes could differentiate into CD43high monocytes in whole blood culture (figure 5A). Since CD43low monocytes can migrate into the lungs and rapidly differentiate, we wondered whether the immature monocyte could differentiate in vivo and enhance the number of CD43high monocytes in the lungs. To test this possibility, we sorted CD43low monocytes from normal rats, labelled them with DiR, and adoptively transferred them into recipient rats (described in figure 5B). At 6 hours after ceasing CPB, these recipient rats showed an increase in the percentage and number of DiR+CD43low monocytes in the lungs (figure 5C,D). Later, the number of DiR+CD43low monocytes started to decline, along with a slight increase in the number of DiR+CD43high monocytes in the lungs. Subsequently, we observed a dramatic increase in the DiR+CD68+ macrophages in the lungs and BALF at 18 hours after cessation of CPB (figure 5D). These results indicated that immature monocyte subsets could further differentiate into mature subsets and increase the number of CD43high monocytes in the lungs after a CPB challenge.

CD43low monocytes give rise to CD43high subsets. (A) Whole peripheral blood was collected and cultured using the in vitro differentiation assay for the indicated times, and the percentages of CD43low and CD43high monocyte subsets were determined by flow cytometry. (B) Experimental schedule for adoptive transfer of DiR-labelled CD43low monocytes. CD43low monocytes were sorted by FACS Aria II from bone marrow (BM) of control rats labelled with DiR, and transferred into control rats through the left femoral vein. After 1 hour, the cardiopulmonary bypass (CPB) model was used. (C and D) The percentages and cell counts of DiR+ CD43low and CD43high monocytes or macrophages in the lungs and the bronchoalveolar lavage fluid (BALF) were analysed by flow cytometry combined with haematometry at the indicated time points. Data are expressed as the mean±SEM of three individuals and are representative of three independent experiments.
Influx of CD43high subsets into the lungs
Next, we tried to directly traffic the migration of CD43high monocytes. However, we could not harvest sufficient CD43high monocytes from the spleen for adoptive transfer. Since fluorescent LX microspheres have been used to track selectively the migration of mouse Gr1low monocytes (equivalent to rat CD43high monocytes) from the peripheral blood to the injured tissues,25 we selectively labelled rat CD43high monocytes by intravenous injection of fluorescent LX microspheres. In the preliminary experiments, we found that all LX+ cells were in the CD43high subset at 24 hours after injection and could be observed in the blood and lungs for >7 days (figure 6A and online supplementary figure S7). Thus, the rats underwent CPB (outlined in figure 6B) at 24 hours after injection. Within 3 hours post CPB, we found that the number of LX+CD43high monocytes sharply decreased in peripheral blood, but increased in the lung tissues, which suggested that LX+CD43high monocytes migrate from the blood into the lungs (figure 6C). Furthermore, we observed an increase in LX+CD43high monocytes as well as LX+CD68+ macrophages in BALF at 24 hours post CPB (see online supplementary figure S8), which suggested that CD43high monocytes might have moved into the alveolar cavity and differentiated into macrophages. When we performed the same procedures with the rats with splenectomy, the influx of LX+CD43high monocytes decreased by ∼50% in the lungs and about fivefold in the BALF (figure 6D). This finding further supported the notion that splenic CD43high monocytes migrated into the lung tissues after CPB.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
CD43high monocytes migrated into the lung post cardiopulmonary bypass (CPB). (A) The mature CD43high monocytes were selectively labelled by fluorescent latex microspheres. Representative flow cytometric histogram of labelling rat CD43high monocytes. To label CD43high monocytes, anaesthetised rats were intravenously injected with Flouresbrite plain 0.5 μm microspheres (2.5% solids (wt/vol)) through the left femoral vein. After 24 hours, the CPB rat model was used. About half the monocytes were the CD43high subset at 3 hours after the latex injection, and more than 95% latex-positive cells in the blood and lung were the CD43high subset at 24 hours after injection. (B) The injection schedule for labelling of the rat CD43high monocyte subset in circulation is shown. (C) Numbers of latex-positive CD43low and latex-positive CD43high monocytes in the peripheral blood, the lung tissues, and the bronchoalveolar lavage fluid (BALF) of rats with CPB at different points were analysed by flow cytometry combined with haematometry (n=3 at each time point). (D) The number of latex-positive monocytes in the lungs and the BALF at 3 hours post CPB in rats with or without splenectomy. Data are expressed as the mean±SEM of three individuals and are representative of three independent experiments.
Discussion
Although a number of technological innovations and pharmaceutical interventions have led to progress in perioperative lung protection, CPB-induced ALI or ARDS is still considered to be a serious complication.7 In the present study, we characterised the distinct roles of monocyte subsets in CPB-induced ALI. We reported a dramatic and selective increase of two monocyte subsets in the peripheral blood of patients undergoing CPB involving the mature inflammatory CD14highCD16+ Mo2 subset that functions as a major source of inflammatory cytokine TNF-α, and the immature CD14lowCD16− Mo0 subset that contributes to the generation of downstream inflammatory subsets. Using a rat model of CPB-induced ALI, we found that inflammatory monocytes and immature monocytes could migrate into the lung from the bone marrow and spleen. More importantly, immature CD14lowCD16− monocytes contributed to ALI by generating their TNF-producing mature descendants.
Higher levels of TNF-α in BALF have been considered as a possible predictor for pulmonary dysfunction after cardiac surgery of patients undergoing CPB.32 In the present study, we showed that the human CD14highCD16+ subset and rat CD43high monocytes expressed higher TNF-α mRNA than other monocyte subsets after CPB, which indicated that inflammatory monocytes might be an important source of TNF-α in CPB-induced aseptic inflammatory responses. The importance of CD14+CD16+ monocytes in inflammation and tissue injury is also well documented in reports of laboratory and clinical studies.12 ,18 ,33 ,34 These cells are considered as inflammatory monocytes, because they express a higher level of HLA-DR, have higher ability of phagocytosis, produce more proinflammatory cytokines and chemokines such as TNF-α, interleukin 6, interferon γ, macrophage inflammatory protein 1α and 1β,19 ,34 ,35 and most importantly, are expanded in many inflammatory disorders. Mouse blood and splenic Ly6clow monocytes, the counterpart of human CD14+CD16+ monocytes, express higher TNF-α than Ly6chigh monocytes.30
Compared with circulating CD14+CD16+ monocytes, the CD14lowCD16− Mo0 subset usually resides in the bone marrow and spleen.23 The increase of this immature subset in the circulation indicates blood-circuit contact-enhanced emigration of immature monocytes from their reservoirs. Our study further increases our knowledge of monocyte subsets, especially during disease conditions. The Mo0 subset differed from the classical Mo1 subset. The Mo0 subset was morphologically more immature, had a lower level of CD14 and CD86, had a higher proliferative ability, and had a lower capacity of TNF-α production. In a previous study, a fraction of CD14+HLA-DRlow/− immature monocytes with immunosuppressive capacity were identified as monocytic myeloid-derived suppressor cells (M-MDSCs).36 Despite sharing a similar phenotype with monocytic MDSCs that are found in patients with leukaemia or cancer,36 ,37 Mo0 monocytes from patients undergoing CPB promoted rather than inhibited T-cell receptor (TCR)-induced T-cell proliferation (see online supplementary figure S9), which indicated that Mo0 monocytes were distinct from M-MDSCs and might still be capable of facilitating adaptive responses. Notably, Mo0 monocytes possess limited ability for TNF-α production, thus they do not seem to contribute directly to inflammatory cytokine-induced tissue damage.
However, we found that these precursor cells could differentiate into mature TNF-α-producing monocytes in response to aseptic inflammatory stimuli. This differentiation process from Mo0 to Mo2 monocytes could occur in the peripheral blood, as indicated by in vitro culture of whole blood samples from patients; or in situ in the lungs after being recruited into the lungs, as supported by in vivo adoptive transfer of DiR+CD43low monocytes in the rat CPB model. The latter finding is consistent with the recent finding that murine Ly-6C+/high monocytes, the counterparts to human CD14+CD16− Mo1 monocytes, could locally differentiate into Ly-6C−/low monocytes, macrophages or inflammatory DCs after being recruited into the site of inflammation.12 Moreover, unlike circulating monocytes from healthy humans that usually terminate the cell cycle, Mo0 monocytes from patients undergoing CPB are highly proliferating cells, as indicated by the higher level of Ki-67. Given that Mo0 and Mo1 monocytes possess differentiation capacity, the excessive proliferation might enable these cells to generate a higher number of mature inflammatory monocytes, which subsequently cause more severe tissue injury.
The differentiation of immature monocyte subsets leads to a very important consequence involving the enhanced accumulation of TNF-α-secreting mature monocytes in the lungs and BALF. The local immune responses in the tissues could be regulated by the recruitment of leukocytes.11 ,38 The first step of monocyte migration is to migrate from their reservoirs in the bone marrow and spleen.11 ,30 CPB induces the release of monocytes from bone marrow, which then migrate into the lungs in the monkey model.39 In rats undergoing CPB, bone marrow exhibited a rapid and dramatic decrease of immature CD43low monocytes, while the spleen showed a decrease in immature CD43low and mature CD43high monocytes. However, since splenectomy resulted in a more apparent decrease in CD43high than CD43low subsets, the spleen might be the main source of mature CD43high monocytes, while the majority of CD43low monocyte influx into the lungs is from the bone marrow.
Because of frequently complicated clinical situations, it is challenging to collect large amounts of samples from a single patient. We were still able to perform studies with samples from an adequate number of patients and obtain statistically meaningful data. However, samples from each patient were not sufficient to complete all studies. Thus, samples used between different experiments were not necessarily from the same group of patients. For the same reason, our study was limited by the small sample size, which may compromise our data interpretation due to less power in statistical analyses. Nevertheless, samples selected for all the studies were randomised to minimise potentially confounding factors. Importantly, data from our studies on the CPB rat model were consistent with the major findings from clinical samples.
Our study could provide possible novel targets for perioperative lung protection by manipulating monocyte subsets at different checkpoints, including exit from the reservoir, migration into the tissues, and differentiation into inflammatory subsets. Recently, this notion has been supported by the finding that angiotensin-converting enzyme inhibition (enalapril) could be beneficial in the treatment of myocardial infarction by preventing the release of monocytes from the splenic reservoir.40 More importantly, we also found that severe sepsis also results in similar changes to circulating monocytes in patients with severe sepsis and septic shock (Han et al, unpublished data). Thus, new strategies based on monocyte manipulation might also be a promising therapeutic approach for a wider range of inflammatory diseases.
References
Footnotes
ZX, JH, and XH contributed equally to this paper.
Contributors Study concept and design: XH and HZ; acquisition of data: ZX, JH, XH, JW, CJ, YH, XW, LS, XD and TL; analysis and interpretation of data: ZX, JH, XH, HW, GL and JZ; first drafting of the manuscript: ZX, JH and XH; critical revision of the manuscript for important intellectual content: all authors; study supervision: XH and HZ; data access and responsibility: XH and HZ had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis and for the submission.
Funding The authors received support from the National Natural Science Foundation of China (grant Nos. 81071587 to ZH, 81070203, 81270327 to HXT).
Competing interests None declared.
Patient consent Obtained.
Ethics approval All human blood samples were collected with written informed consent, and the study conformed to the tenets of the Declaration of Helsinki.
Provenance and peer review Not commissioned; externally peer reviewed.