Article Text

Abstract
There has been much recent interest in the role of the vitamin D axis in lung disease, which includes vitamin D, vitamin D receptor (VDR) and vitamin D-binding protein (VDBP; also known as Gc-globulin). VDBP is a serum protein which has immunomodulatory functions relevant in the lung, predominantly relating to macrophage activation and neutrophil chemotaxis. Variations within its gene are also associated with airways disease, implying a role for the protein product in pathogenesis. Thus far the majority of evidence relates to chronic obstructive pulmonary disease (COPD), but is scant in other airways diseases, such as asthma and bronchiectasis. VDBP also acts as a scavenger protein to clear extracellular G-actin released from necrotic cells, which may be of relevance in severe lung infections and acute lung injury. Vitamin D protects against the development of cancer and tuberculosis, although optimal levels are unknown. The majority of circulating vitamin D is bound to VDBP, and its uptake into cells occurs in both bound and unbound forms, which suggests the role of VDBP warrants further study in these conditions as well. This article reviews the evidence of the role VDBP and its gene (GC) in a range of lung diseases, including asthma, COPD and tuberculosis.
- COPD mechanisms
- lung cancer
- macrophage biology
- respiratory infection
- tuberculosis
Statistics from Altmetric.com
Introduction
The potential role of vitamin D in asthma,1 chronic obstructive pulmonary disease (COPD)2 and tuberculosis (TB)3 has recently been highlighted in a series of editorials, and is summarised in table 1. The majority of vitamin D in the circulation is bound to vitamin D-binding protein (VDBP, also known as Gc-globulin), which has anti-inflammatory and immunomodulatory functions independent of vitamin D carriage.4 5 These roles, together with their relationship to serum vitamin D (25-hydroxyvitamin D (25-OHD)) levels, may therefore be important in a range of lung diseases.
Vitamin D and lung disease
The human Gc-globulin was originally identified by Hirschfeld in 1959 as a marker in the λ-globulin of the human serum by serum electrophoresis.6 Initially it was characterised as a group-specific component or Gc-globulin, but its identity as the plasma protein that binds vitamin D was not discovered until 1975,7 when it gained its current name, vitamin D-binding protein (VDBP). VDBP was then known to transport 25-OHD, the major circulating form of vitamin D, and 1,25-dihydroxyvitamin D (1,25-OHD), the most active vitamin D metabolite, but over the years it has been appreciated that it has other roles beyond effects of the balance between these two forms on bone metabolism. VDBP is expressed in many tissues, including liver, kidney, gonads and fat.8 It is also expressed by neutrophils,9 10 contributes to macrophage activation,4 augments monocyte and neutrophil chemotaxis to C5-derived peptides11 12 and acts as a scavenger protein to clear extracellular G-actin released from necrotic cells.13–15 Any or all of these functions are likely to be relevant in the lung.
The human VDBP gene
The human vitamin D-binding protein gene (GC) is part of a gene cluster that includes the albumin (ALB) and α-fetoprotein (AFP) genes (termed the albumin multigene family) and, like the other genes in the cluster, is robustly expressed in the liver and other tissues.8 GC is located on chromosome 4, is ∼42 kb in size, and is comprised of 13 exons,16 one of which is entirely untranslated, and two others only partially translated.17 Transcription of the gene is regulated by a liver-enriched transcription factor at binding sites close to GC on chromosome 4.18 The location of the gene cluster, relative gene positions and the structure of GC are shown in supplementary figure 1 online. Some genetic terminology is also defined in the supplementary material.
GC polymorphisms
GC is highly polymorphic, with three commonly recognised variants (GC1F, GC1S and GC2) and >120 rarer variants.19 Single nucleotide polymorphisms (SNPs; rs4588 and rs7041) in exon 11 of the gene result in the common isoforms, termed GC1 and GC2; GC1 is then subdivided into GC1F and GC1S. Their protein products differ at positions 416 and 420, such that GC1F and GC1S proteins are identical except for a substitution of glutamic acid for aspartic acid in GC1S at position 416, whilst GC2 differs from GC1F by having a further substitution of lysine for threonine at position 42020 (table 2). These polymorphisms affect protein function, such that GC2 is less able to be converted to macrophage-activating factor,4 resulting in reduced macrophage function in GC2 carriers.
Haplotypes of rs7041 and rs4588 result in structurally different proteins
There is also significant variation in the ability of these common variants to bind vitamin D, perhaps driven by the rs7041 genotype,21 which may have an impact on serum 25-OHD levels. This may partly explain racial and geographical differences in GC allele frequencies,22 such that those with higher vitamin D binding affinity (such as GC1F) are more common in darker skinned races, and highly sun-exposed regions. Serum levels of both 25-OHD and 1,25-OHD vary according to GC genotype, even within racial groups,23 and this, or variations in the other protein functions, may account for associations of the GC genotype with a variety of diseases, including those discussed here. In addition, the response to vitamin D supplementation may relate to GC genotype.24 Unlike other VDBP functions, there are no known variations in actin binding capacity as a result of genetic variation.20 The role of VDBP in macrophage activation and neutrophil chemotaxis led to studies of genetic associations of GC in lung diseases where such processes might be important in pathogenesis. These will be discussed in disease-specific sections later.
Structure and function of VDBP
VDBP is a glycosylated α-globulin, ∼58 kDa in size. It is 458 amino acid residues in length and folds into a disulfide-bonded, triple-domain structure, further divided into two repeated, homologous domains of 186 amino acids (domains I and II) and a shorter domain of 86 residues at the C-terminus (domain III).25 Its structure is shown in supplementary figure 2 online. In simplified terms, domain I binds vitamin D25 whilst it is predominantly domain III that binds actin,26 the functions which are independent of each other.
Circulating vitamin D and its metabolites are bound to both albumin and VDBP. Since the affinity of albumin for 25-OHD and 1,25-OHD is substantially lower than that of VDBP27 the vast majority are bound to VDBP, but because of its relative abundance a proportion remain bound to albumin. Since the serum levels of both carrier proteins exceed those of 25-OHD and 1,25-OHD, the vast majority of these carrier proteins are empty.28 The relationship of VDBP and vitamin D concentrations is not yet clear. Low total vitamin D concentrations were reported in the original VDBP knockout mouse model, but did not relate to that of the active form of vitamin D (1,25-OHD) in more recent work,29 although these concepts have not been studied widely in man.
Vitamin D may be taken up into cells by diffusion of unbound vitamin across cell membranes, and by endocytosis of that bound to VDBP via binding of VDBP to megalin30 and cubulin,31 a process illustrated in figure 1. This was first recognised in renal cells, but has now been reported in other tissues, including macrophages.28 These facts have a bearing on reports relating vitamin D to lung disease, as most assays measure both bound and unbound forms. It is also important to realise that 25-OHD levels must be very low (<10 ng/ml) before the concentration of 1,25-OHD is affected, due to its regulation by the calcium homeostasis systems.33 The serum level of 25-OHD considered normal (or sufficient) is a topic of debate at present, with some reference laboratories known to us using a level of 30 ng/ml, and others 20 ng/ml. Since most of the immunomodulatory function of vitamin D relates to 1,25-OHD34 it may be that only patients exhibiting severe vitamin D deficiency (<10 ng/ml) will have any alterations in immune function, since it is only this group in whom 1,25-OHD will also be low. This is likely to be relevant in studies of lung diseases, such as TB, where immune modulation is biologically important.
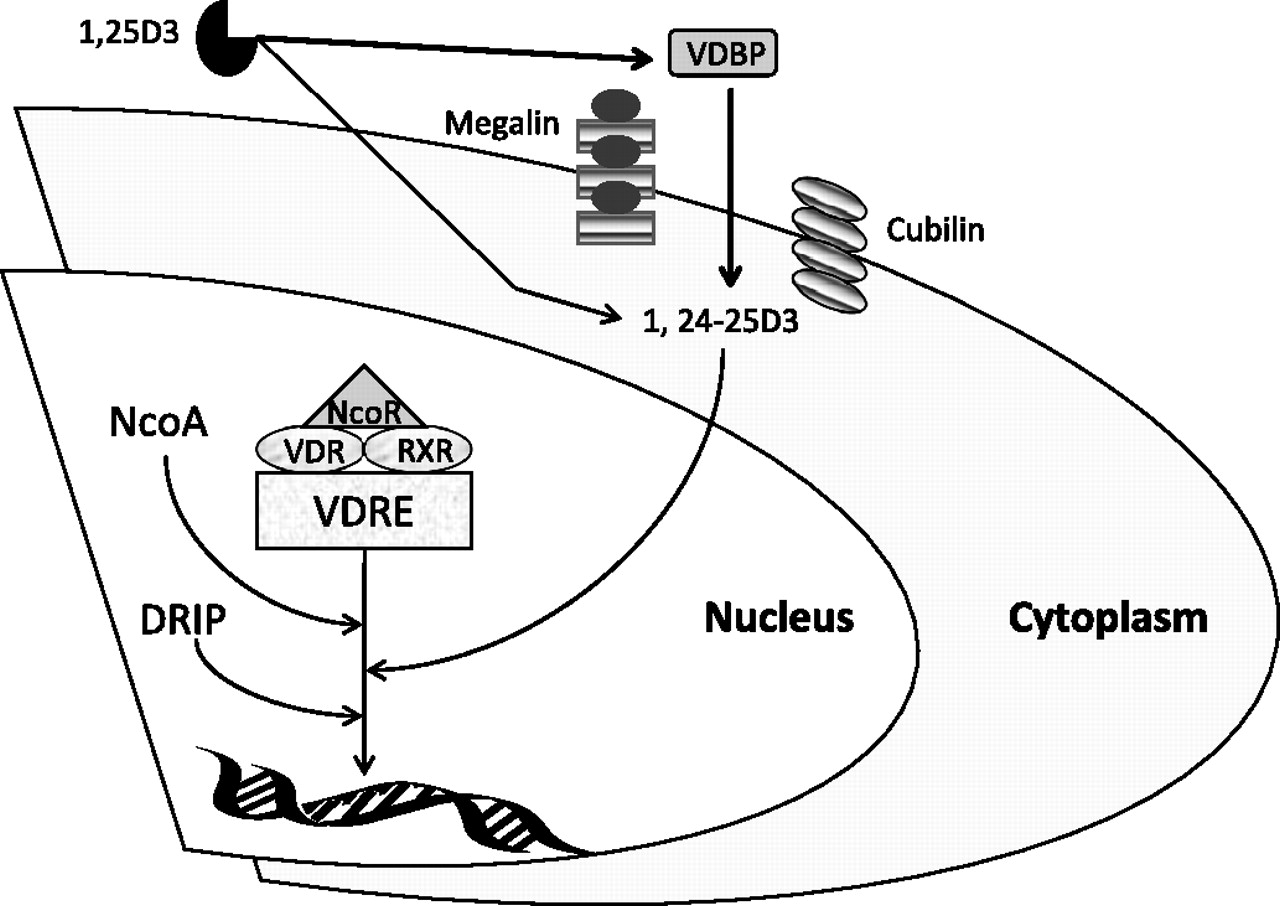
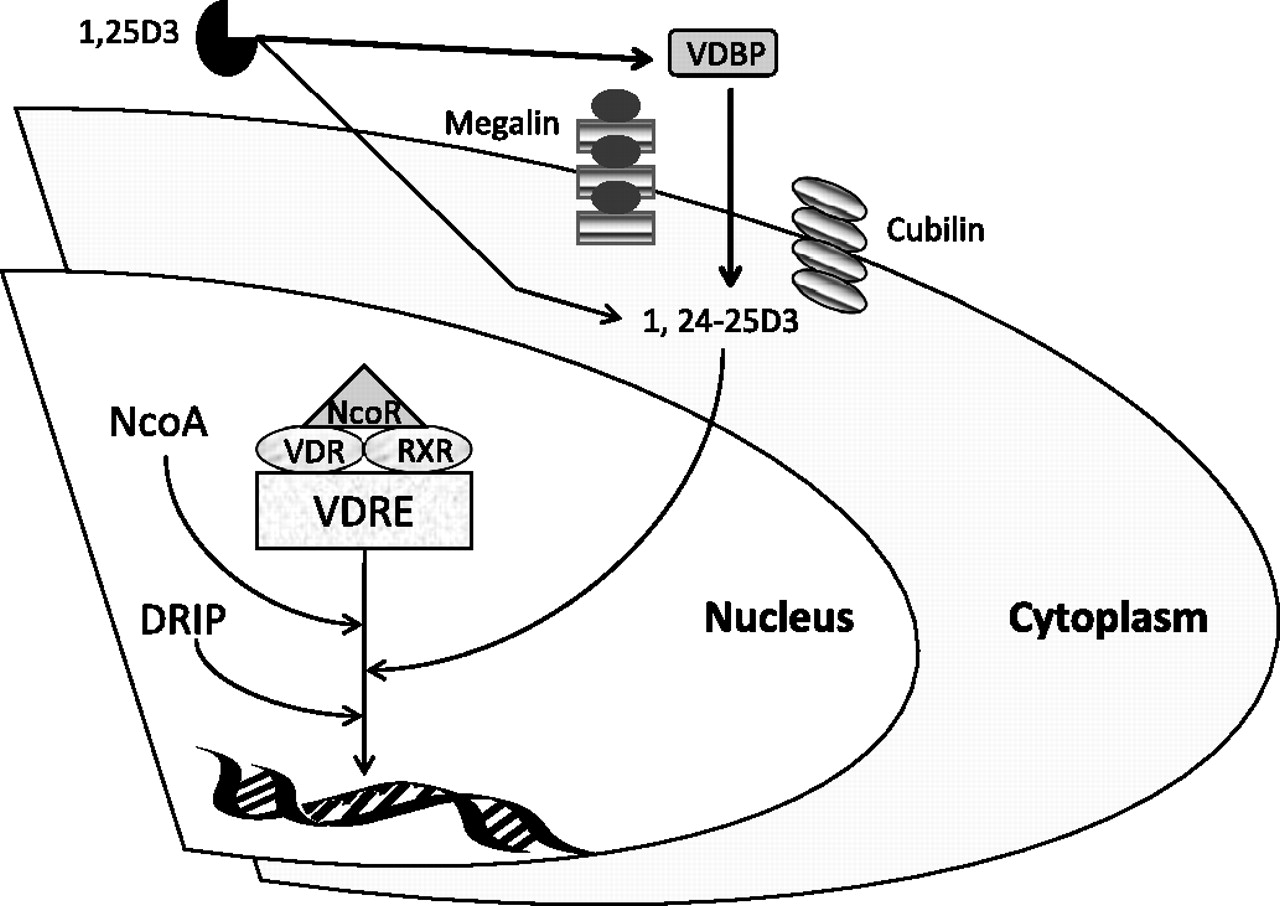{kind=link}
How the vitamin D axis influences cellular response. The diagram shows how vitamin D and vitamin D-binding protein (VDBP) influence gene transcription via the vitamin D receptor (VDR). Free 1,25-hydroxyvitamin D (1,25-OHD) enters cells via diffusion and bound 1,25-OHD by interaction of VDBP with transporter proteins (cubilin and megalin). The metabolite of 1,25-D3 (1,24-25-D3) binds to VDR. VDR is associated with corepressors (NCoR), which dissociate from VDR after ligands (such as 1,24-25-D3) bind. Ligand-bound VDR forms a heterodimer with the retinoid X receptor (RXR) and recruits coactivators (NCoA), resulting in histone acetylation. The resulting VDR complex initiates transcription of accessible DNA templates. The expression of VDR target genes results in cell growth inhibition and synthesis of antimicrobial peptides, such as catheledicin. DRIP, vitamin D receptor-interacting protein; VDRE, vitamin D receptor response element. Adapted from Ting et al.32
The ability of VDBP to bind actin blocks the formation of F-actin networks that might otherwise occlude the vasculature following cellular damage. The affinity for actin monomers is high (Kd=10−9 M),26 and is consistent with actin binding being a major function of VDBP. Since cell death may result in the liberation of large amounts of actin into the extravascular space, and VDBP acts to clear this from the circulation, we might expect VDBP levels to be depressed and actin–VDBP complexes elevated, in any condition where cell death occurs. This hypothesis has been confirmed in a single study of adult respiratory distress syndrome (ARDS), a disease characterised by massive cellular injury.35 Low circulating levels of VDBP have also been reported more recently in sepsis.36
The other main functions of VDBP relate to macrophage activation4 and neutrophil chemotaxis.11 VDBP is converted to macrophage-activating factor by the action of either β-galactosidase from B lymphocytes or sialidase from T lymphocytes on carbohydrate side chains of the protein.4 VDBP is not directly chemotactic to neutrophils, but does enhance the chemotactic effect of complement-derived peptides (C5a), which are able to interact with domain I of VDBP.37 CD44 and annexin 2 are thought to play a key role in subsequent interaction of VDBP complexes with neutrophils.38 VDBP-binding sites are upregulated on activated neutrophils,39 suggesting that changes in its circulating concentration might occur in inflammatory conditions. Consistent with this, in vitro work has shown that GC transcription is enhanced by proinflammatory cytokines.40 The functions of VDBP are summarised in table 3.
The functions of vitamin D-binding protein (VDBP)
Location of VDBP
VDBP is present in various body fluids including serum, peritoneal fluid41 and cerebrospinal fluid,42 and is also found on the surface of many cell types including human neutrophils.10 Although the vast majority of serum VDBP is derived from expression and secretion by liver parenchyma cells, small contributions by non-hepatic cell types including human monocytes are recognised.43 This implies widespread functionality of the protein, consistent with its effects on innate immunity and actin binding.
Chronic obstructive pulmonary disease
GC and genetic susceptibility
Cigarette smoking is the main environmental risk factor for developing COPD, but genetic susceptibility also plays a role in disease of both the airways and the parenchyma.44 In addition to the accepted susceptibility conferred by α-1-antitrypsin deficiency (AATD) (reviewed elsewhere45), adequately replicated genetic associations are recognised in candidate genes suggested by the pathogenetic themes of protease–antiprotease imbalance, oxidant–antioxidant imbalance and inflammation. In addition there may be a role for nicotine addiction, given the association between polymorphisms in the α-nicotinic acetylcholine receptor locus (CHRNA3/5) and COPD.46 The genes or regions associated with COPD in a least one meta-analysis, or in at least two independent populations in genome-wide association (GWA) studies are SERPINE2,47 the MMP cluster on chromosome 11,48 49 the CHRNA3/5 locus,46 HHIP,46 50 the GSTP genes,51 IL1B,51 EPHX152and TNFA.51 53 GC has been studied only in smaller COPD populations, and as yet is not the subject of a meta-analysis. Nevertheless it is appropriate to consider current evidence for its role in COPD.
Most studies of GC in COPD (summarised in table 4) have concentrated on known functional variants; such as the GC2 and GC1S alleles, caused by non-synonymous SNPs (rs4588 and rs7041). The relationship between these SNPs, the listed genetic variants and their resultant protein products is shown in table 2. As such it is now recognised that GC2, GC1S and GC1F are not alleles as such, but haplotypes composed of combinations of the SNPs at these loci. We will therefore refer to them as variants from here on. An individual may be homozygous or heterozygous for each variant, depending on the two haplotypes present. The GC1S variant has not been associated with COPD in any racial group.54–56 The GC2 variant appears protective in Caucasians.55–57 The GC1F variant has been consistently associated with a range of COPD phenotypes in Asian subjects,57–59 but results in Caucasians have been inconsistent.55 60 61 This is probably because Caucasians have a lower frequency of GC1F, thus the studies were underpowered. A single recent study has reported their results according to SNP genotype rather than the more commonly reported variants; in that work, the rs7041 TT genotype was associated with both low 25-OHD levels and risk of COPD.21
GC polymorphism and the risk of chronic obstructive pulmonary disease (COPD)
An alternative explanation for inconsistency of study results in COPD may be that the true functional variant lies in linkage disequilibrium (LD) with one of the studied variants, or that the phenotype it associates with is very specific, so it is not detectable in heterogeneous populations. With this in mind it is of interest that SNPs adjacent to GC associated with forced expiratory volume in 1 s (FEV1) in a recent GWA study62 implying that in an adequately powered study a genetic variant influencing FEV1 can be detected that may well exhibit a degree of LD with GC. Further information pertaining to the strength of LD between these SNPs and the GC2 and GC1F variants may help to ascertain if the reported association could in fact be due to GC variation.
In AATD, which is recognised to exhibit similar genetic associations to usual COPD, we have reported an influence of GC polymorphisms on airway bacterial colonisation and subsequent bronchiectasis.63 It is also of note that GC variants have been associated with diffuse panbronchiolitis58 (a condition seen almost exclusively in Japan) which leads to bronchiectasis.64 This may be due to similar mechanisms to those in COPD and AATD, but will require further study.
Very little functional work concerning GC polymorphisms has been undertaken in COPD; one study has reported differences in molecular structure which could play a role,65 but it is more likely that known differences in conversion to macrophage-activating factor4 are important. Differences in neutrophil chemotactic activity between the GC2, GC1F and GC1S proteins have not been found.61 Both activated macrophages and neutrophilic inflammation are believed to be important in COPD pathogenesis; since the former is affected by GC variation, this supports a role for the gene in susceptibility.
VDBP
There has been relatively little work concerning the role of VDBP in COPD, despite the wealth of genetic studies. The protein is present in bronchoalveolar lavage (BAL) fluid from subjects with COPD and asymptomatic smokers.5 66 In general VDBP levels were higher in COPD,5 but the effect of smoke exposure is uncertain, as results pertaining to this and VDBP function were inconsistent. It should be noted that since transcription of GC is affected by proinflammatory cytokines, VDBP might be considered an acute phase reactant. As such, differentiation of its role in the lung to ascertain if it is distinct from this will be important. VDBP also interacts with a key mediator of lung damage in COPD—neutrophil elastase (NE) cleaves the VDBP-binding site on neutrophils, such that VDBP is released into the circulation,67 where it would become bioactive. Furthermore, the ability of VDBP complexes to mediate neutrophil chemotaxis is prevented by NE inhibitors.67 This area warrants further research to clarify its relevance to COPD, and potential for modification by treatment.
Tuberculosis
GC and genetic susceptibility
Given that VDBP is involved in both neutrophil chemotaxis and macrophage activation, it is conceivable that airway defence against infection could be altered by GC genotype. Genetic susceptibility to TB has been found in a genome-wide linkage study,68 though to date most studies have been relatively small and concentrated on candidate genes relating to immune function. This limits their utility in determining new disease mechanisms but, where associations are well replicated, does not detract from their results. At least one GWA study is now underway,69 which has potential to reveal hitherto unrecognised aspects of pathogenesis. Meta-analysis level support for genetic susceptibility exists for SSC11A1 polymorphisms in Asians,70 and a non-synonymous SNP in interferon γ (IFNG).71
If the vitamin D axis plays a role in TB pathogenesis,3 variation within genes such as GC and VDR could be relevant in promoting resistance or susceptibility to the infection. VDR has been more widely studied and genetic variation within it appears to influence lymphocyte response to Mycobacterium tuberculosis.72 Nevertheless a meta-analysis of VDR studies was inconclusive,73 perhaps due to small study sizes and population heterogeneity. HIV status in particular may influence the apparent effect of susceptibility loci, perhaps because in HIV-positive individuals this surpasses the small risk attributable to genetic factors.74 Three studies have reported risk of TB infection in relation to GC genotype. GC2 homozygotes were more common in TB relative to controls in two studies in Asian populations,75 76 but the risk appears to depend on an interaction between vitamin D status and genotype.76 The association might be consistent with their reduced ability to convert GC to macrophage-activating factor, but requires further study of the reasons for synergy with vitamin levels. In Caucasian Russians, no GC genotype was associated with TB.77
VDBP and vitamin D
The main focus of vitamin D axis research in TB has been vitamin D itself. The purpose of this review is to concentrate on other aspects of the axis, so it is appropriate to consider how these relate to vitamin D concentrations. Circulating vitamin D levels have often been reported to be low in TB, as summarised elsewhere,3 and vitamin D deficiency has been proposed as part of a mechanism for annual influenza epidemics.78 Nevertheless the first trial of supplementation in TB did not improve outcomes79 perhaps because vitamin D concentrations were not improved markedly in the treatment group, or because part of the effect of the vitamin D axis in TB lies away from the vitamin. VDBP is elevated in serum during human80 and bovine81 mycobacterial infections, though it is not known if this relates directly to vitamin D. It is also unclear if VDBP is simply a marker of the acute inflammatory response, since other acute phase reactants were similarly elevated,80 or truly reflective of a role for the protein in pathogenesis. Further studies of both vitamin D and VDBP are warranted in infectious lung disease to clarify this.
Lung cancer
GC and genetic susceptibility
The vitamin D endocrine system is involved in a wide variety of biological processes including regulation of cell proliferation and differentiation, and can modulate such processes in cancer cells.82 Consequently GC and VDR may both be considered potential candidate genes for malignancy. The literature on genetic susceptibility to malignancy is extensive, and many GWA studies have been carried out already. In lung cancer the most well replicated associations at the GWA level lie in (or close to) CHRNA3/5,83 TERT84 and CLPTM1L.84 Meta-analysis level support exists for p53,85 XPC86 and hOGG1 (Asians only),87 amongst others. Several VDR polymorphisms have recently been associated at meta-analysis level with malignancy, but this was not specific to lung cancer.88 No studies have considered GC in lung cancer susceptibility, and studies of the GC1F and GC2 alleles in other malignancies have yielded conflicting results.89 90
VDBP and vitamin D
Animal and epidemiological studies suggest that active vitamin D metabolites (such as 1,25-OHD) prevent progression and metastasis of lung cancer.91 92 Consistent with this, higher rates of total cancer mortality have been reported in regions with less UV-B radiation, where vitamin D levels tend to be lower,92 and improved survival has been observed in patients with early-stage non-small cell lung cancer (NSCLC) with high vitamin D levels.93 The protective effects of vitamin D are believed to result from its role as a nuclear transcription factor that regulates cellular mechanisms central to the development of cancer, including cell growth, differentiation and apoptosis. Inhibition of metastasis and angiogenesis seems to be responsible for the anticancer effects of vitamin D in the lung.94 It should, however, be noted that although there may be good mechanistic reasons for suggesting that vitamin D has a direct role in pathogenesis, this has yet to be supported by clinical trial data in lung cancer indicating that supplementation and correction of deficiency is beneficial.
The VDR has also been studied in a murine lung cancer model and is present in both normal lung epithelial cells and those from lung adenomas.95 In one study, circulating VDBP levels were similar between subjects with cancer (although not specifically lung cancer) and healthy individuals.96 This does not necessarily mean that the vitamin D axis is not involved in cancer risk. Possible interpretations of the negative result are: (1) the vitamin D axis has a role local to the tumour site; (2) that the effect is mediated purely by vitamin D and VDR; or (3) VDBP's role is small and difficult to quantify. Current understanding of the role of the vitamin D axis in malignancy is shown in figure 1, and would tend to favour the first two of these hypotheses. However, the techniques for measurement of VDBP are now much more sensitive,97 such that further study of VDBP in lung cancer should be undertaken to determine any effect. This would be logical follow-up work to proteomic work in a mouse model of lung cancer, which suggested that VDBP acts as a disease biomarker.98
Asthma
GC and genetic susceptibility
In common with COPD, it is recognised that asthma may develop as a consequence of a variety of gene–environment interactions. Genetic associations of asthma have been widely published, and the findings and reproducibility reviewed recently.99 Briefly, those genes with associations replicated at the GWA level are IL4R, ORMDL3 and IRAK3,99 whilst additional meta-analysis level support exists for SNPs within IL4,100 TNFA,101 GSTM1,102 GSTT1,102 ADAM33103 and ADRB2.104 Once again, although genes in the vitamin D axis have been studied and implicated, they do not have the strongest supporting evidence in the field.
VDR has been more widely studied than GC in asthma, but results have been inconsistent.105–109 One small study of GC in asthma did not find an association, although it was probably underpowered to detect a difference.110 Overall these results suggest that genes within the vitamin D axis may be less important in asthma than COPD, perhaps because of the shift towards eosinophilic rather than neutrophilic inflammation.
VDBP and vitamin D
Murine models of pulmonary eosinophilic inflammation suggest that vitamin D supplementation alters cytokine expression profiles, immunoglobulin E levels, and the pattern of airway eosinophilia during allergen sensitisation, suggesting that the vitamin D axis could influence the development of allergy and asthma.111 Reports of both low and high 25-OHD levels in relation to asthma can be found in the medical literature, which have been reviewed elsewhere,112 though, as before, the relationship of this to the rest of the vitamin D axis has not been considered in detail. VDBP is also found in the BAL fluid in animal models of asthma,113 but there have been no reports comparing VDBP levels of healthy human subjects and those with asthma. The inconsistency of vitamin D studies and the paucity of evidence relating to VDBP mean that no firm conclusions regarding a role in asthma can be made currently.
Acute lung injury
Acute lung injury is pathologically characterised by diffuse alveolar damage, with neutrophils, macrophages and protein-rich oedema fluid in the alveolar spaces, together with capillary injury, and disruption of the alveolar epithelium.114 The involvement of neutrophils and macrophages might support a role for VDBP. However, little supportive evidence of the type seen in the other lung diseases discussed here exists. To date there have been few large studies of genetic susceptibility to acute lung injury, and no genome-wide work or meta-analyses of genetic susceptibility loci. The best replicated associations lie within the candidate genes IL6, ACE and SFTPB, the evidence for this being summarised elsewhere.115 No candidate gene studies pertaining to elements of the vitamin D axis have been published.
Since cell death may result in the liberation of large amounts of actin into the extravascular space, and VDBP acts to clear this from the circulation, we might expect VDBP levels to be depressed and actin–VDBP complexes elevated, in any condition where cell death occurs. This hypothesis has been confirmed in a single study of ARDS, a disease characterised by massive cellular injury.34 Low circulating levels of VDBP have also been reported more recently in sepsis.35
Conclusions
A growing body of research supports the view that vitamin D and VDBP influence the development of COPD and TB, and that they do so via immunomodulatory effects relating predominantly to macrophage function. Evidence of differences in protein function according to genotype, together with numerous genetic association studies, points to a role for VDBP in particular, with recent work suggesting that genotype interacts with vitamin D status to influence risk of disease. Functional work to clarify how VDBP contributes to pulmonary infection and inflammation, and how it relates to serum 25-OHD, is now under way and will be a key part of the investigation of the vitamin D axis, and supplementation of vitamin D in lung disease.
References
Footnotes
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.