Article Text

Abstract
Background Recent genetic and animal studies have implicated matrix metalloproteinase-12 (MMP-12) in the pathogenesis of chronic obstructive pulmonary disease (COPD). It has previously been shown that individuals homozygous for the A/A allele of rs652438 in MMP-12 are over-represented among patients with severe COPD (n=1517). A study was undertaken to examine the functional basis of these findings.
Methods rs652438 A and G variants were generated by site-directed mutagenesis and transfected into COS7 cells where they were expressed. Casein zymography and a specific FRET activity assay were used to compare MMP-12 activity between alleles. Cell migration was examined using a transwell assay. Patients from two COPD cohorts were genotyped for rs652438 and associated with inflammatory cell number in bronchoalveolar lavage fluid (n=10) and induced sputum (n=262); the emphysema score (n=1428) was assessed by CT scanning.
Results Mean MMP activity was 2.95-fold higher by zymography (p=0.0049) and 3.45-fold higher by FRET assay (p=0.0001) for the A allele than the G allele. Mean migration of COS7 cells expressing the A allele was 2.31-fold greater than for those expressing the G allele (p=0.0001). Macrophage numbers were greater in bronchoalveolar lavage fluid (1.28-fold increase, p=0.033) and induced sputum (1.58-fold increase, p=0.083) of A/A individuals compared with A/G heterozygotes. The presence of the A allele was dose-dependently associated with increased emphysema (p=0.016).
Conclusions The rs652438 SNP alters MMP-12 activity with the A allele being more active, which is associated with increased macrophage infiltration and emphysema in the lungs of patients with COPD. These findings further implicate MMP-12 and this SNP in COPD.
- rs652438
- matrix metalloproteinase
- 1082A/G
- 357Asn/Ser
- emphysema
- COPD mechanisms
- emphysema
- macrophage biology
Statistics from Altmetric.com
- rs652438
- matrix metalloproteinase
- 1082A/G
- 357Asn/Ser
- emphysema
- COPD mechanisms
- emphysema
- macrophage biology
Key messages
What is the key question?
How does a single nucleotide polymorphism in the haemopexin-like domain of MMP-12 protect patients against severe COPD?
What is the bottom line?
The protective variant of MMP-12 reduces metalloproteinase activity and cell migration in vitro and is associated with lower lung macrophage numbers and emphysema in patients with COPD.
Why read on?
By investigating how this genetic variant in MMP-12 protects against severe COPD, this study addresses one of the factors contributing to the idea of the ‘susceptible smoker’ in COPD.
Introduction
It is widely accepted that alveolar destruction in chronic obstructive pulmonary disease (COPD) is the result of an imbalance of proteolytic enzymes and their inhibitors based on the observation that severe α1-antitrypsin deficiency predisposes cigarette smokers to the development of pulmonary emphysema. This is thought to be due to uninhibited neutrophil elastase activity degrading elastin, a major component of lung connective tissue.1
A number of other proteolytic enzymes are also capable of degrading elastin, including the matrix metalloproteinases (MMPs). These proteases can degrade all the major protein components of the extracellular matrix (ECM). Evidence from genetic studies, animal models and human disease suggest that MMP-12, the major metalloelastase derived from alveolar macrophages, plays a role in lung destruction in COPD.2–6 Recently, a large multiple cohort association study linked variation in MMP-12 with both lung function in smokers and the risk of developing COPD2 while, in animal models of COPD, deletion of MMP-12 in mice exposed to cigarette smoke protects these animals from emphysema.3 In patients with COPD elevated levels of MMP-12 are present in the airways4 5 and increased numbers of MMP-12 expressing macrophages.6
We recently performed a case–control genetic association study into MMPs 1, 9 and 12. This identified haplotypes in MMP-12 of two single nucleotide polymorphisms (SNPs), rs652438 and rs2276109, which were associated with severe and very severe COPD (GOLD stages III and IV). In the study by Hunninghake et al2 rs652438 was not associated, possibly due to different phenotypes tested. This suggests that these SNPs are modifiers of disease severity, and supports the idea that MMP-12 is a mediator of lung damage in COPD.7 In our previous study about 18% of Europeans were found to have at least one protective allele at these SNP positions, giving a significantly reduced risk of severe disease development.7
The SNPs at rs2276109 and rs652438 are located in the promoter region of the gene and haemopexin domain of the enzyme, respectively, and could directly influence MMP-12 activity through altered expression or substrate binding. The former has been demonstrated for rs2276109, where the A allele in the MMP-12 promoter results in increased MMP-12 transcription.8 The rs652438 A/G SNP causes a missense change of asparagine to serine at amino acid position 357, located in exon 8, which encodes the haemopexin domain. The haemopexin domain of MMP-12 is involved in substrate specificity and may interact with the enzyme's catalytic domain.9 We hypothesised that this SNP could alter the function of MMP-12, thereby altering the pathological processes in COPD.
Material and methods
Additional data on methods are provided in the online supplement.
Patient cohorts
Two cohorts were used in this study, a local Nottingham cohort of patients with COPD10 and ECLIPSE (Evaluation of COPD Longitudinally to Identify Predictive Surrogate End-points).11
Nottingham cohort
We recruited smokers with COPD defined by GOLD criteria. Patients were aged 40–80 years, had a smoking history of at least 10 pack-years and had not required antibiotics or oral steroids for the previous 6 weeks. Patients with α1-antitrypsin deficiency, radiological evidence of interstitial lung disease, previous thoracic surgery or taking inhaled corticosteroids were excluded. Pre- and post-bronchodilator spirometry was performed according to American Thoracic Society guidelines and breathlessness was recorded using the Medical Research Council (MRC) dyspnoea score. Those with reversibility >10% of baseline forced expiratory volume in 1 s (FEV1) after inhaling 400 μg salbutamol were excluded. Subjects with FEV1/forced vital capacity (FVC) of <70% were classified as having COPD.
ECLIPSE cohort
Patients with COPD aged 40–75 years with baseline post-bronchodilator FEV1 <80% of the predicted value, baseline post-bronchodilator FEV1/FVC of <0.7 and a smoking history of >10 pack-years were enrolled. Exclusion criteria were related to diagnosis (known respiratory disorders other than COPD and severe α1-antitrypsin deficiency), prior medical history (known history of significant inflammatory disease other than COPD), a COPD exacerbation within 4 weeks of enrolment, having undergone lung surgery, recent diagnosis of cancer, having received a blood transfusion in the 4 weeks before the start of the study, inability to walk, taking part in a blinded drug study, treatment with oral corticosteroids at inclusion and participation in studies with radiation exposure.
Information on the cell percentage composition of bronchoalveolar lavage fluid (BAL) was provided for the Nottingham cohort. Absolute cell number and emphysema scores were provided for ECLIPSE, providing some replication for the cell number measure described by the Nottingham cohort. Emphysema scores were obtained using CT scanning performed at full suspended inspiration. A multislice CT scanner was used, taking contiguous images at 1 or 1.25 mm thickness. Scanning was set at 120 kVp and 40 mAs. A low spatial frequency reconstruction algorithm was used for analysis (GE-standard, Siemens Healthcare, Erlangen, Germany). Genotypes of rs652438 were determined in the Nottingham cohort by direct sequencing. ECLIPSE genotypes were provided using methods described elsewhere.11
Vector creation
MMP-12 (pCMV-XL4 vector) was obtained from Origene (Rockville, USA). The rs652438 SNP G allele was introduced via the GeneEditor kit (Promega, Southampton, UK) according to the manufacturer's instructions using mutagenic primer 5′-TAATTAGCAGTTTAAGACC-3′. After DNA preparation (Qiagen, Crawley, UK), inserts were sequenced and aligned using ClustalW (http://www.ebi.ac.uk/Tools/clustalw2/index.html) to confirm inserts were identical apart from the SNP. The vector without an insert was used as null control in all experiments.
Cell culture
COS7 cells were grown in DMEM using 10% fetal bovine serum at 37°C and 5% CO2. Vectors were transiently transfected using Fugene 6 (Roche, Mannheim, Germany). After 24 h incubation, cells were serum starved, supernatants collected after 24 h and stored at −80°C.
Analysis of enzymatic activity
Casein zymography was used to perform analysis of activity as detailed elsewhere.12 Changes to the protocol were 12% casein gels (Invitrogen, Paisley, UK) and quantification against 10 ng recombinant human MMP-12 (R&D Systems, Minneapolis, USA). Images were scanned and analysed using Image J V.1.40 (Wayne Rasband, NIH, USA). Bands were normalised against standard and presented as a ratio.
MMP-12 activity was also measured using the Sensolyte 520 MMP-12 assay kit (Anaspec, San Jose, California, USA) according to the manufacturer's instructions. The assay was run on a Flexstation with Softmax Pro software (Molecular Devices, Sunnyvale, California, USA) at excitation/emission wavelengths of 490/520 nm respectively. Recombinant human MMP-12 (10 ng) was the reference standard. Substrate-only samples were included for background subtraction purposes. All measurements were performed in triplicate and the experiments were repeated three times.
Western blot
Supernatants and human MMP-12 standard (10 ng) were resolved using 10% SDS page gels followed by blotting onto Hybond-P membrane (GE Healthcare, Buckinghamshire, UK) and probed with IgG anti-human MMP-12 catalytic antibody (R&D Systems, Minneapolis, USA). HRP-conjugated goat anti-mouse IgG (Sigma Aldrich, Gillingham, UK) was used. Visualisation was performed using the ECL western blotting detection kit (GE Healthcare).
Cell viability assays
Total cell counts and cell viability measurements were performed using a haemocytometer and MTT assay as described elsewhere.12 13 Quantification of total protein levels in supernatants was performed using a Bradford based method14 (Bio-Rad Protein Assay, Berkley, California, USA).
Real-time PCR
Quantification of MMP-12 mRNA was performed as described elsewhere.12 β-actin was used for normalisation. Primer sequences are as follows: MMP-12 forward: 5′-CATTCAGGAGGCACCAACTTGTTC-3′, reverse: 5′-CCTTTGGATCACTAGAATGGCC-3′; β-actin forward: 5′-GGATGCAGAAGGAGATTACTG-3′, reverse: 5′-CGATCCACACAGAGTACTTG-3′.
Minigene analysis of splicing
Exonic splice silencing was analysed using the Minigene technique. Details of the method used are provided in the online supplement.
Migration assay
The transwell migration assay was performed as previously described.12 Changes to the protocol were use of collagen IV (0.1 mg/ml in serum-free medium) coating and 10% serum as a chemotactic stimulus. Migration was measured at 8 h in three replicates in three independent experiments.
Statistical analysis
Data sets were tested for normality using the D'Agostino and Pearson test and analysed using t tests (Prism 4, Graphpad Software, La Jolla, USA). Linear regression of SensoLyte 520 kinetic assay was also performed in Prism 4. Cohort characteristics were investigated using the descriptive statistics function in SPSS V.14 (SPSS Inc). Analysis of the ECLIPSE cohort was performed using a linear regression model adjusted for confounding factors in SPSS V.14. For all tests, p values of <0.05 were considered statistically significant.
Results
Study population characteristics
The characteristics of the populations used in this study are detailed in table 1.
Basic characteristics of COPD cohorts
Analysis of enzymatic activity
MMP-12 activity was examined using casein zymography of COS7 supernatants from cells expressing the A and G constructs. COS7 cells expressing the null vector had no detectable caseinolytic activity (limit of detection 1 ng per lane). In supernatants from COS7 cells expressing the MMP-12 constructs, caseinolytic activity was present at 54 kDa and 45 kDa, corresponding to the pro- and intermediate-active forms of MMP-12 (figure 1). Using the t test and comparison of the means, densitometry of caseinolytic bands showed that the activity of the A allele was 2.95-fold (p=0.0049) higher than that of the G allele.

(A) Western blot of rs652438 A and G paired transfections. Expression levels of matrix metalloproteinase-12 (MMP-12) were found to be equal between the transfection of the different constructs. (B) Representative casein zymogram showing expression of the pro and intermediate forms of MMP-12 from vector-transfected cell supernatants. Below, densitometry of three independent experiments. The A allele has mean±SEM band intensity of 77.11±14.26% compared with the 10 ng MMP-12 standard while the is G allele has a band intensity of 26.11±6.39%. The A allele activity is 2.95-fold higher than the G allele (p=0.0049). (C) Sensolyte 520 MMP-12 assay endpoint read. The A allele has mean±SEM activity of 48.30±3.44% compared with the 10 ng MMP-12 standard while the is G allele has 14.13±1.30% activity. The A allele activity is 3.42-fold higher than the G allele (p=0.0001; n=3).
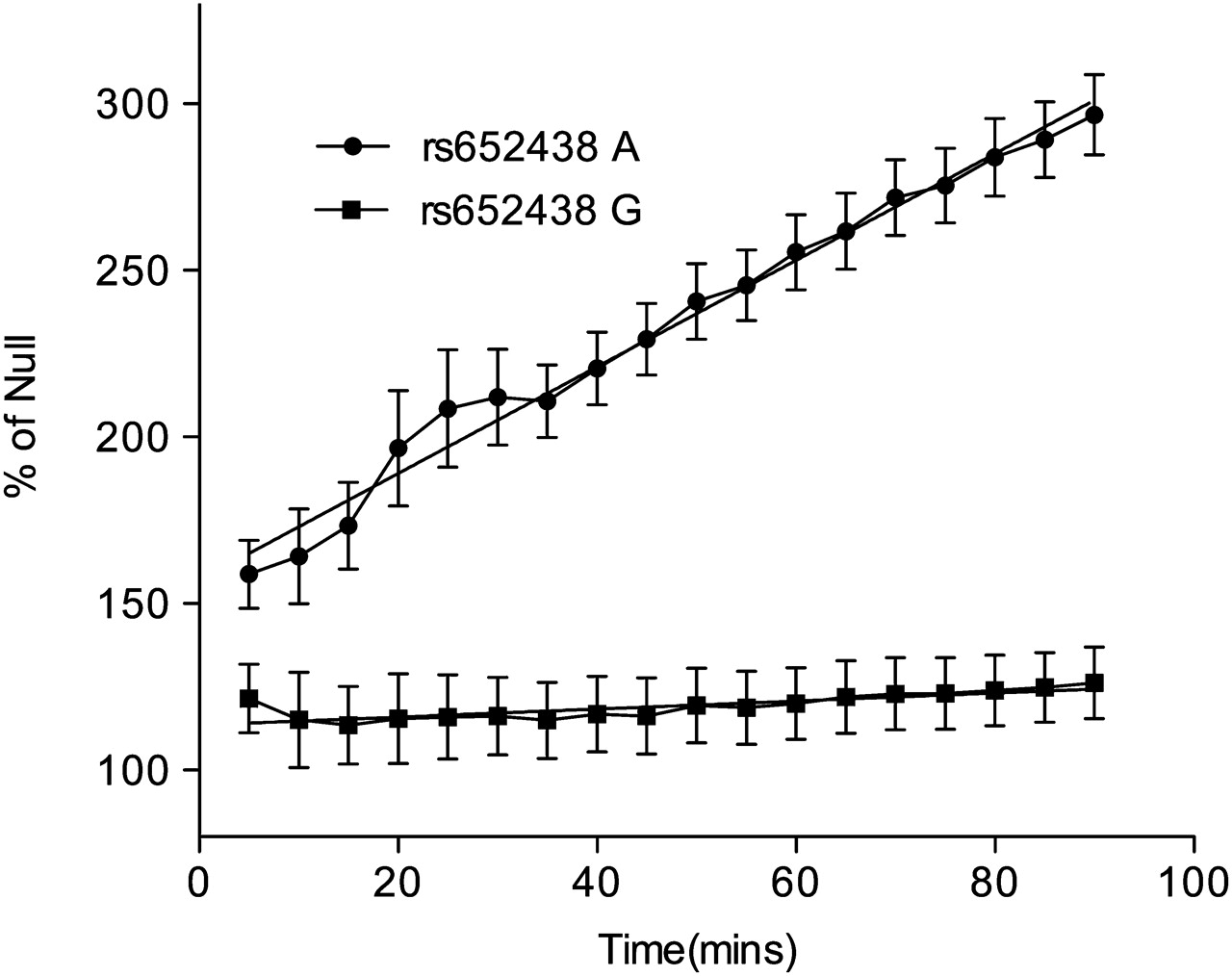
We next used the Sensolyte 520 MMP-12 ELISA. Using the t test and comparison of the means, the A allele of rs652438 was 3.42-fold (p=0.0001) more active than the G allele (figure 1). The Sensolyte assay was also used to provide a kinetic read of MMP-12 activity (figure 2). Using the linear regression function in Prism, the slope of substrate cleavage against time for the A allele was 1.6 CI±0.106 relative fluorescence units/min (RFU) (p=0.0001 vs empty vector) and the G allele slope was 0.12 CI±0.096 RFU (p=0.22 vs empty vector). The MMP-12 activity of the G allele was not different (p=0.2153) from the null vector baseline.
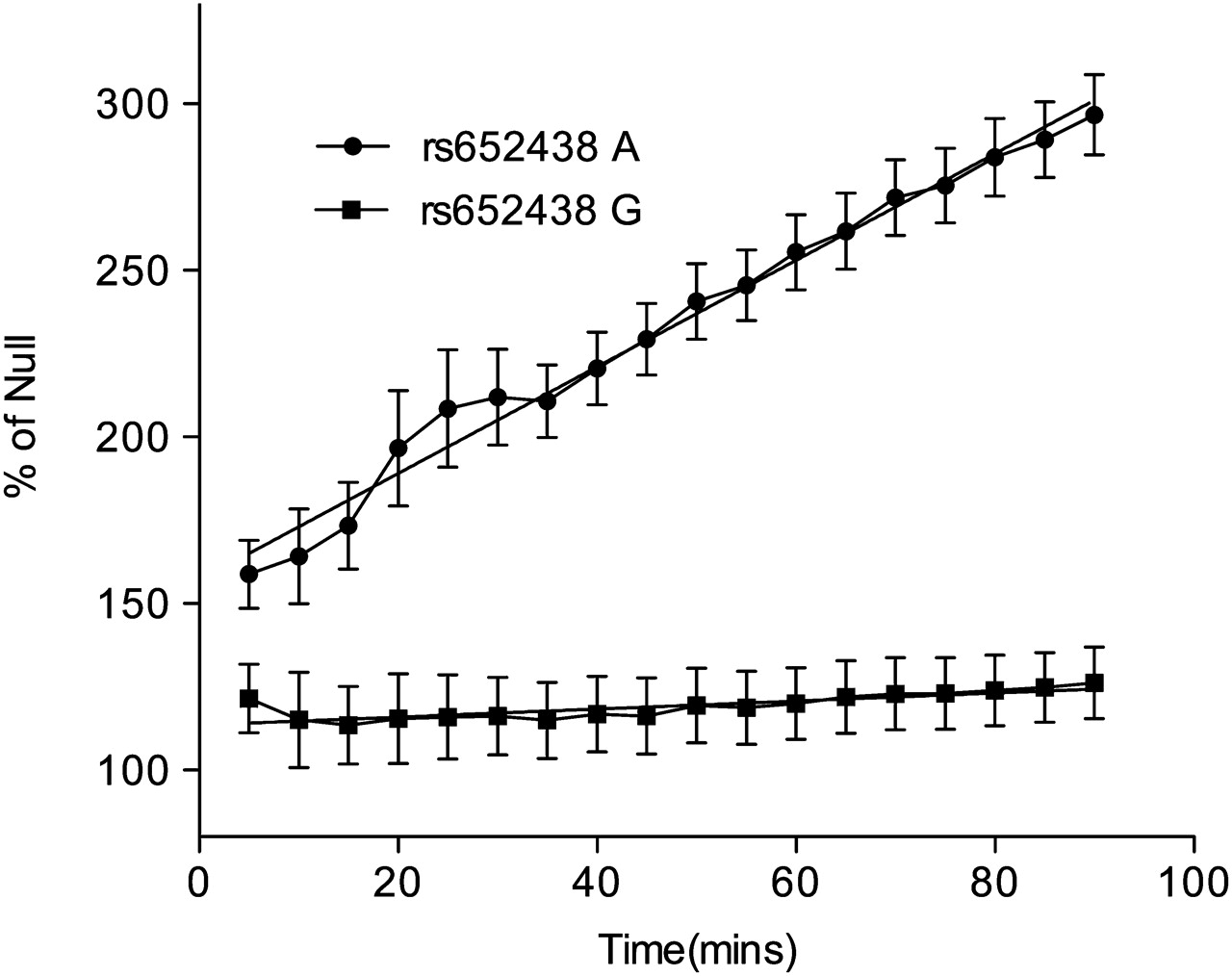
Sensolyte 520 MMP-12 assay kinetic read. The A allele of rs652438 is significantly different from the null allele baseline slope (1.6000±0.1062, p=0.0001) while the G allele is not significantly different from the baseline slope (0.1201±0.0963, p=0.2153; n=3).
Effect of SNP on cell viability and MMP-12 expression
Examination of MMP-12 constructs was performed to determine whether the SNP affects MMP-12 mRNA expression, cell number or viability. Real-time PCR and Minigene determined that the rs652438 polymorphism did not influence mRNA levels (figure 3). Using the t test, no significant differences were seen in cell number, supernatant total protein or viability (figure 4).

(A) Real-time PCR demonstrating robust and equal expression of matrix metalloproteinase-12 (MMP-12) constructs (n=3). No endogenous MMP-12 was detected. (B) Real-time PCR of exon trap vector with MMP-12 exon 8 inserted. The 160 bp exon 8 is transcribed along with the vector-specific exons, giving a 417 bp band. Because both bands are 417 bp in size and of equal intensity, exonic silencing does not occur when the G allele is present. If silencing had occurred, a 257 bp band would be formed with only the vector-specific exons transcribed.
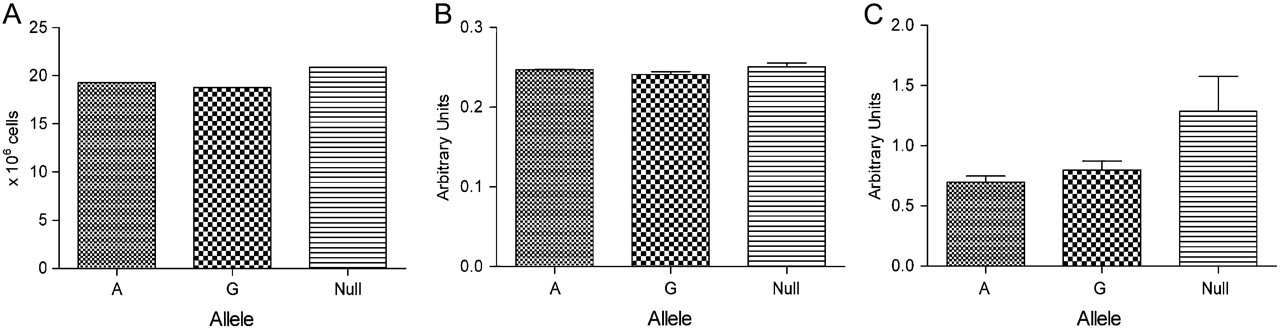
(A) Total cell count and (B) Bradford assay showing no significant differences in total cell number or total secreted protein levels in A and G alleles of matrix metalloproteinase-12 (MMP-12) rs652438 vector-transfected COS7 cells. (C) MTT assay indicating no significant differences in viable cells (n=3 for all experiments).
Cell migration
Transwells were coated with collagen IV, a non-fibrillar collagen expressed in basement membranes which can be degraded by MMP-12.15 Using t tests and mean values as a comparator, both MMP-12 allele transfected cells had increased migration compared to cells containing the null vector. Further to this, A allele containing cells had significantly increased migration (2.31-fold increase, p=0.0001) when compared to those transfected with the G allele (figure 5).

Chemotactic transwell migration assay (n=3) showing a 2.31-fold (p=0.0001) increase in migration for A allele transfected rs652438 COS7 cells compared with the G allele. Baseline used is the migration of null (N) vector-transfected COS7 cells. Cell migration photographs are representative of three independent experiments.
We investigated whether this effect on migration was relevant in patients with COPD. We examined data from the Nottingham cohort using BAL fluid samples in which the percentage composition values of inflammatory cells were available. Patients were genotyped for rs652438 using direct sequencing. Using a t test and comparing the means, we found a 1.28-fold (n=10, p=0.033) increase in the percentage of BAL fluid macrophages for A/A homozygotes compared with A/G genotypes (figure 6). There has also been evidence that MMP-12 is involved in neutrophil recruitment to the airways16 but in our study we did not find any significant differences as determined by t test. A/A genotypes had 4.820±2.239% composition of neutrophils while A/G genotypes had 21.040±9.058% composition (p=0.0602).
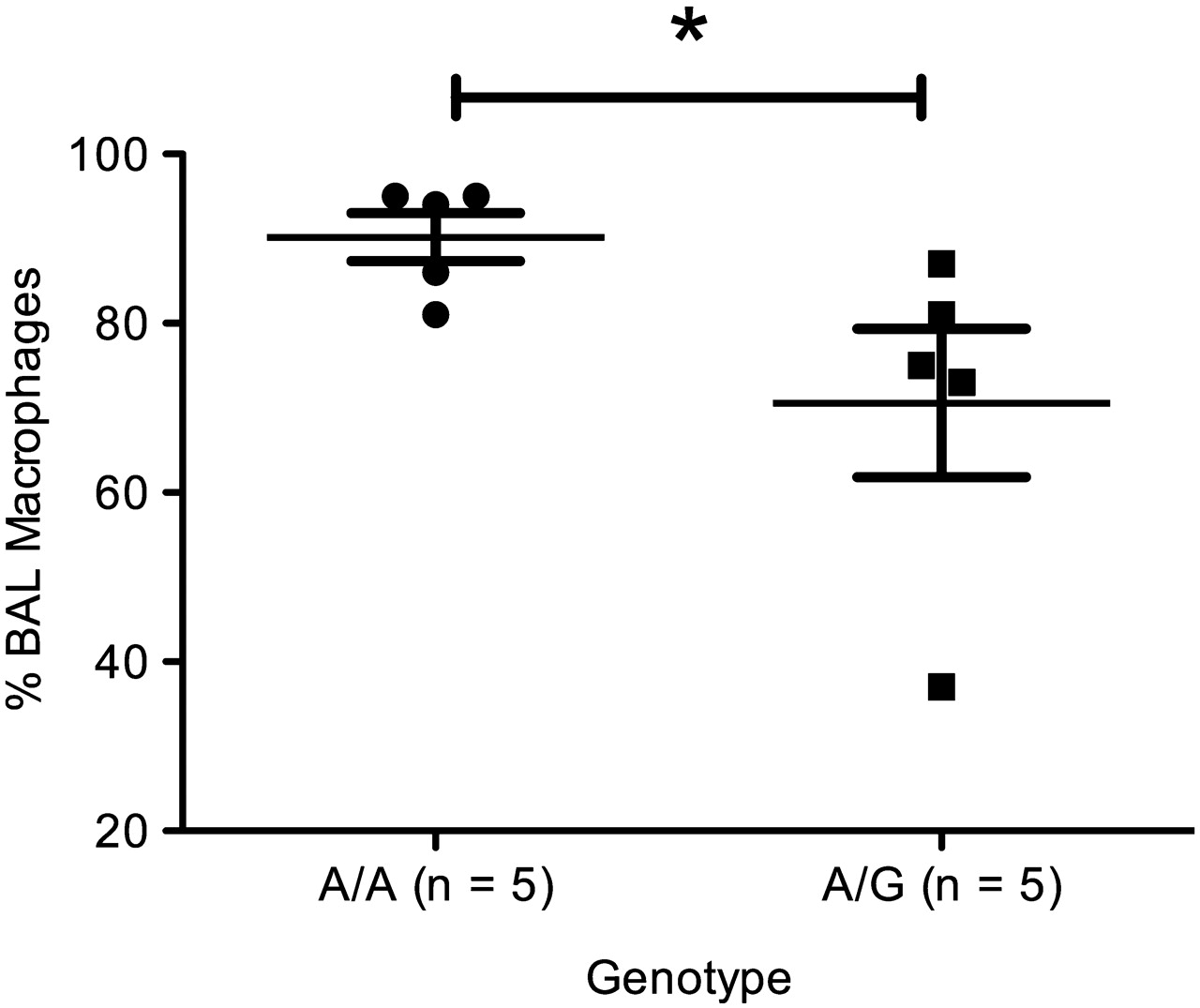
Percentage of macrophages in bronchoalveolar lavage (BAL) fluid from the Nottingham cohort. Individuals with A/A genotype had mean±SEM 90.20±2.85% macrophages while A/G individuals had 70.60±8.75% macrophages, a 1.18-fold increase (p=0.033).
In order to provide some replication, information from the ECLIPSE cohort was obtained regarding absolute inflammatory cell numbers according to genotype. As the rs652438 SNP is not present on the Illumina 550 genome-wide genotyping platform used in ECLIPSE, we studied the rs651159 intronic SNP which is in full linkage disequilibrium (r2=1) with rs652438 in HapMap CEU, EGP CEPH and AFD EUR submitter populations to the HapMap.17 Analysis of rs651159 allows 100% inference of rs652438 genotypes.
The focus of the analysis was the association of inferred rs652438 genotypes with absolute cell numbers of macrophages and neutrophils in induced sputum. In this large cohort, BAL fluid samples were not routinely taken and macrophage numbers were examined in induced sputum. Further to this, a comparable cell percentage phenotype in the induced sputum was not available. In the 232 patients for whom genotype and induced sputum data were available, there were 205 A/A homozygotes, 27 A/G heterozygotes but no G/G homozygotes.
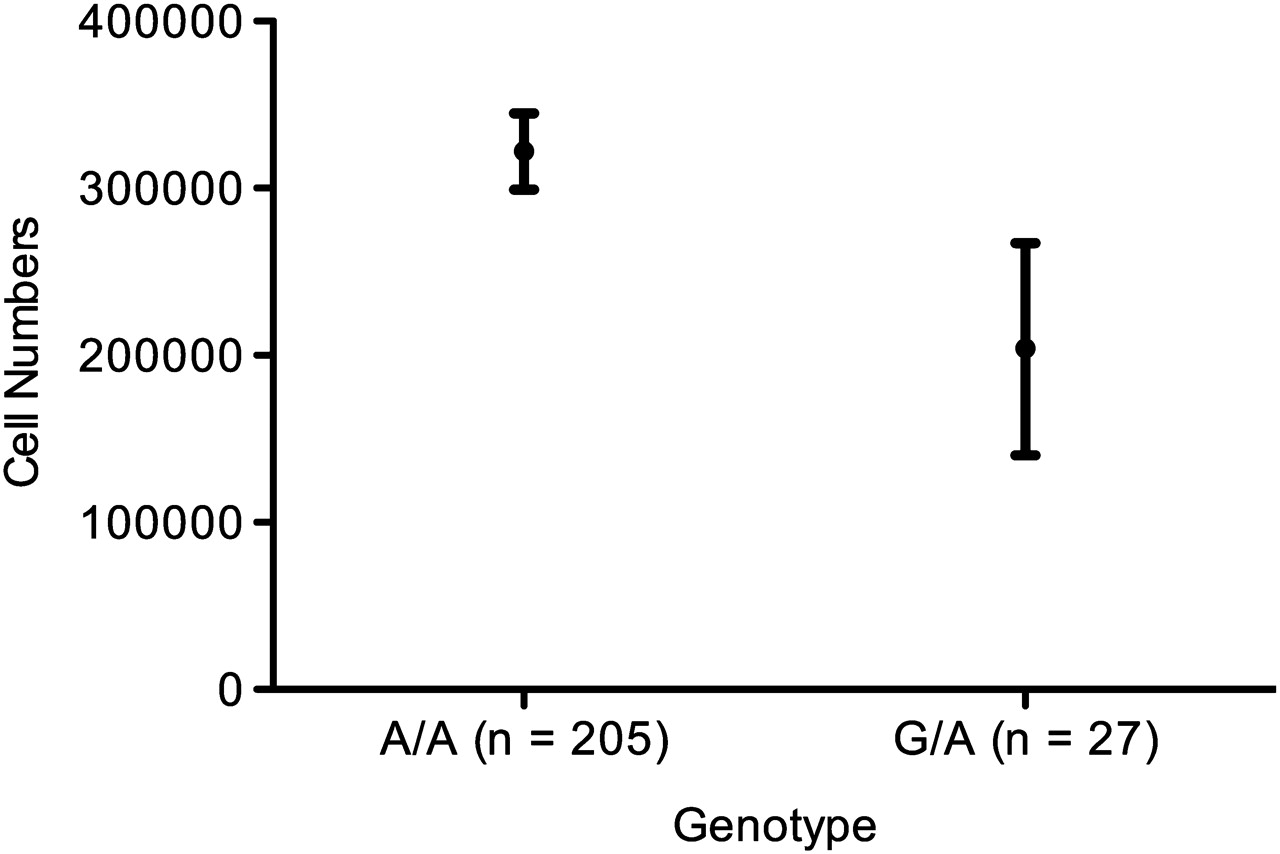
Absolute macrophage number in induced sputum was the dependent variable while genotype is the independent variable of interest. The final linear regression model was adjusted for smoking history, age, sex, pack-years and GOLD stage. Comparing the means showed a 1.58-fold increase in macrophage numbers in induced sputum samples of A/A homozygotes compared with A/G heterozygotes (figure 7), which did not reach statistical significance (p=0.083, adjusted r2=0.369) using a linear regression test.

Absolute macrophage number in induced sputum by genotype in the ECLIPSE cohort. A/A homozygotic individuals had a mean±SE of 3.22±0.23×105 cells per count compared with 2.04±0.64×105 cells per count for A/G heterozygotes, a 1.58-fold increase in cell number (p=0.083, adjusted r2=0.369).
The absolute neutrophil number was also investigated using linear regression adjusted for smoking history, age, sex, pack-years and GOLD stage No significant differences (p=0.306, adjusted r2=493) were found between the A/A group (mean±SE 2.48×106±0.25×106 cells per count) and the A/G group (mean±SE 3.25×106±0.71×106 cells per count).
Emphysema scores
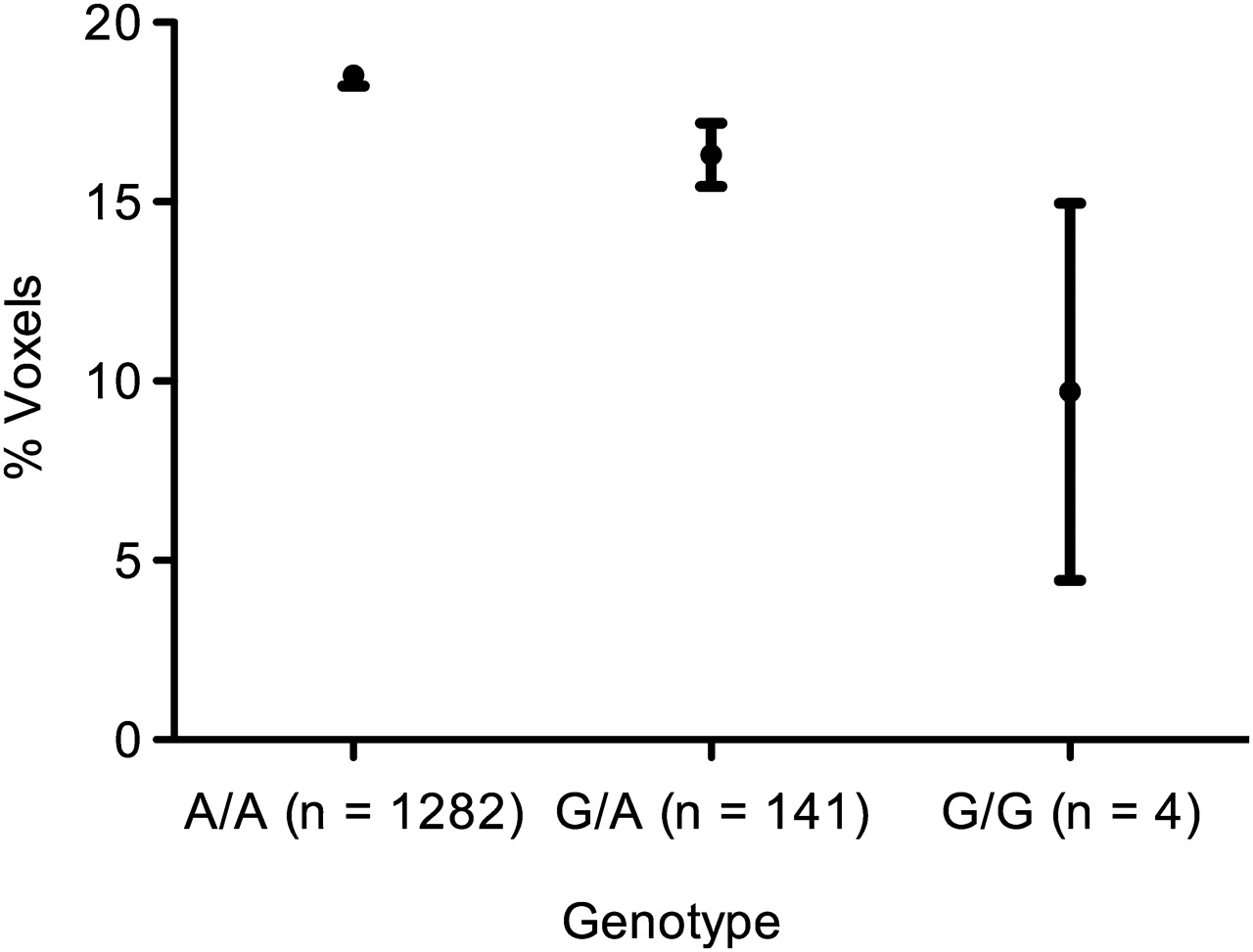
As in the previous investigation, the rs651159 SNP was used to infer the genotypes of rs652438 in ECLIPSE. There were 1282 A/A homozygotic individuals, 141 A/G heterozygotes and four G/G minor homozygotes. The emphysema score determined by CT scanning was used as a quantitative trait in association analysis against genotype. The final linear regression model was adjusted for smoking history, age, sex, pack-years and GOLD stage. There was a dose-dependent effect on emphysema with each G allele (p=0.016, adjusted r2=0.245), with a reduction in low attenuation areas between the A/A major and G/G minor homozygotes. Comparison of the means of the A/A and G/G homozygotes showed a 1.91-fold increase in the emphysema score (figure 8).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Emphysema score by genotype. The mean±SE values for the different genotypes were: A/A, 18.52±0.29; G/A, 16.31±0.89; G/G, 9.70±5.26. The increase between the major and minor homozygotes in emphysema scores is 1.91-fold (p=0.016, adjusted r2=0.245; n=1427).
Discussion
This study demonstrates that an exonic SNP alters the activity of MMP-12. Our findings provide evidence for the role of the rs652438 SNP in macrophage migration and emphysema severity, with the A allele over-represented in these cases. Consistent with these findings, we found that the A allele significantly increased the activity of MMP-12.
The MMP-12 protein exists in three major forms: an inactive 54 kDa form which has the propeptide domain cleaved to create the 45 kDa active form, and this active 45 kDa form is also processed to form a 22 kDa protein with the haemopexin domain removed. Although processed to the 22 kDa form, we do not think that this will have affected our in vitro findings as, in common with a previous study, we found that, in cell supernatants, the 45 kDa form is stable under storage at −80°C and for the duration of the assays.18 However, the purified recombinant MMP-12 readily underwent autocatalytic processing, suggesting the presence of stabilising factors in cell culture supernatants.
The haemopexin domain in MMP-12 has been hypothesised to have a flexible interaction with the catalytic domain compared with other MMPs, suggesting a role for the haemopexin domain in modulating enzymatic activity.19 The rs652438 SNP causes a neutral-polar mutation of arginine to serine, which suggests that the effect of the SNP seen in our study is through stabilisation of protein flexibility rather than a direct conformational change.
An interesting finding was the investigation of SNP alleles with regard to macrophage migration. The transwell migration assay set up to model macrophage invasion in the airways found significantly increased migration for the A allele. Furthermore, this finding was replicated in humans with statistically significant differences between individuals with A/A and A/G genotypes in the number/proportion of macrophages found in BAL fluid and induced sputum. However, it must be noted that grouped replication rather than direct replication is provided as the phenotypes available to us were not identical for the Nottingham and ECLIPSE populations. Furthermore, the rarity of the G/G genotype resulted in no individuals with this genotype in either dataset used to study inflammatory cells. These findings are consistent with cigarette-induced emphysema as they provide a correlation between the suggested macrophage recruitment role of MMP-12 in mice and humans.3 It is interesting that activity differences in the protease appear to translate to COPD-specific phenotypes. It would be worthwhile determining whether the SNP has a relationship with other COPD processes such as the levels of pathological agents secreted by alveolar macrophages or increased cleavage of α1-antitrypsin, a substrate for MMP-12.20
Another finding of this study was the association between the rs652438 SNP and severity of emphysema, with A/A homozygotes having more severe emphysema. Smokers with a less active form of MMP-12 may therefore be partially protected from severe emphysema. However, despite this being a single test of association, it persisted after adjusting for confounding factors (p=0.016). Thus, although these associations are consistent, further validation is required through replication in a larger cohort for macrophage infiltration and emphysema.
It has been known for some time that only a proportion of individuals who smoke develop COPD,21 and our findings contribute some genetic and functional evidence to the concept of the ‘susceptible smoker’. However, the rarity of the A/G and G/G genotypes at MMP-12 rs652438 (around 3.8% of individuals with severe or very COPD possess at least one G allele7) suggests that a number of other loci are likely to contribute to COPD susceptibility. It should also be noted that the ECLIPSE populations used in this study were smokers with COPD and thus would be predicted to have a lower frequency of the minor G allele compared with a ‘healthy’ smoker group.7 The rarity of these minor homozygotes suggests that, although it confers a protective effect against emphysema, it is negatively selected against. This could be due to the central role of MMP-12 in inflammatory processes and a possible protective role in cancer metastasis.22 Low MMP-12 activity in healthy individuals could thus be undesirable, resulting in susceptibility to various conditions.
MMP-12 has been implicated in a number of other diseases—specifically, the rs652438 SNP has been implicated in lung cancer metastasis and survival23 24—and our findings may also have implications for these diseases.
Perhaps the most important finding of this study is that it provides evidence that the findings in the MMP-12 knockout mouse model are relevant to the pathogenesis of human COPD. Knockout mice have impaired levels of tumour necrosis factor α (TNFα) release in response to acute smoke exposure.25 This is interesting as TNFα receptor knockout mice develop far less emphysema than control mice in response to smoke exposure.26 MMP-12 has also been shown to be able to cleave α1-antitrypsin.27 Furthermore, investigation of MMP-12 and associated pathways in COPD could lead to an intervention strategy appropriate to individuals with emphysema.
In conclusion, this study provides evidence that an exonic SNP can alter MMP-12 activity and this relates to certain aspects of the COPD phenotype. Together with previous research, this strengthens the case for the role of MMP-12 as a key protease in COPD. It is unusual to obtain correlations of genotype with phenotypic expression to the extent demonstrated in this study.
Acknowledgments
The authors would like to thank the members of the Evaluation of COPD Longitudinally to Identify Predictive Surrogate End-points (ECLIPSE) Steering Committee for providing data from this cohort. We are grateful to Lorna Magowan for technical assistance.
References
Supplementary materials
Web Only Data thx.2011.159087
Files in this Data Supplement:
Footnotes
NK and SRJ are joint senior authors.
Funding This study is funded by a Medical Research Council Capacity Building Studentship.
Competing interests None.
Ethics approval This study was conducted with the approval of the Nottingham Research Ethics Committee and all patients gave informed consent. Investigations into ECLIPSE resource were approved by the ECLIPSE steering committee. Informed consent and collection of the cohort was as described by Vestbo et al.11
Provenance and peer review Not commissioned; externally peer reviewed.