Article Text

Abstract
Rationale Primary ciliary dyskinesia (PCD) is an autosomal recessive, genetically heterogeneous disorder characterised by oto-sino-pulmonary disease and situs abnormalities (Kartagener syndrome) due to abnormal structure and/or function of cilia. Most patients currently recognised to have PCD have ultrastructural defects of cilia; however, some patients have clinical manifestations of PCD and low levels of nasal nitric oxide, but normal ultrastructure, including a few patients with biallelic mutations in dynein axonemal heavy chain 11 (DNAH11).
Objectives To test further for mutant DNAH11 as a cause of PCD, DNAH11 was sequenced in patients with a PCD clinical phenotype, but no known genetic aetiology.
Methods 82 exons and intron/exon junctions in DNAH11 were sequenced in 163 unrelated patients with a clinical phenotype of PCD, including those with normal ciliary ultrastructure (n=58), defects in outer and/or inner dynein arms (n=76), radial spoke/central pair defects (n=6), and 23 without definitive ultrastructural results, but who had situs inversus (n=17), or bronchiectasis and/or low nasal nitric oxide (n=6). Additionally, DNAH11 was sequenced in 13 subjects with isolated situs abnormalities to see if mutant DNAH11 could cause situs defects without respiratory disease.
Results Of the 58 unrelated patients with PCD with normal ultrastructure, 13 (22%) had two (biallelic) mutations in DNAH11; and two patients without ultrastructural analysis had biallelic mutations. All mutations were novel and private. None of the patients with dynein arm or radial spoke/central pair defects, or isolated situs abnormalities, had mutations in DNAH11. Of the 35 identified mutant alleles, 24 (69%) were nonsense, insertion/deletion or loss-of-function splice-site mutations.
Conclusions Mutations in DNAH11 are a common cause of PCD in patients without ciliary ultrastructural defects; thus, genetic analysis can be used to ascertain the diagnosis of PCD in this challenging group of patients.
- Cilia
- dynein
- Kartagener syndrome
- dextrocardia
- heterotaxy
- airway epithelium
- atypical mycobacterial infection
- bacterial infection
- cystic fibrosis
- infection control
- nebuliser therapy
- rare lung diseases
- respiratory infection
- bronchiectasis
- paediatric lung disaese
- histology/cytology
- tobacco and the lung
- paediatric physician
- respiratory measurement
- asthma
- asthma epidemiology
- clinical epidemiology
- paediatric asthma
- lung proteases
- asthma genetics
- lung physiology
Statistics from Altmetric.com
- Cilia
- dynein
- Kartagener syndrome
- dextrocardia
- heterotaxy
- airway epithelium
- atypical mycobacterial infection
- bacterial infection
- cystic fibrosis
- infection control
- nebuliser therapy
- rare lung diseases
- respiratory infection
- bronchiectasis
- paediatric lung disaese
- histology/cytology
- tobacco and the lung
- paediatric physician
- respiratory measurement
- asthma
- asthma epidemiology
- clinical epidemiology
- paediatric asthma
- lung proteases
- asthma genetics
- lung physiology
Key messages
What is the key question?
Dynein axonemal heavy chain 11 (DNAH11) is known to be a primary ciliary dyskinesia (PCD)-causing gene in very few families, but data regarding the mutation prevalence are lacking.
This study details the mutation profiling of DNAH11 coding region and splice junctions in a large cohort of 163 families with PCD.
What is the bottom line?
Analysis of this large cohort shows that DNAH11 mutations are exclusively seen in patients with PCD with normal ciliary ultrastructure.
Why read on?
Diagnosis of PCD is challenging in patients with normal ciliary ultrastructure.
This large-scale study reports that approximately 22% of all patients with PCD with normal ciliary ultrastructure harbour mutations in DNAH11.
Taken together, this study provides compelling confirmation that there is a genetic basis for PCD in many patients with a presumptive diagnosis of PCD based on clinical features and measurements of nasal nitric oxide, even in the presence of normal ciliary ultrastructure.
Additionally, since DNAH11 encodes an outer dynein arm protein, the number of patients with outer dyenin arms defects were evaluated for mutations in this gene and were negative, showing that despite mutations in DNAH11, outer dynein arms are not affected structurally.
Further, this provides a strong rationale for additional studies to discover other genetic mutations that can cause PCD in patients with normal ciliary ultrastructure.
Introduction
Primary ciliary dyskinesia (PCD) [OMIM# 244400 (http://www.ncbi.nlm.nih.gov/Omim/)] is a rare, genetically heterogeneous disorder. Defective ciliary and/or flagellar function underlies the clinical manifestations, which include chronic oto-sino-pulmonary disease. Situs inversus totalis occurs in around 50% of patients (Kartagener syndrome) and situs ambiguus occurs in at least 6%.1–4
The diagnosis of PCD is important for the initiation of clinical care. The diagnosis largely relies on demonstration of ciliary ultrastructural defects by transmission electron microscopy (EM), but this test fails to support the diagnosis of PCD in patients with normal ultrastructure. Genetic testing holds promise as a diagnostic approach in patients with a clinical phenotype compatible with PCD, as approximately 50% of PCD can be accounted for by biallelic mutations in 12 genes.5–23 Mutations in two genes; dynein axonemal intermediate chain 1 (DNAI1), and dynein axonemal heavy chain 5 (DNAH5), that code for ciliary outer dynein arm (ODA) proteins are the most common genetic causes of PCD (18–30% of PCD),9 10 13 14 and mutations in the remaining genes are relatively uncommon.
DNAH11 encodes a ciliary ODA protein. Mutations in DNAH11 were originally described in a patient with a genetic diagnosis of cystic fibrosis, but who also had features of PCD, but normal ciliary ultrastructure.19 Subsequent reports conclusively demonstrated that mutant DNAH11 causes PCD in patients with normal ultrastructure.19 DNAH11-mutant cilia have a reduced waveform amplitude and hyperkinetic beating pattern.20 21 Based on these findings, a European consensus conference modified the diagnostic algorithm for PCD, and highlighted the importance of high-speed videomicroscopy analysis to evaluate ciliary beat pattern.24
To estimate the mutation frequency in DNAH11 in PCD, we undertook a large study of 163 unrelated patients with PCD displaying a variety of ciliary EM findings, including patients with a compatible PCD phenotype, but without ciliary ultrastructural defects.
Materials and methods
The study included 195 patients with PCD from 163 unrelated families of which 137 were simplex families with only one member affected, 25 were multiplex families with two or more affected siblings and a family with three affected members from an isolated population, and 13 unrelated subjects with isolated situs abnormalities (see online supplement, table E1). The majority of patients were evaluated at the University of North Carolina (UNC) (n=98) or University Hospital, Freiburg (n=38). The remaining patients were evaluated at sites in the Genetic Disorders of Mucociliary Clearance Consortium and other specialised PCD centres in Europe, Australia and Israel (see online supplement). Evaluations included medical and family history, physical examination, spirometry, sputum microbiology, chest radiograph and/or CT scan, and nasal nitric oxide (nNO) measurement in most patients, as described.8 25 The diagnosis of PCD in patients with a compatible phenotype was assessed by ciliary ultrastructure (see below). When ciliary ultrastructure by EM analysis or immunofluorescence was normal, a presumptive diagnosis was made by adjunct tests (ciliary waveform analysis, and/or nNO measurements; see online supplement).11–13 25 26 Subjects with isolated situs abnormalities (n=13) had normal ciliary ultrastructure and nNO, and no clinical features of PCD (online supplement, figure E1). This study was approved by the Committee for the Protection of the Rights of Human Subjects at participating institutions, and written consent was obtained.
Ciliary ultrastructural and waveform analysis
Epithelial cells were obtained by nasal curettage from the inferior turbinate, processed for EM, and ≥20 cilia with adequate images were interpreted at UNC by three blinded observers (JLC, MRK, MWL and/or SLM), as described.8 25 27 28 Videomicroscopy was performed as previously described20 29 30 (details given in online supplement).
Mutation profiling
DNA was extracted from blood, buccal swabs or lymphoblastoid cell lines from proband and available relatives, as described (details given in online supplement).8 25 31 For the evaluation of mutation frequency among unrelated families, one patient with PCD per family was used for the full DNAH11 sequencing and analysis. The majority of sequencing of 82 exons and splice junctions was performed by NHLBI Genotyping and Resequencing Services in Seattle (http://rsng.nhlbi.nih.gov/scripts/index.cfm) using Sanger sequencing. The remainder of the sequencing was performed by Sanger sequencing at UNC (see details and primer sequences in the online supplement, methods and table E2). Estimates of allele frequencies for missense variants were obtained using direct sequencing or restriction endonuclease digestion (online supplement, methods) in at least 104 chromosomes from anonymised patients without PCD (patients with haemophilia) of Caucasian ethnicity. Additionally, 1000 Genomes (http://www.1000genomes.org/) and dbSNP public databases (http://www.ncbi.nlm.nih.gov/projects/SNP/) were searched.
Results
Clinical phenotype of study subjects
Patients with PCD
There were 195 patients (163 families) with PCD (or presumed PCD), including 90 men (46%) and 105 women (54%) between the ages of 2 months and 75 years. Parental consanguinity was present in 21 (13%) families. The majority of families were of Caucasian origin (79%), and the remaining families represented a broad mixture of ethnicities (online supplement, table E1). Situs inversus and situs ambiguus were present in 80 (41%) and 15 (8%) patients, respectively. Most patients had neonatal respiratory distress (70%), recurrent otitis media (82%), sinusitis (95%), and bronchiectasis (70%) by chest CT scan (online supplement, table E1). Of the 101 patients who had nNO measured, the values were low (24.6±22.6 nl/min; mean±SD) compared with values (376±124 nl/min) reported in healthy controls.24 Other details of the clinical features and nNO levels are available (online supplement, table E1). Patients with normal ciliary ultrastructure, according to EM (online supplement, figure E1) or immunofluorescence staining techniques, were considered to have a presumptive diagnosis of PCD. This was based on a compatible clinical phenotype (including bronchiectasis in most patients) and/or situs abnormalities, as well as low levels of nNO and dyskinetic/hyperkinetic waveform and/or increased beat frequency in videomicroscopy studies, consistent with previous reports.20
Subjects with isolated situs abnormalities
There were 13 unrelated subjects with situs abnormalities but no clinical features of PCD, and all subjects who were tested (n=10) had normal nNO levels. Thus, these 13 subjects were considered to have isolated situs abnormalities unrelated to PCD (online supplement, table E1). These subjects were included because mouse models of DNAH11 orthologue32–34 were originally reported to have isolated situs abnormalities without the respiratory phenotype.
Mutation profiling
There were 58 unrelated patients from mutation profiling who had a clinical phenotype, nNO levels, and/or ciliary waveform or situs abnormalities compatible with PCD, but the diagnosis could not be confirmed in the patients or their affected siblings by demonstration of a defect in ciliary ultrastructure. Of these 58 unrelated patients with a presumptive diagnosis of PCD, 20 had at least one mutation in DNAH11, and the clinical demographics, nNO levels, situs status, ciliary phenotype and mutations are summarised in tables 1 and 2.25 30 35 Of these 20 patients, 15 had two (biallelic) mutations, including three homozygotes, and 12 compound heterozygotes (table 1). Seven of the 15 patients with biallelic mutations had an affected sibling with identical mutations (table 2). Most of the 15 families with biallelic mutations had a patient with PCD and situs abnormalities (13 of 15) (table 2), which probably represents an ascertainment bias. As with patients with PCD and ultrastructural defects, there was an age-related distribution of bronchiectasis in patients with biallelic mutations. Three of the six patients without bronchiectasis were ≤8 years old (table 2).
Details of DNAH11 mutations in 20 unrelated patients with primary ciliary dyskinesia (PCD)
Clinical, demographic and ciliary features of 20 unrelated families carrying DNAH11 mutations
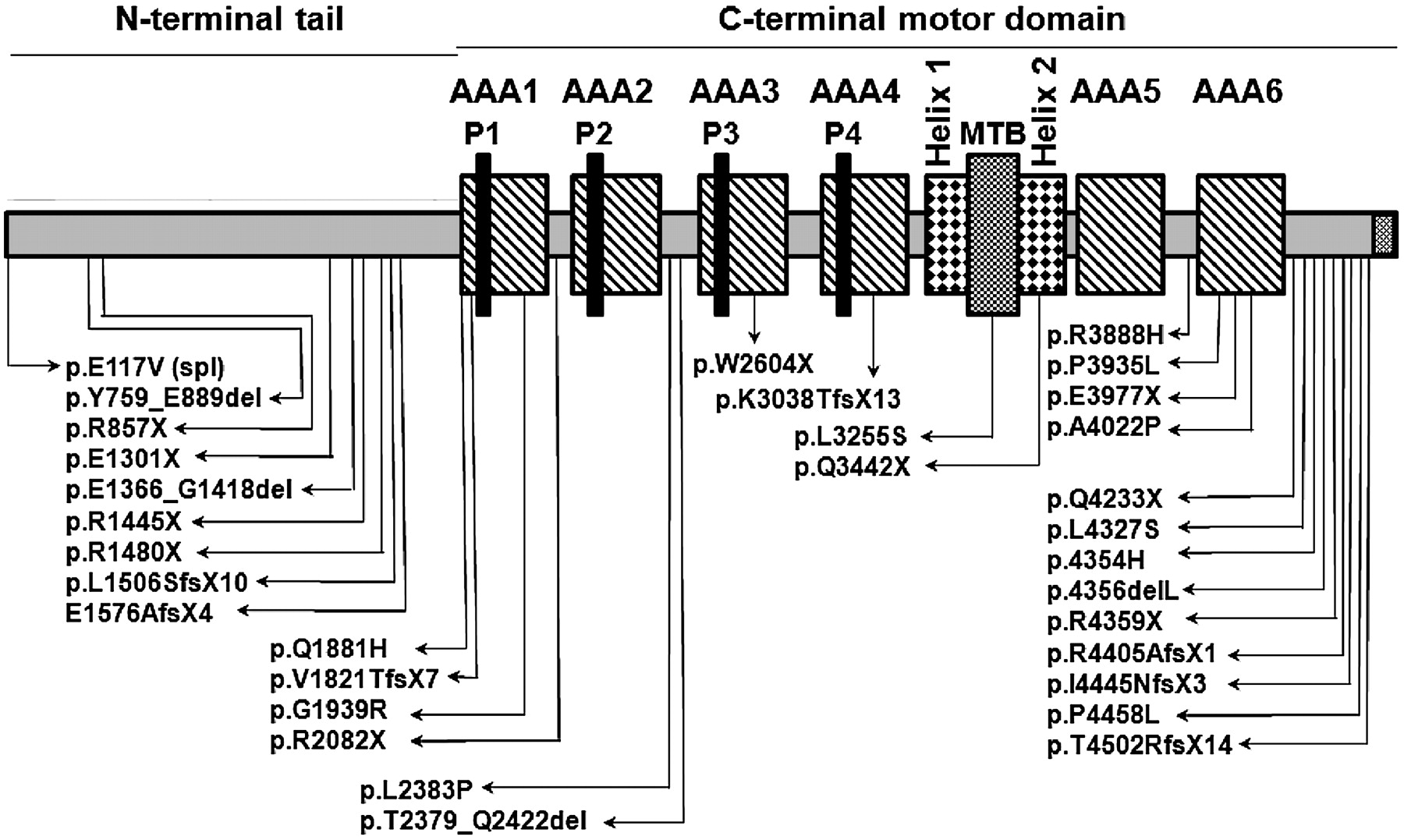
We identified 35 mutant alleles, not previously observed.19–21 These included nonsense mutations (n=11), small insertions-deletions (n=6), splice-site mutations (n=7), and missense mutations (n=11). Except for three patients with homozygous mutations, each mutation appeared only once, which demonstrates extensive allelic heterogeneity (see all 32 unique mutant alleles and their corresponding protein domain in figure 1 and online supplement, table E4). Carrier studies in families showed that mutations were inherited in trans, and segregation analysis was consistent with an autosomal recessive trait. Selected pedigrees illustrate the segregation analysis (figure 2), and additional families in which segregation analysis was possible with either biallelic mutations (online supplement, figure E2) or with only monoallelic mutation (online supplement, figure E3) are presented in the online supplement.

Schematic representation of dynein axonemal heavy chain 11 (DNAH11) (not to scale) showing AAA1–6 domains, four P-loops, the microtubule binding domain (MTB) and helix 1 and 2. The positions of all the mutations are shown.

Representative pedigrees showing autosomal recessive mode of inheritance for dynein axonemal heavy chain 11 (DNAH11) mutations. Segregation analysis from the parents, siblings and the extended family members demonstrates that mutations were inherited in trans (A–D), and there was no bias for gender or situs status. Additional pedigrees are presented in the online supplement figures E2, E3.
cDNA analysis of splice-site mutations
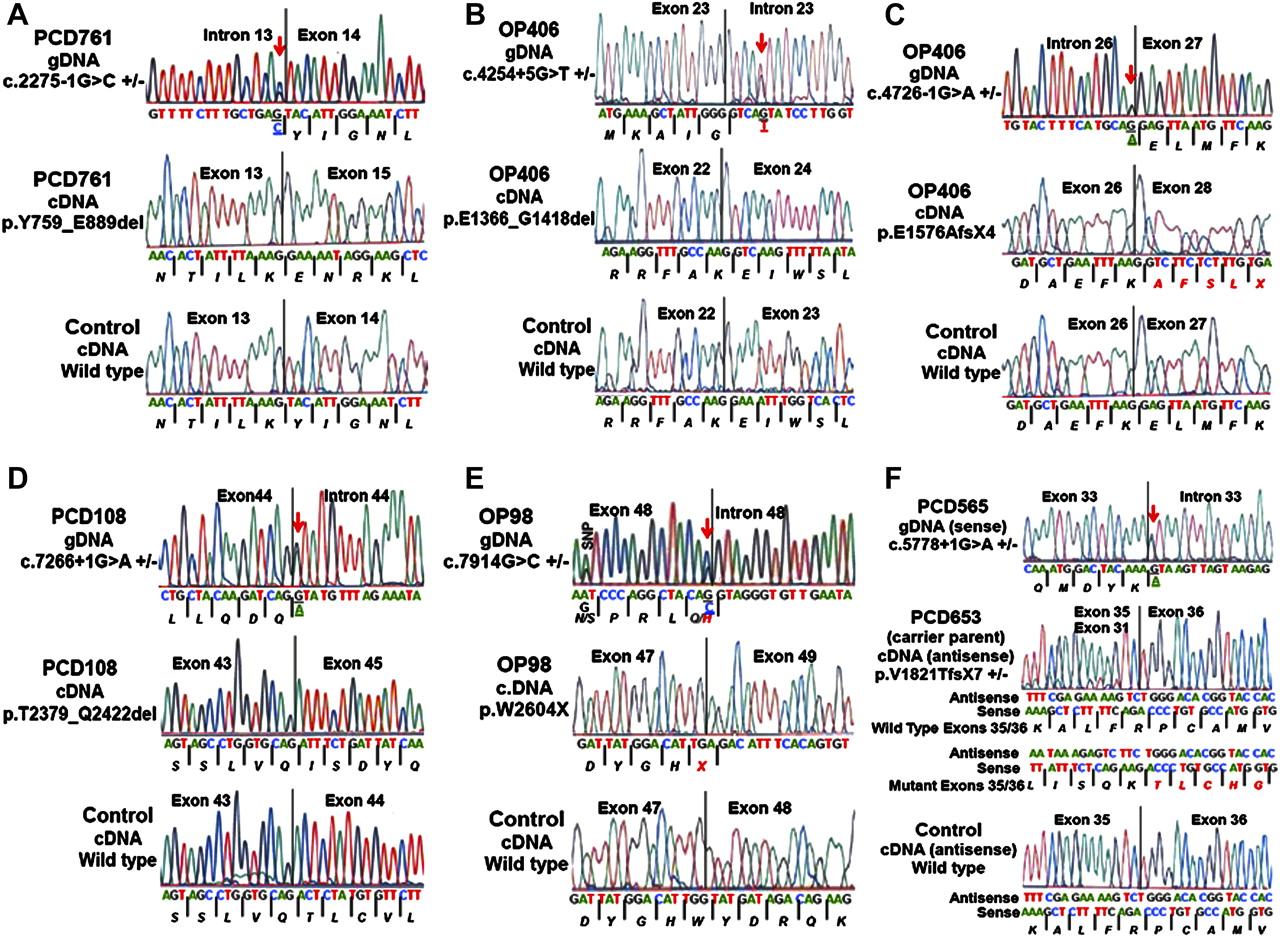
RNA was available for transcript studies for six of the seven splice-site mutations. Three of these splice mutations (c.2275-1G→C; c.4254+5G→T; c.7266+1G→A) caused in-frame deletions of exon 14 (131 amino acids), exon 23 (53 amino acids), and exon 44 (44 amino acids), respectively (table 3, figure 3). Additionally, three mutations (c.4726-1G→A; c.5778+1G→A; c.7914G→C) caused out-of-frame deletions of exon 27, exons 32–35 and exon 48, respectively, leading to premature stop signals (table 3, figure 3).
Effect of DNAH11 splice mutations on cDNA transcript using reverse transcriptase PCR (RT-PCR) in patients with primary ciliary dyskinesia (PCD)
{kind=link}
{kind=link}
{kind=link}
Effect of splice-site mutations on the dynein axonemal heavy chain 11 (DNAH11) transcript using reverse transcriptase PCR. (A) Splice-acceptor site mutation in intron 13 (c.2275-1G→C) in patient PCD761 led to the in-frame deletion of exon 14 that consisted of 131 amino acid residues. (B) Splice-donor site mutation in intron 23 (c.4254+5G→T) in patient OP406-II:2 led to the in-frame deletion of exon 23 that consisted of 53 amino acid residues. (C) Splice-acceptor site mutation in intron 26 (c.4726-1G→A) in patient OP406-II:2 led to out-of-frame deletion of exon 27, and resulted in a premature stop signal. (D) Splice-donor site mutation in intron 44 (c.7266+1G→A) in patient PCD108 led to the in-frame deletion of exon 44 that consisted of 44 amino acid residues. (E) Splice-donor site mutation in exon 48 (c.7914G→C) in patient OP98-II:1 led to out-of-frame deletion of exon 48, and resulted in a premature stop signal. (F) Splice-donor site mutation in intron 33 (c.5778+1G→A) in patient PCD565 led to out-of-frame deletion of exons 32–35, and resulted in a premature stop signal. The cDNA was available only from the carrier parent of patient PCD565, which was used to check the transcript. All of the six panels with three electropherograms each shows the genomic location of the mutation (top) with a red arrow and bases underlined, mutant cDNA transcript (middle) and wild-type transcript (bottom). Amino acid residues are italicised and the protein product due to the out-of frame mutation is shown in red font. The genomic base change for the mutation is underlined. A known single nucleotide polymorphism (SNP) was observed in OP98-II:1 and its location is shown. Further details on reverse transcriptase PCR are shown in table 3 (primer sequences shown in online supplement, table E3).
Correlation of genotype with ultrastructure and ciliary waveform
The genetics of PCD involves locus, allelic and ultrastructural heterogeneity; thus, we studied patients with different ciliary EM findings, including patients with normal ultrastructure, but compatible clinical phenotype. Mutations in DNAH11 were exclusively seen in patients with a clinical phenotype of PCD and normal ciliary ultrastructure. Each of the 14 patients (11 families) with biallelic mutations in DNAH11 who were tested by videomicroscopy had the characteristic hyperkinetic beating pattern and reduced waveform amplitude, as previously reported (see table 2 and online supplement, movies E1 and E2).20 None of the other groups carried mutations, including patients with isolated situs abnormalities. In total, we identified biallelic DNAH11 mutations in 13 (22%) of the 58 unrelated families with compatible clinical phenotype, low nNO and confirmed normal ciliary ultrastructure and/or abnormal videomicroscopy. Despite full gene (coding region) sequencing, we found only one mutant allele in five patients (four with confirmed normal ultrastructure), which could reflect either a second mutation in DNAH11 (introns or promoter regions, or large indels), or a heterozygous mutation in a different ciliary gene (which would represent a digenic mode of inheritance), or biallelic mutations in a PCD gene other than DNAH11.
Population studies
There were 10 unique missense variants, one possible single nucleotide polymorphism (SNP), two splice mutations and one amino acid deletion that were studied to examine their role as pathogenic or benign. Due to the nature of the sequence-based assay, certain amplicons (exons 33, 44 and 80) harboured splice and nonsense mutations in addition to variants of interest, and they were examined as well. Each of these variants was identified in only one of the 163 unrelated patients with PCD who were tested, and never identified in 13 with isolated situs abnormalities. Additionally, these missense variants were not observed in at least 104 alleles tested in subjects without PCD, ethnically matched when possible (ethnically matched controls were not available for three subjects). In addition, these variants were predicted to be deleterious based on in silico program ‘Mutation Taster’ (http://neurocore.charite.de/MutationTaster/). Furthermore, none of these missense variants or loss-of-function or splice mutations were seen in 1000 Genomes (http://www.1000genomes.org/) or dbSNP (http://www.ncbi.nlm.nih.gov/projects/SNP/) databases, except for having been listed from this study in dbSNP. Taken together, these data suggest that these variants are not benign polymorphisms (online supplement, table E5).
Polymorphisms and variants of unknown significance
DNAH11 is a large gene, and we identified 310 novel and/or known polymorphisms. The polymorphisms and corresponding SNP database number (http://www.ncbi.nlm.nih.gov/SNP/) are available (see online supplement, table E6). The novel variants that are not present in the SNP database were considered benign due to the high minor allele frequency in patients with PCD (online supplement, table E6, footnotes). One rare variant (c.11059A→G; p.K3687E) was seen on only one allele of a patient with PCD and an ODA defect, and was not seen in either control or isolated situs abnormality groups. This was a non-synonymous substitution, conserved (80%) across species, and present at the third last base of exon 67 near the splice-donor site. Due to the unavailability of RNA, we could not check the effect of this variant on splicing. We classified this substitution as a variant of uncertain significance, because mutations in DNAH11 are seen (otherwise) exclusively in patients with normal ciliary ultrastructure; plus, a second mutation was not identified, despite full gene sequencing.
Errors in published sequence of DNAH11
During analysis of cDNA from nasal epithelial cells and lymphoblastoid cell lines from two unrelated control subjects, we observed errors in the Ensembl database (http://uswest.ensembl.org/index.html), and published sequence of DNAH11.19 The last 15 bases of exon 22 (and five amino acid residues) are not present in the DNAH11 transcript from multiple control subjects (details in bottom panel of figure 3B and online supplement, figure E4A). These five amino acids were previously shown in the human DNAH11,19 but not in other species, which is congruent with sequence error. Additionally, six bases in exon 32 of the Ensembl database (and two amino acid residues) are not present in the DNAH11 transcript from multiple control subjects (correct cDNA sequence for exons 22 and 32, and multiple sequence alignment in online supplement, figure E4). Due to errors in the publicly available sequences, the full-length DNAH11 will contain 4216 amino acids and the mutation nomenclature for all the previously published mutations (and variants/SNPs) will change (see online supplement, table E7 for mutation nomenclature that corresponds with the current and formerly published sequenced information). The Genbank (www.ncbi.nlm.nih.gov/genbank/) accession numbers for the updated DNAH11 exons 22 and 32 sequences are JQ247524 and JQ247523 respectively.
Discussion
It is challenging to confirm a diagnosis of PCD in patients with a compatible clinical phenotype, but who do not have hallmark defects in ciliary ultrastructure. Some specialised centres use nNO measurement as an aid to diagnosis. A few centres use videomicroscopy to evaluate ciliary waveform to confirm the diagnosis, but this assay is difficult and limited in availability.
Mutations in DNAH11 have been reported in four families in which patients with PCD have normal ciliary ultrastructure.19–21 However, the prevalence of DNAH11 mutations, and genotype–ciliary phenotype correlations, are not well defined. In this study, we tested the hypothesis that mutations in DNAH11 are a relatively common cause of PCD in patients with normal ciliary ultrastructure. We studied a large number of well characterised patients with PCD and different ciliary ultrastructural phenotypes to determine the frequency of DNAH11 mutations in each group.25 In patients with normal ciliary ultrastructure, the clinical phenotype was typical of PCD, including a high prevalence of respiratory distress in full-term neonates, chronic otitis media and sinusitis, productive cough, bronchiectasis, situs abnormalities and infertility (online supplement, table E1). In addition, these patients had low nNO and/or abnormal immunofluorescence with ciliary antibodies and/or abnormal ciliary waveform with limited range of motion and hyperkinesis, which are compatible with PCD (tables 1 and 2).20
We determined that biallelic mutations in DNAH11 are relatively common (22%) in patients with PCD without a defined ciliary ultrastructural defect (table 1). None of the patients with PCD and ultrastructural defects had mutations in DNAH11. Thus, disease-causing mutations in DNAH11 appear specific for patients with PCD and normal ciliary ultrastructure. It is difficult to determine the proportion of all patients with PCD carrying biallelic mutations in DNAH11, since the fraction of patients with PCD and normal ciliary ultrastructure is not known. However, several studies, and the experience of our centres, estimate that at least 30% of patients with PCD have normal axonemal ultrastructure2; thus, DNAH11 mutations may occur in around 6–7% of all patients with PCD.
Segregation analysis in families was consistent with trans allelic inheritance of the mutation as an autosomal recessive trait (table 2, figure 2 and online supplement, figures E2 and E3). Pedigree analysis showed horizontal transmission, and carrier analysis showed that parents carried the mutation, but were clinically unaffected; therefore, autosomal dominant inheritance was ruled out (online supplement, figure E2). In the five patients in whom a second mutation was not identified, it is likely that a second mutation in DNAH11 is present but not discovered by sequence analysis (eg, promoter, intronic or large insertions-deletions).15 Alternatively, a few of these patients may only be a carrier of a DNAH11 mutation, and the actual biallelic PCD-causing mutations are present in a different gene. Finally, there might be a heterozygous mutation in another axonemal gene, and (together with the identified DNAH11 mutation), would represent a digenic mode of inheritance; however, digenic inheritance has never been reported in PCD.
Of the 20 unrelated patients carrying mutations, there were 35 mutant alleles, including seven splice-site mutations (table 1). These splice-site mutations abrogated splicing in all six cases tested, which resulted in shorter DNAH11 transcripts (table 3, figure 3). We also made the following conclusions: the p.E117V splice-donor site variant (when RNA was not available) and 10 missense variants were likely disease causing because each variant was seen only once, and not seen in the dbSNP and 1000 Genomes databases; variants were absent in control subjects who were tested; the majority of missense mutations had a loss-of-function mutation on the trans allele; the amino acid affected by the missense mutations was highly conserved across species, and in silico analyses predicted it to be deleterious; and the majority of missense mutations were in a conserved AAA module or were on a microtubule binding domain (table 1 and figure 1). We also discovered some errors in the published sequence of DNAH11; thus, the mutation nomenclature needs to be updated based on the currently revised sequence (online supplement, table E7, figure E4).
The ability to establish (or rule out) a diagnosis of PCD by a genetic test in patients with a compatible phenotype and normal ciliary ultrastructure is significant at several levels. For example, several reports suggest that the vast majority (around 90%) of patients with PCD have defined ultrastructural defects.2 3 36 37 However, this perspective may greatly underestimate the number of patients with PCD and normal ciliary ultrastructure, particularly in patients with normal situs status. At an individual case level, the importance of being able to establish (or exclude) PCD by a genetic test is demonstrated by the situation in one of our families (UNC101; figure 2C), in which one woman (#623) had a compatible clinical phenotype and low levels of nNO consistent with PCD, but no situs abnormalities. Her sister (#627) also had some clinical features of PCD, as did an 8-year-old paternal half sister (#635). Before genetic testing was possible, we were unable to clarify the diagnosis of PCD in this family. Subsequently, we defined biallelic nonsense mutations in DNAH11 in the proband and the full sibling, but the half sibling did not carry any mutation.
There are some instructive genotype–phenotype correlations in Chlamydomonas and murine orthologs of mutant DNAH11. The Chlamydomonas reinhardtii orthologue of DNAH11 is β-dynein heavy chain (β-DHC), and Chlamydomonas mutants of β-DHC can assemble outer arm subunits into the flagellar axoneme, but swimming velocity and/or beat frequency are reduced.38–41 In humans, immunofluorescence studies show normal distribution of ODA proteins (DNAH9 and DNAH5) in a patient with biallelic DNAH11 mutations.29 Thus, mutant DNAH11 does not cause defective ODA assembly, but causes defective ciliary function.2 20 The mouse orthologue of DNAH11 (Dnahc11) is left-right dynein (lrd) and lrd null mice have situs defects.32 34 The spontaneously occurring mouse model of Dnahc11 (inversus viscerum mutant; iv/iv) has situs defects and recent work shows these mice have no detectable ciliary beat frequency, and suffer otitis media and rhinitis, even though they have normal ciliary ultrastructure.32 33 42
In conclusion, our large-scale mutation analysis indicates that biallelic mutations in DNAH11 occur in 22% of patients with a clinical phenotype of PCD, but normal ciliary ultrastructure, and is consistent with an autosomal recessive mode of inheritance. Transcript analysis of six splice-site mutations revealed abrogation of normal splicing. These data clearly establish that clinical disease (PCD) occurs in patients with normal ciliary ultrastructure. This study also demonstrates that genetic analysis of DNAH11 can be useful to assist in the diagnosis of PCD, and supports the concept to search for additional genetic origins of PCD.
Web resources
At a glance commentary
Primary ciliary dyskinesia (PCD) is an autosomal recessive, genetically heterogeneous disorder with oto-sino-pulmonary disease. Most patients are diagnosed on the basis of ciliary ultrastructural defects. This study identified biallelic mutations in DNAH11 in 22% of 58 unrelated patients with normal ciliary ultrastructure, which validates the concepts of ciliary dysfunction in the presence of normal ultrastructure, and the use of genetic analysis to facilitate the diagnosis of PCD.
Acknowledgments
We are grateful to all the patients with PCD and the family members for their participation in this research. The authors would like to thank Ms Michele Manion, who founded the US PCD Foundation. We are indebted to other investigators and the coordinators of the ‘Genetic Disorders of Mucociliary Clearance Consortium’ which is part of the Rare Disease Clinical Research Network (http://rarediseasesnetwork.epi.usf.edu/gdmcc/index.htm), including Dr Jeffrey Krischer (Data Management and Coordinating Center, Tampa, Florida, USA), Mr Reginald Claypool, Ms Tanya Glaser and Ms Meghan O'Connell (National Institute of Allergy and Infectious Diseases, Bethesda, Maryland, USA), Dr Jeffrey Atkinson and Ms Jane Quante (Washington University in St Louis, Missouri, USA), Ms Shelley Mann (The Children's Hospital, Denver, Colorado, USA), Drs Ronald Gibson and Moira Aitken and Ms Sharon McNamara (Children's Hospital and Regional Medical Center, Seattle, Washington, USA), Dr Carlos Milla and Ms Jacquelyn Zirbes (Stanford University Medical Center, Palo Alto, California, USA), Ms Donna Wilkes (The Hospital for Sick Children, Toronto, Ontario, Canada), and Ms Caroline O'Connor (University of North Carolina at Chapel Hill, USA). The authors also thank Drs Larry Ostrowski, Peadar Noone, Hilda Metjian, Deepika Polineni, Adam Shapiro, Jessica Pittman and Mr Kunal Chawla for thoughtful discussion; Ms Lu Huang and Ms Rhonda Pace for technical assistance; and Ms Elizabeth Godwin and Ms Cindy Sell for administrative support. The authors would like to acknowledge the following people for providing DNA analysis for patients with PCD and their families: Dr Eitan Kerem from Hadassah University Hospital, Israel; Dr. H. Blau from Schneider Medical Center of Israel, Israel, Dr Israel Amirav from Ziv Medical Center, Israel, Dr Lucy Morgan from Concord Hospital, Australia, Dr Robbert de Iongh from University of Melbourne, Australia, Dr Scott Bell from The Prince Charles Hospital, Australia, Dr Hannah Mitchison from University College London, England, Dr Ugo Pradal from Cystic Fibrosis Center Verona, Italy.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Movie 1
- Movie 2
Footnotes
See Editorial, p 377
Funding MRK, MWL, JLC, MJH, SLM, SDD, TWF, KNO, SDS, MR, KEB, MCA, AL and MAZ are supported by National Institute of Health research grant 5 U54 HL096458-06, funded by the Office of the Director, and supported by ORDR and NHLBI, NIH. MRK and MAZ are supported by National Institutes of Health grant 5 R01HL071798. TWF is supported by R01 HL08265 and Children's Discovery Institute. KNO is supported by the Intramural Research Program of the National Institute of Allergy and Infectious Diseases. JLC is supported by Clinical Innovator Award by Flight Attendant Medical Research Institute. HO is supported by a grant from the Deutsche Forschungsgemeinschaft (DFG Om 6/4, GRK1104, SFB592). Resequencing was provided by the University of Washington, Department of Genome Sciences, under US Federal government contract number N01-HV-48194 from National Heart, Lung, and Blood Institute. This work was supported in part by grants RR00046, UL1 RR025747 and UL1 RR025780 from the National Center of Research Resources, NHLBI P01 HL034322, NIH and CFF R026-CR07. This consortium, Genetic Disorders of Mucociliary Clearance is part of NIH Rare Diseases Clinical Research Network (RDCRN). Funding and/or programmatic support for this project was provided by grant 5 U54 HL096458-06 from the NHLBI and the NIH Office of Rare Diseases Research (ORDR). The views expressed do not necessarily reflect the official policies of the Department of Health and Human Services; nor does mention by trade names, commercial practices, or organisations imply endorsement by the U.S government.
Competing interests The first and the last authors are part of the patent for DNAH11 gene mutations.
Ethics approval IRB at the University of North Carolina.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement We will abide by the data-sharing policy of the journal.