Article Text

Abstract
Background Persistence of myofibroblasts is believed to contribute to the development of fibrosis in idiopathic pulmonary fibrosis (IPF). Transforming growth factor β1 (TGFβ1) irreversibly converts fibroblasts into pathological myofibroblasts, which express smooth muscle α-actin (α-SMA) and produce extracellular matrix proteins, such as procollagen I (α1). Reactive oxygen species produced by NADPH oxidases (NOXs) have been shown to regulate cell differentiation. It was hypothesised that NOX could be expressed in parenchymal pulmonary fibroblasts and could mediate TGFβ1-stimulated conversion of fibroblasts into myofibroblasts.
Methods Fibroblasts were cultured from the lung of nine controls and eight patients with IPF. NOX4, α-SMA and procollagen I (α1) mRNA and protein expression, reactive oxygen species production and Smad2/3 phosphorylation were quantified, in the absence and in the presence of incubation with TGFβ1. Migration of platelet-derived growth factor (PDGF)-induced fibroblasts was also assessed.
Results It was found that (1) NOX4 mRNA and protein expression was upregulated in pulmonary fibroblasts from patients with IPF and correlated with mRNA expression of α-SMA and procollagen I (α1) mRNA; (2) TGFβ1 upregulated NOX4, α-SMA and procollagen I (α1) expression in control and IPF fibroblasts; (3) the change in α-SMA and procollagen I (α1) expression in response to TGFβ1 was inhibited by antioxidants and by a NOX4 small interfering RNA (siRNA); (4) NOX4 modulated α-SMA and procollagen I (α1) expression by controlling activation of Smad2/3; and (5) NOX4 modulated PDGF-induced fibroblast migration.
Conclusion NOX4 is critical for modulation of the pulmonary myofibroblast phenotype in IPF, probably by modulating the response to TGFβ1 and PDGF.
- Interstitial fibrosis
- oxidative stress
Statistics from Altmetric.com
Introduction
Idiopathic pulmonary fibrosis (IPF) is the most common idiopathic interstitial pneumonia. IPF is a disease with poor prognosis and an aggressive nature, and poses major challenges to clinicians.1 2 Currently, no effective treatments exist to stop ongoing fibrosis in IPF. Accumulation and persistence of myofibroblasts is believed to contribute to the development of fibrosis.3 α-Smooth muscle actin (α-SMA) expression, increased proliferative capacity, and increased generation and secretion of the extracellular matrix proteins such as collagen and fibronectin are key hallmarks of myofibroblast differentiation in fibrotic disorders.4 It has been established that myofibroblast differentiation occurs through combinatorial signals involving transforming growth factor β1 (TGFβ1) and integrin signalling.5
Several studies suggest that oxidant–antioxidant imbalances in the lower respiratory tract play a significant role in the pathogenesis of IPF. Reactive oxygen species (ROS) can promote a profibrogenic environment in the lung through the modulation of the protease–antiprotease balance.6 Also, ROS activate and/or mediate the effects of growth-regulatory cytokines, including TGFβ1.7 8 However, the endogenous sources of oxidants in IPF are poorly known.
The NADPH oxidase (NOX) proteins generate ROS, namely superoxide, by electron transfer to oxygen. Superoxide can be further converted to hydrogen peroxide by cellular superoxide dismutases. NADPH oxidases consist of membrane-associated and cytosolic subunits (for review, see Lambeth9). The most thoroughly investigated NADPH oxidase is the phagocytic gp91phox (NOX2)-containing subunit that interacts with a second membrane-bound subunit, p22phox, and several cytosolic and regulatory subunits including p47phox, p67phox and Rac. Both NOX1 and NOX4 are homologues of NOX2, and are expressed in different non-phagocytic cells.9 It has been recently shown that NOX1 and NOX4 are expressed in pulmonary arterial adventitial fibroblasts and that NOX4 contributes to the increase in ROS generation under hypoxic conditions, stimulating proliferation and inhibiting apoptosis of these cells.10 Furthermore, NOX4 is involved in TGFβ1-induced human cardiac fibroblast differentiation into myofibroblasts.11 Therefore, we hypothetised that NOX1 and NOX4 could also be expressed in parenchymal pulmonary fibroblasts and could mediate the effects of TGFβ1.
The aim of this study was to investigate NOX1 and NOX4 expression in pulmonary fibroblasts from control subjects and patients with IPF, and to analyse their respective role in mediating TGFβ1-induced fibroblast differentiation into myofibroblasts, a key phenomenon in IPF pathogenesis.12 The myofibroblast phenotype was analysed by examining α-SMA and procollagen I (α1) expression.
Materials and methods
See online supplement for further details.
Patients with lung fibrosis
Fibroblasts were derived from lung tissue sampled from eight patients with IPF. Lung samples were obtained by open lung biopsy (n=4) or at the time of lung transplantation (n=4). IPF was diagnosed according to the American Thoracic Society/European Respiratory Society (ATS/ERS) consensus criteria.13 Patients (six males; two females) had a median age of 50 years (range, 44–69 years). Six were ex-smokers, one never smoked and one was an active smoker. At the time of lung sampling, three patients were treated with low dose oral corticosteroids, associated with azathioprin in one patient.
Control patients
Fibroblasts were derived from lung samples from nine patients (six males, three females) undergoing lung surgery for removal of a primary lung tumour. Normal lung was obtained from a non-involved segment, remote from the solitary lesion. Median age was 54 years (28–68 years). Two patients had never smoked, four were ex-smokers and three were active smokers.
Culture of fibroblasts
Human lung fibroblasts were cultured from lung explants until passage five as previously described.14 In some experiments, cells were incubated in the presence of TGFβ1 (10 ng/ml, R&D Systems, Lille, France) or vehicle.
Cellular toxicity
Cellular toxicity and viability were assessed by trypan blue exclusion and lactate dehydrogenase (LDH) release in the medium, as described previously.15
Intracellular ROS production
Intracellular ROS production was quantified by measuring H2-DCFH-DA (2,7-dichlorofluorescein diacetate) oxidation, as described previously.15 16 Results are expressed in fluorescent arbitrary units.
Real-time reverse transcription–PCR (RT–PCR)
NOX1, 2, 3, 4 and 5, α-SMA and procollagen I (α1) mRNA expression was quantified by quantitative real-time RT–PCR by use of the PCR ABI 7700 apparatus (Applied Biosystems, Carlsbad, California, USA). Expression of mRNA was normalised to that of ubiquitin. In previous experiments, we verified that ubiquitin mRNA expression did not change under the different experimental conditions.
Cell transfection with small interfering RNA (siRNA)
To suppress endogenous NOX4 expression, experiments involved a specific siRNA (Xeragon-Qiagen, Valencia, California, USA,) against NOX4 designed on the target region from the NOX4 gene (5′-AAACCGGCAGGAGUUUACCCAG-3′). Scrambled siRNA (non-homologous to the human genome; 5′-AACCAGCAAGGUGUAUCGCCAC-3′) was used as a control.
Western blot analysis
Western blot was performed as described previously17 with use of the following antibodies: polyclonal antihuman NOX4 (1/1000, Novus Biologicals, Littleton, Colorado, USA), monoclonal anti-α-actin (1/3000, Sigma-Aldrich, Saint Quentin Fallavier, France) and monoclonal antihuman anti-Smad2 and 3 (1/500, R&D Systems, Lille, France). The expression of NOX4 was normalised in respect to the expression of β-actin. The expression of non-phosphorylated Smad2 (monoclonal antibody, 1/400, Cell Signalling, Saint Quentin Yvelines, France) was used to normalise the expression of phospho-Smad2/3.
Cell migration assay
Cell motility was examined as previously described18 using a modified Boyden chamber apparatus (Transwell Costar, Chorges, France) in response to 24 h stimulation with recombinant human platelet-derived growth factor-BB (rhPDGF-BB) 10 ng/ml (R&D systems) or medium alone. The chemotactic index was determined as the ratio of cells moving in response to a chemoattractant relative to cells moving in response to medium control.
Immunohistochemical analysis
Immunohistochemistry was performed as previously described.17 Cryostat sections (4–6 μm thick) were fixed in acetone and reacted with the primary antibody rabbit polyclonal anti-NOX4 (1/500 dilution, kindly provided by Professor Victor Thannickal, University of Alabama at Birmingham, Birmingham, Alabama, USA).
Statistical analysis
See online supplement for further details.
All the data are displayed as medians, IQR and minimum and maximum values. All of the analysis were performed by non-parametric tests. A p value <0.05 was considered as statistically significant.
Results
Expression of NOX mRNA and production of oxidants in pulmonary fibroblasts
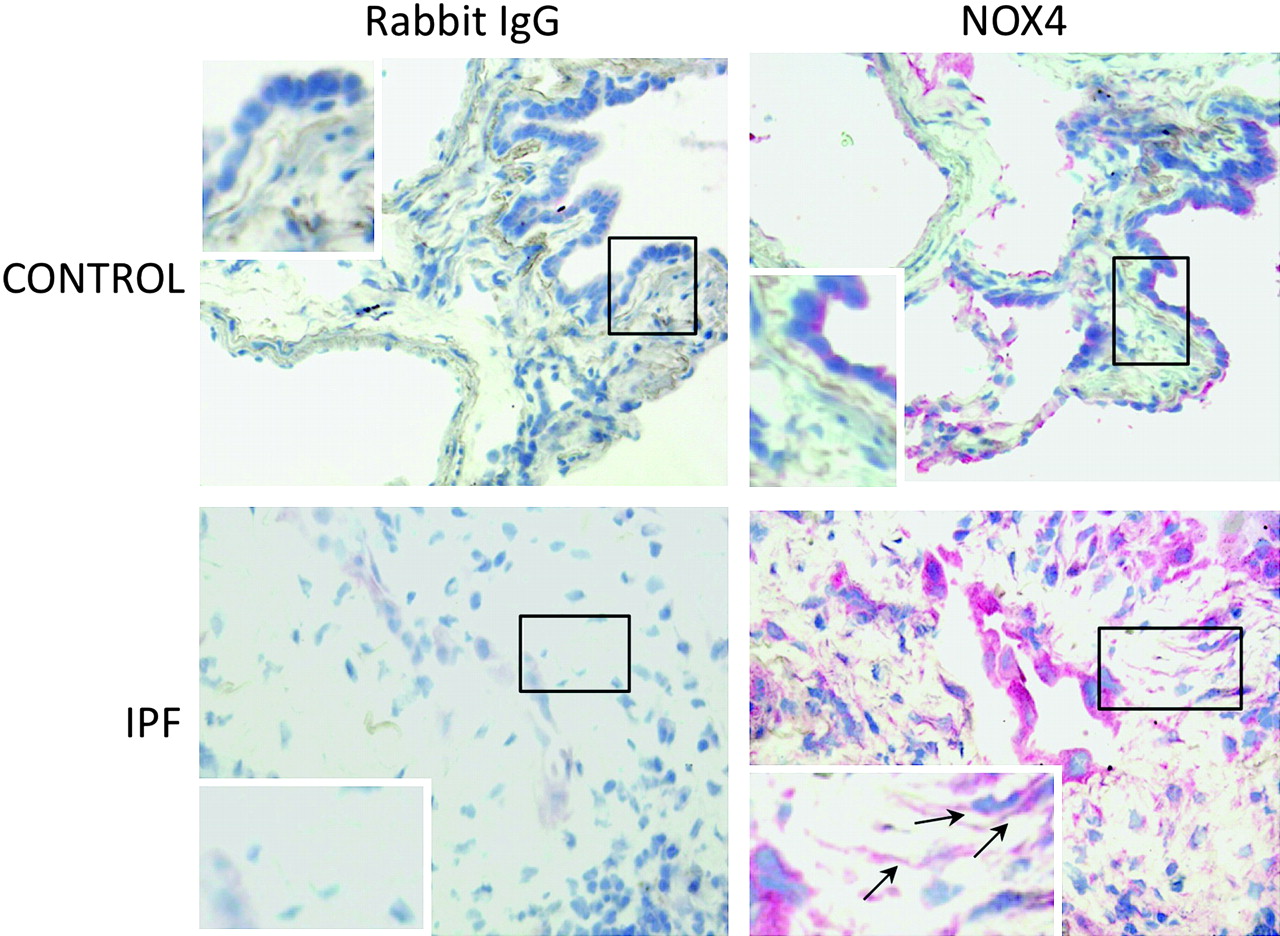
Lung fibroblasts from control patients expressed NOX1, 2, 4 and 5 mRNA to similar degrees, whereas expression of NOX3 was not detected in these cells (figure 1A–D). When comparing IPF and control fibroblasts, NOX4 was the only NOX whose mRNA expression was increased in the former as compared with the latter condition (p=0.043). The increased NOX4 expression in IPF fibroblasts was confirmed at the protein level by western blot (p=0.025 control vs IPF, figure 1E). NOX4 mRNA and protein induction in IPF fibroblasts was accompanied by an increased intracellular ROS production in these cells as compared with control fibroblasts (p=0.028, figure 1F). Immunohistochemistry performed on control and IPF lung samples demonstrated that lung fibroblasts expressed immunoreactive NOX4 in situ in the fibrotic lung (figure 2). Bronchial and alveolar epithelial cells and pulmonary endothelial cells also expressed immunoreactive NOX4.
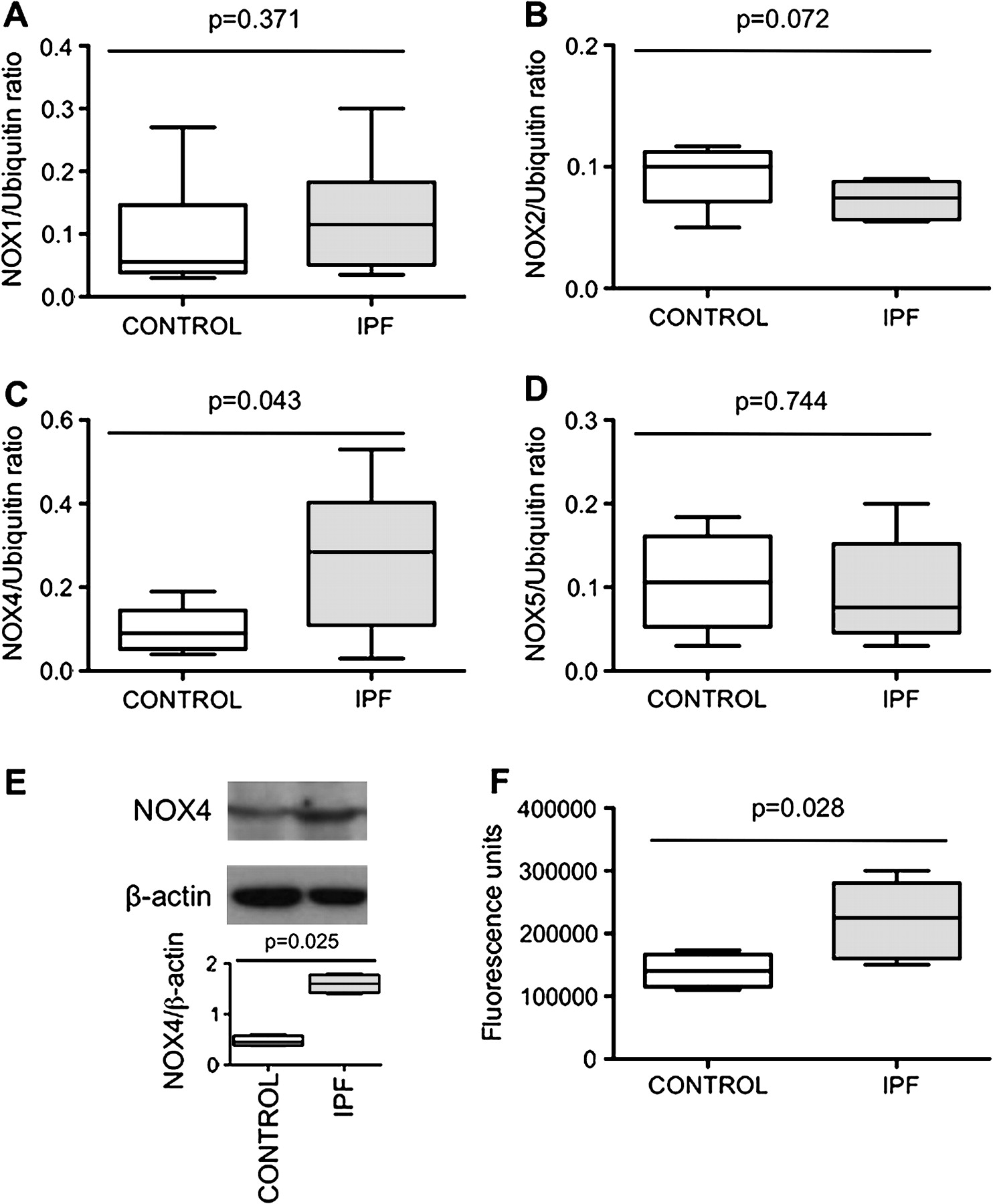
(A–D) NOX1, 2, 4 and 5 mRNA expression in lung fibroblasts from patients with idiopathic pulmonary fibrosis (IPF) compared with controls expressed as the ratio to ubiquitin mRNA levels. Data are presented as a box and whisker plot with median, IQR and minimum and maximum values. (E) Typical western blot and quantification of NOX4 protein expression (67 kDa) in lung fibroblasts from patients with IPF and controls. NOX4 is expressed as the ratio to β-actin expression. Data are presented as a box and whisker plot with median, IQR and minimum and maximum values. (F) Reactive oxygen species production by IPF and control fibroblasts assessed by measurement of H2-DCFH-DA (2,7-dichlorofluorescein diacetate) oxidation. The values, expressed as arbitrary fluorescence units, are presented as a box and whisker plot with median, IQR and minimum and maximum values.
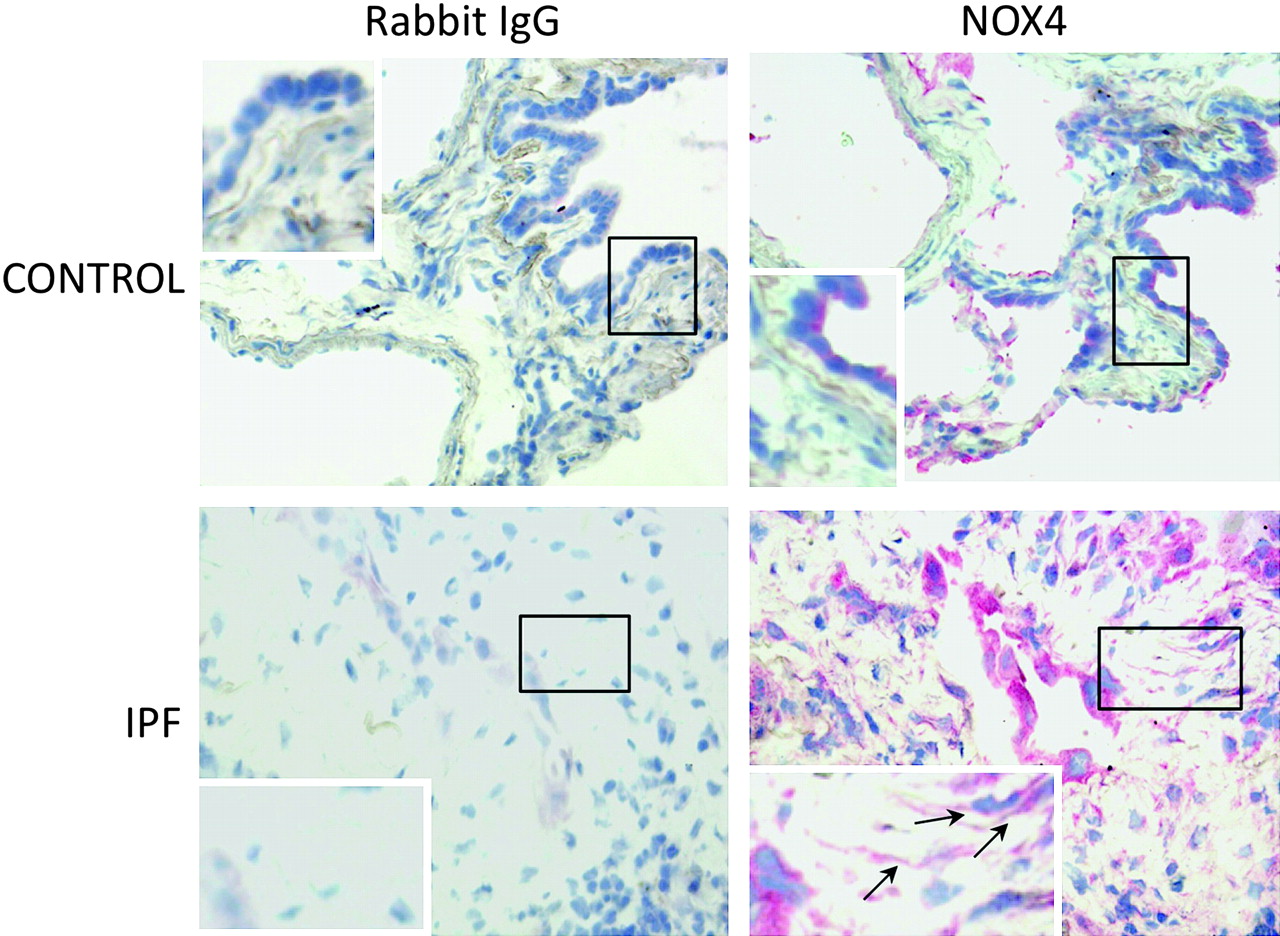
Detection of immunoreactive NOX4 in the lung. Immunohistochemistry shows that bronchial and alveolar epithelial cells express NOX4 in the normal lung. In idiopathic pulmonary fibrosis (IPF) lung samples, hyperplastic alveolar cells and fibroblasts (arrows) are strongly labelled. The control antibody always gave a negative signal.
Expression of markers of myofibroblast diferentiation
The content of α-SMA mRNA was increased in IPF fibroblasts as compared with controls (p=0.045; figure E1 in the online supplement) whereas procollagen I (α1) mRNA content was similar in both groups (p=0.310; figure E1 in the online supplement). Analysis of fibroblasts from patients with IPF showed a significant correlation between α-SMA or procollagen I (α1) mRNA and NOX4 mRNA expression (Spearman rho=0.994, p<0.0001 in both cases; figure E1 in the online supplement), suggesting a role for this NADPH oxidase homologue in myofibroblast differentiation.
Effect of TGFβ1
Since NOX4 was overexpressed in IPF fibroblasts and correlated with markers of fibroblast differentiation into myofibroblast, and since TGFβ1 is an essential inducer of myofibroblast differentiation,5 we analysed if TGFβ1 modulated NOX4 expression by lung fibroblasts. Incubation of control fibroblasts with TGFβ1 for 18 h induced a threefold increase of NOX4 mRNA content (figure 3A, p=0.014). In IPF fibroblasts, TGFβ1 induced an eightfold increase of NOX4 mRNA content (figure 3B, p=0.008). As expected, TGFβ1 increased α-SMA and procollagen I (α1) mRNA expression in control and IPF fibroblasts (figure 3A,B, p=0.009, p=0.008, p=0.034 and p=0.029 for α-SMA in control and IPF fibroblasts and for procollagen I (α1) in control and IPF fibroblasts, respectively). The increase in α-SMA was also observed at the protein level (figure 3C). Incubation of control fibroblasts with TGFβ1 induced a significant increase in ROS production, measured by oxidation of DCFH (figure 3D). This increase was more important in IPF than in control cells at 1, 3 and 24 h post-TGFβ1 (p=0.033, p=0.039 and p=0.028 at each time, respectively, figure 3D).

(A and B) Effect of transforming growth factor β1 (TGFβ1) in the presence or in the absence of N-acetylcysteine (NAC, 1 mM) or diphenylene iodonium (DPI, 10 μm) on NOX4, α-smooth muscle actin (α-SMA) and procollagen I (α1) mRNA expression in lung fibroblasts from controls and patients with idiopathic pulmonary fibrosis (IPF). Data are presented as a box and whisker plot with median, IQR and minimum and maximum values. (C) Typical western blot and quantification of α-SMA protein expression in lung fibroblasts after TGFβ1 incubation in the presence or in the absence of NAC (1 mM) or DPI (10 μm). α-SMA is expressed as the ratio to β-actin expression. Data are presented as a box and whisker plot with median, IQR and minimum and maximum values. (D) TGFβ1 stimulates reactive oxygen species (ROS) production in human lung fibroblasts. Control and IPF fibroblasts were stimulated with TGFβ1 for 1–24 h and ROS production was assessed by measurement of H2-DCFH-DA (2,7-dichlorofluorescein diacetate) oxidation. The four figures show production by the two groups of cells at 1, 3, 12 and 24 h post-TGFβ1, respectively. Values are presented as a box and whisker plot with median, IQR and minimum and maximum values.
Role of NOX4 in TGFβ1-induced myofibroblast differentiation
We first examined if ROS, the product of NOX4 activity, were involved in TGFβ1-induced myofibroblast differentiation. First, we examined the effect of the antioxidant N-acetylcysteine (NAC). Pretreatment of control and IPF fibroblasts for 1 h with NAC (1 mM) totally prevented the increase of α-SMA and procollagen I (α1) mRNA expression induced by TGFβ1 (figure 3A–C). This result strongly suggested that the oxidant–antioxidant imbalance is involved in the effect of TGFβ1.
In order to examine the involvement of NOX4 in TGFβ1 effects, we first incubated the fibroblasts for 1 h with diphenylene iodonium (DPI, 10 μM), an inhibitor of flavoprotein-dependent oxidases including NOX.19 DPI prevented TGFβ1-induced α-SMA and procollagen I (α1) mRNA expression in both control and IPF fibroblasts (figure 3A–C).
Secondly, we examined the effects of cell transfection with a NOX4 siRNA. NOX4 siRNA significantly reduced NOX4 mRNA levels at 24 h in both control and IPF fibroblasts, whereas no such effect was observed with lipofectamine or scrambled siRNA (figure 4A,B).

(A and B) Effect of cell transfection with a small interfering RNA (siRNA) anti-NOX4 on NOX4, α-smooth muscle actin (α-SMA) and procollagen I (α1) mRNA expression induced by transforming growth factor β1 (TGFβ1) in lung fibroblasts from controls and patients with idiopathic pulmonary fibrosis (IPF). SiScr, scrambled siRNA. NOX4, α-SMA and procollagen I (α1) mRNA levels are expressed as the ratio to ubiquitin mRNA levels. Values are presented as a box and whisker plot with median, IQR and minimum and maximum values.
Transfection with NOX4 siRNA significantly inhibited the effect of TGFβ1 on myofibroblast differentiation in both control and IPF cells (figure 4A,B). Indeed, expression of α-SMA and procollagen I (α1) was reduced by 67% and 68% and by 64% and 54% in control and IPF fibroblasts, respectively. These results demonstrate that NOX4 mediates the effect of TGFβ1 on myofibroblast differentiation.
It has to be noted that both NAC and DPI reversed NOX4 mRNA induction by TGFβ1, showing probably an autocrine role for NOX4-derived ROS in NOX4 induction by TGFβ1.
None of the different pharmacological treatments and transfection procedures altered cell viability (data not shown).
Role of NOX4 in TGFβ1-induced Smad2 and 3 phosphorylation
It has been shown that Smads mediate TGFβ1 induction of α-SMA in human lung fibroblasts.20 21 Since phosphorylation of Smad2/3 can be modulated by ROS,11 we analysed whether NOX4 was involved in Smad2/3 phosphorylation induced by TGFβ1. The results of these experiments show that 1 h cell pretreatment with both NAC and DPI (1 mM and 10 μM, respectively) prevented Smad2/3 phosphorylation induced by 18 h incubation with TGFβ1 in control and IPF fibroblasts (figure 5A,B). Cell transfection with the NOX4 siRNA gave similar results (figure 5A,B). These results demonstrate that NOX4-produced ROS are involved in Smad2/3 phosphorylation by TGFβ1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Typical western blot showing the expression of phosphorylated (p-Smad2/3) and non-phosphorylated Smad2/3 in human lung fibroblasts from controls and patients with idiopathic pulmonary fibrosis (IPF) after incubation with transforming growth factor β1 (TGFβ1) in the presence or in the absence of a scrambled small interfering RNA (siScr), a siRNA anti-NOX4 (siNOX4), diphenylene iodonium (DPI, 10 μm) or N-acetylcysteine (NAC, 1 mM). A Box and whisker plot with median, IQR and minimum and maximum values shows quantification of p-Smad2/3 expressed as the ratio of Smad2/3 expression.
Role of NOX4 in PDGF-induced fibroblast migration
Finally, we analysed if NOX4 was involved in fibroblast migration induced by PDGF-BB, a strong migration stimulus. First, we examined if PDGF modulated NOX4 expression by lung fibroblasts. Incubation of control fibroblasts and IPF fibroblasts with PDGF for 24 h (10 ng/ml) induced a 1.5- and 2.5-fold increase of NOX4 mRNA content, respectively (figure E2 in the online supplement, p=0.028 and p=0.002, respectively).
Having demonstrated that NOX4 was upregulated by PDGF, we analysed its role in cell migration by incubating cells with NAC or DPI. Both NAC and DPI suppressed PDGF-induced fibroblasts migration in control and IPF fibroblasts, showing a role for NOX4 in this process (figure E2 in the online supplement and data not shown).
Discussion
The main results of the present study are that (1) NOX4 mRNA expression was upregulated in pulmonary fibroblasts from patients with IPF and correlated with mRNA expression of α-SMA and procollagen I (α1) mRNA; (2) TGFβ1 upregulates NOX4, α-SMA and procollagen I (α1) mRNA and α-SMA protein expression in control and IPF human pulmonary fibroblasts; (3) the change in α-SMA and procollagen I (α1) expression in response to TGFβ1 requires NOX4-derived ROS; (4) NOX4 modulates α-SMA and procollagen I (α1) expression by controlling activation of Smad2/3; and (5) NOX4 modulates fibroblasts migration induced by PDGF-BB. These new findings support the notion that NOX4 is critical for modulation of pulmonary myofibroblast phenotype in IPF, probably by modulating the response to TGFβ1 and PDGF. Similar results concerning the response to TGFβ1 were very recently reported by Hecker and colleagues22 in an article published after the present study was submitted for publication.
Expression of NOX4 in control fibroblasts
Expression of NOX4 protein was detected in lungs from both control subjects and patients with idiopathic pulmonary artery hypertension; the expression was predominantly in the medial layer of pulmonary arteries and was higher in patients than in controls.10 Accordingly, pulmonary artery adventitial fibroblasts in culture also expressed both NOX1 and NOX4.23 In agreement with these findings, in the present study we found that parenchymal pulmonary fibroblasts from controls and patients with IPF expressed both NOX1 and NOX4. This result is different from data reported in cardiac fibroblasts in which NOX1 expression was barely detectable,11 stressing the cell specificity of NOX expression. To the best of our knowledge this is the first demonstration that non-vascular pulmonary fibroblasts express NOX. It is noteworthy that our experiments were performed on cells in the fifth passage, since progressive downregulation of NOX4 expression has been correlated with increasing passage number.24 However, our immunohistochemical results demonstrate that fibroblasts do express immunoreactive NOX4 in situ in the fibrotic lung.
NOX4 upregulation in IPF fibroblasts
The mechanism(s) involved in NOX4 upregulation in IPF fibroblasts are difficult to analyse since very few data concerning the regulation of NOX4 activity and expression are available. Recent data from recombinant NOX4 expression suggest that NOX4 enzymatic activity depends on the membrane-associated p22phox subunit, whereas cytosolic subunits, phosphorylation or interaction with Rac are apparently not required for its activation.25 26 Concerning the regulation of gene expression, and the relevant transcription factors involved, very few data are available since promoter studies of NOX4 are lacking. Our data clearly show that TGFβ1 increases NOX4 expression both in control and in IPF fibroblasts, in accordance with previous data in cardiac fibroblasts and airway smooth muscle.11 27 28 Interestingly, the increase was stronger in IPF fibroblasts than in controls. One can speculate that TGFβ1 can be responsible for NOX4 upregulation in IPF fibroblasts ex vivo via an autocrine loop, since expression of TGFβ1 has been shown to be increased in these cells.29 Furthermore, high levels of TGFβ1 produced by epithelial cells could contribute to upregulation in fibroblasts in situ in the lung of patients with IPF.30
NOX4 induction by TGFβ1 was inhibited by NAC and DPI, suggesting that NOX-produced ROS are implicated in NOX4 gene expression regulation. We obtained similar results in human alveolar bronchiolar and epithelial cell lines,15 but the molecular mechanisms involved are unknown. Clearly, more studies are needed to understand NOX4 gene expression regulation in IPF fibroblasts.
Role of NOX4 in myofibroblast differentiation in IPF
Whatever the mechanism(s) involved in gene expression regulation, NOX4 was involved in myofibroblast differentiation in IPF. This conclusion is supported by (1) the correlation between basal NOX4 and α-SMA and procollagen I (α1) mRNA expression; and (2) the prevention of TGFβ1 differentiating effects by the NOX4 siRNA. This last phenomenon was also observed in cardiac fibroblasts.11 It has to be noted that we cannot ensure that NOX4-produced ROS were involved in myofibroblat differentiation in the present study since we did not measure fibroblast ROS production in NOX4 siRNA-treated cells. However, several pieces of experimental evidence strongly suggest that the effects we observed were mediated by ROS produced by NOX4. First, NOX4 was the only NOX homologue upregulated in IPF fibroblasts; secondly, ROS production was higher in these cells as compared with control cells; and, thirdly, similar results in terms of α-SMA and procollagen I (α1) expression were obtained by incubating these cell with the antioxidant NAC and by transfecting them with a NOX4 siRNA. Furthermore, a recent study with controlled recombinant expression of NOX4 demonstrated a strong correlation between NOX4 mRNA level and ROS generation.31 Thus, in this experimental setting, the activity of NOX4 was demonstrated to depend directly on its gene expression levels. Moreover, we and others have previously shown that siRNA against NOX4 significantly reduced the basal ROS level in rat smooth muscle cells and in pulmonary epithelial cell lines.15 32 Collectively, these data strongly support a role for NOX4-produced ROS in myofibroblast differentiation.
Interestingly, immunohistochemistry showed a strong expression of NOX4 by hyperplastic alveolar epithelium in the fibrotic lung (figure 2). In the kidney, NOX4 has been shown to be involved in the regulation of epithelial–mesenchymal transition.33 Although not specifically studied, this phenomenon could also occur in the lung and implicate NOX4 in the fibrogenic process through multiple pathways.
Role of NOX4 in TGFβ1-induced myofibroblast differentiation and PDGF-induced migration
TGFβ1 stimulates differentiation of fibroblasts into myofibroblasts using Smad2/3 signalling.21 After initial phosphorylation, Smad2/3 activation in response to TGFβ1 is prolonged (hours to days depending on the cell type). Interestingly, we showed that Smad2/3 phosphorylation was upregulated by TGFβ1 after 18 h of incubation. The antioxidant NAC, the NOX inhibitor DPI and the siRNA anti-NOX4 led to a marked inhibition of Smad2/3 phosphorylation, as demonstrated previously in human cardiac fibroblasts.11 These data imply that oxidants produced by NOX4 mediate α-SMA and procollagen I (α1) upregulation in pulmonary fibroblasts in response to TGFβ1, probably by stimulating phosphorylation and activation of Smad2/3. In addition, NOX4 mediates PDGF-induced fibroblast migration.
In summary, we have shown NOX4 upregulation in pulmonary fibroblasts from patients with IPF, and identified a novel role for NOX4 as an essential mediator of Smad2/3 transcription factor activation in response to TGFβ1 in pulmonary fibroblasts. Because of the critical role of TGFβ1-induced myofibroblast activation in IPF, these data provide insights into novel mechanisms with potential therapeutic implications for IPF.
Acknowledgments
We acknowledge Françoise Zerah (Assistance Publique-Hôpitaux de Paris, Service d'Explorations Fonctionnelles, Hôpital Henri Mondor, Créteil, France) for statistical advice.
References
Supplementary materials
Web Only Data thx.2009.113456
Files in this Data Supplement:
Footnotes
BC and JB shared senior authorship.
Funding NA was supported by Chancellerie des Universités de Paris (legs Poix), and JB by INSERM and Assistance Publique-Hôpitaux de Paris (Contrat d'Interface). This work was supported by the European Commission (FP 7, European IPF Network) and by the Agence Nationale de la Recherche (ANR Physio 2006, FIBROPNEUMO).
Competing interests None.
Patient consent Obtained.
Ethics approval This study was approved by the local ethics committee of Saint Germain en Laye Nospital and stored biopsies were reported to our institutional board (Délégation à la Recherche Clinique, Assistance Publique-Hôpitaux de Paris, Carré Historique de l'Hôpital Saint- Louis, Paris, France).
Provenance and peer review Not commissioned; externally peer reviewed.