Article Text

Abstract
Background: Chronic lung infection with Pseudomonas aeruginosa is the most severe complication for patients with cystic fibrosis (CF). This infection is characterised by endobronchial mucoid biofilms surrounded by numerous polymorphonuclear leucocytes (PMNs). The mucoid phenotype offers protection against the PMNs, which are in general assumed to mount an active respiratory burst leading to lung tissue deterioration. An ongoing respiratory burst by the PMNs has, however, not been demonstrated previously in endobronchial secretions from chronically infected patients with CF.
Objective: Based on the accumulating evidence for depletion of molecular oxygen (O2) in the mucus in infected CF bronchi, it was hypothesised that the O2 depletion in the mucus in infected CF bronchi may be accelerated by the respiratory burst of the PMNs due to the reduction of O2 to the superoxide anion (O-2) by the phagocyte NADPH oxidase (Phox).
Methods: Methods were established to isolate the O2 consumption by the respiratory burst from aerobic respiration in freshly expectorated sputum from chronically infected patients with CF.
Results: Inhibition of the Phox with diphenylene iodonium (DPI) delayed O2 depletion, nearly abolished staining of O-2-producing PMNs with hydroethidine and inhibited the rapid luminol-enhanced chemiluminescence in sputum. Furthermore, the total O2 consumption was correlated to the concentration of PMNs in the sputum samples.
Conclusion: The results demonstrate that CF sputum contains PMNs with an active consumption of O2 for O-2 production and suggest that the respiratory burst is ongoing and causes accelerated O2 depletion due to formation of O-2 in the lungs of chronically infected patients with CF.
Statistics from Altmetric.com
Chronic lung infection with Pseudomonas aeruginosa is the most severe complication in patients with cystic fibrosis (CF).1 The thick viscous endobronchial mucus hampers mucociliary clearance, which renders the CF lungs susceptible to chronic infections.2 The chronic P aeruginosa lung infection in patients with CF is characterised by endobronchial mucoid biofilms surrounded by numerous polymorphonuclear leucocytes (PMNs).3 4 In spite of the bactericidal mechanisms of the PMNs and aggressive antibiotic treatments, the biofilms persist. P aeruginosa may obtain protection against the PMNs by formation of mucoid biofilm.1 Thus, instead of clearing the bacteria the PMNs are suspected to promote progressive lung tissue deterioration by releasing proteolytic enzymes and reactive oxygen species (ROS) responsible for premature death in CF.1 5 Even though PMNs with evidence of priming and phagocytic activity have been isolated from CF airways, the activity of PMNs in the lungs of patients with CF with chronic P aeruginosa lung infection is far from being resolved.5 Interestingly, anaerobic conditions in the mucus have recently been demonstrated in non-obstructed bronchi of patients with CF with chronic P aeruginosa lung infection.3 The cause of this O2 depletion may be O2 consumption by the epithelia and by P aeruginosa.3 We hypothesised that the respiratory burst of the PMNs contributes to the O2 consumption in the mucus of the infected CF airways. The interaction between bacteria and the PMNs may increase O2 consumption as a consequence of the extra respiration during phagocytosis.6 This process is caused by the respiratory burst resulting from the one-electron reduction of O2 by the phagocyte NADPH oxidase (Phox) to O-2.7 To evaluate the capability of the respiratory burst to deplete O2, we constructed a simple reaction chamber that allowed real-time measurement of the O2 concentration during phagocytosis of P aeruginosa by PMNs. The respiratory burst was isolated from aerobic respiration by blocking the respiratory chain with KCN, which previously has been employed to demonstrate the resistance of the respiratory burst to inhibition of the aerobic respiration in PMNs.8 The aerobic respiratory component was exposed by inhibiting the Phox with diphenylene iodonium (DPI). These in vitro findings were utilised to estimate the O2 consumption and the ROS production by the respiratory burst of the PMNs in freshly expectorated sputum from patients with CF with chronic P aeruginosa lung infection.
Methods
Sputum samples
The study was performed on leftover sputum expectorated for routine bacteriology from 29 patients with CF with chronic P aeruginosa infections after obtaining permission by informed consent.
Characteristics of the 29 patients with cystic fibrosis
Chronic P aeruginosa infection was defined as the presence of P aeruginosa in the lower respiratory tract at each monthly culture for ⩾6 months, or for a shorter time in the presence of increased antibody response to P aeruginosa (⩾2 precipitating antibodies, normal: 0–1).9 Before measurement of luminol-enhanced chemiluminescence (LEC) and dissolved O2, the samples were brought into homogenous suspensions by gentle resuspension with a pipette. Normoxic conditions were achieved by diluting 10 times with Krebs–Ringer buffer equilibrated in ambient air. The concentration of leucocytes in the diluted sputum was determined as previously described.10 11 The estimation of the frequency of PMNs is described in online supplementary fig 5.
Preparation of PMNs
Human blood samples were obtained from normal healthy volunteers by venous puncture and the PMNs were isolated by gradient sedimentation and centrifugation, as previously described, before suspending the PMNs in Krebs–Ringer buffer with 10 mM glucose and 5% (v/v) normal human AB-positive sera at a density of 5×109 PMNs/l.10 11
Bacteria
Inoculum for phagocytosis by PMNs was prepared from exponentially growing cultures of the widely studied P aeruginosa wild-type strain (PAO1) (online supplementary fig 1).
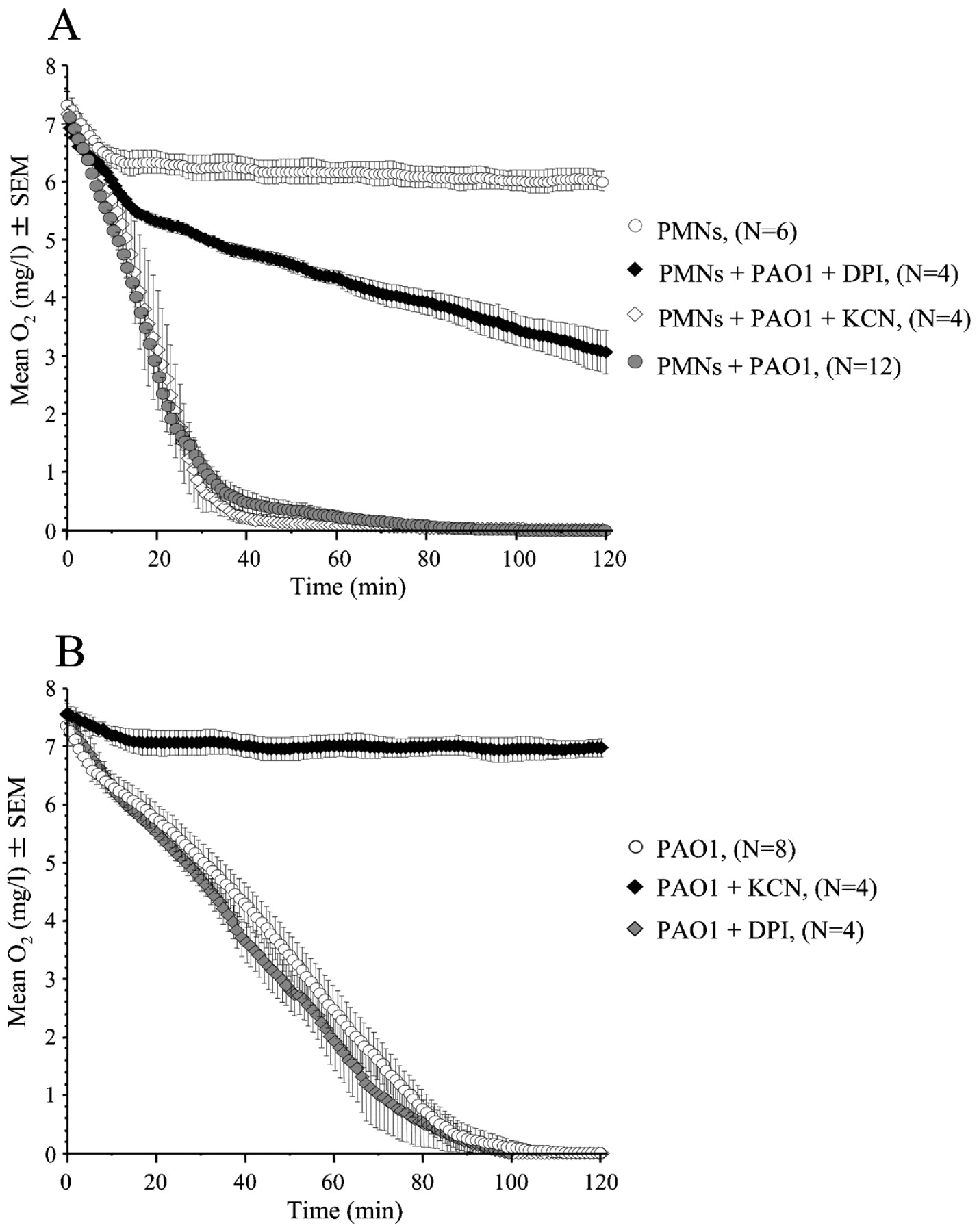
O2 consumption by the respiratory burst and aerobic respiration in the reaction chamber. (A) Increased O2 consumption during phagocytosis (p<0.001), inhibition of the respiratory burst during phagocytosis by treatment with 50 μM diphenylene iodonium (DPI) (p<0.001), and unchanged O2 consumption during phagocytosis by treatment with 2 mM KCN during phagocytosis of strain PAO1 (15×109/l) by normal polymorphonuclear leucocytes (PMNs) (5×109/l). (B) Decreased O2 consumption by PAO1 (15×109/l) during treatment with 2 mM KCN (p<0.001) and unaffected bacterial O2 consumption during treatment with 50 μM DPI. Statistical analysis was performed by two-way analysis of variance.
Estimation of the respiratory burst by LEC
The LEC from aliqouts of 100 μl of isolated PMNs or sputum was determind as outlined in online supplementary fig 3.
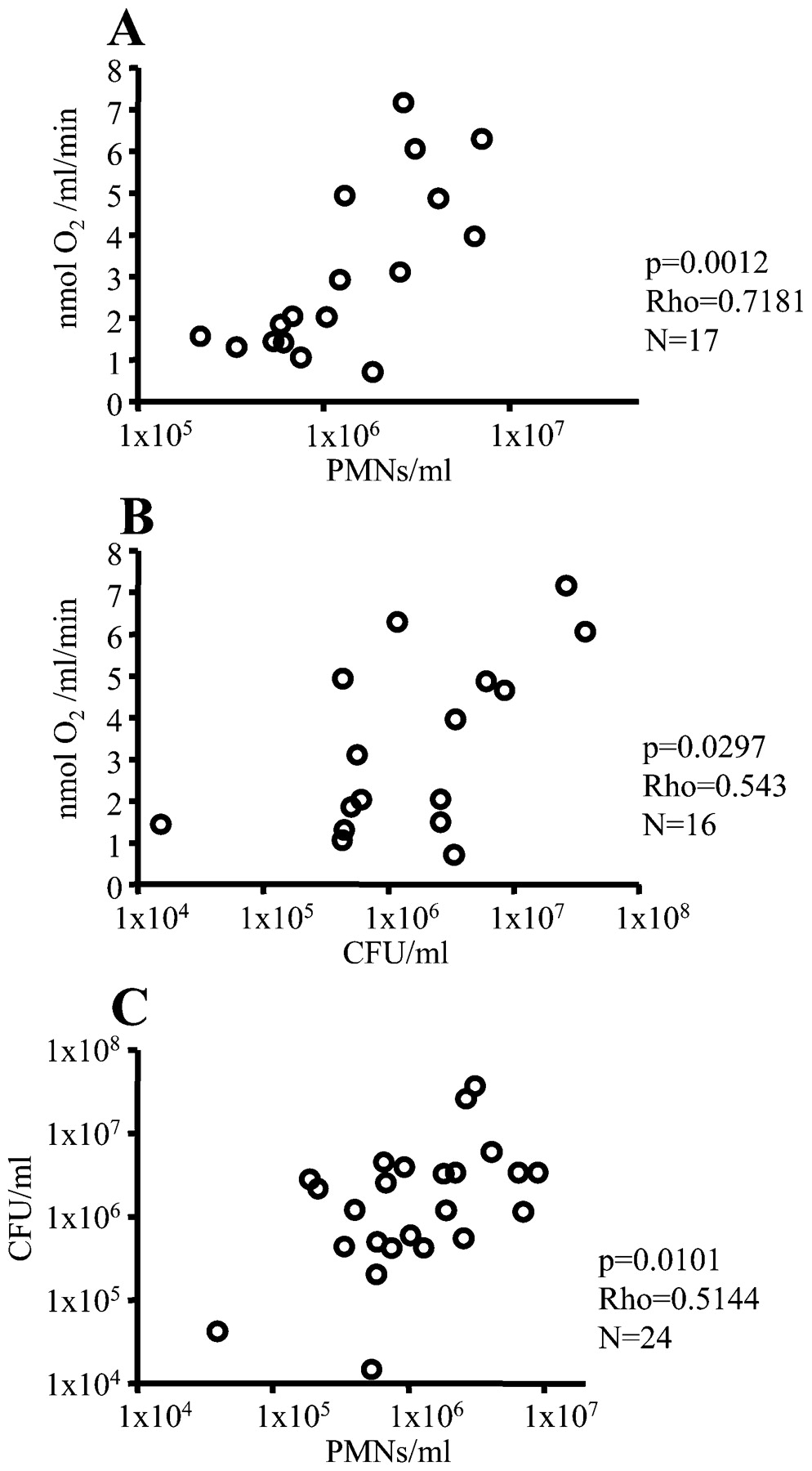
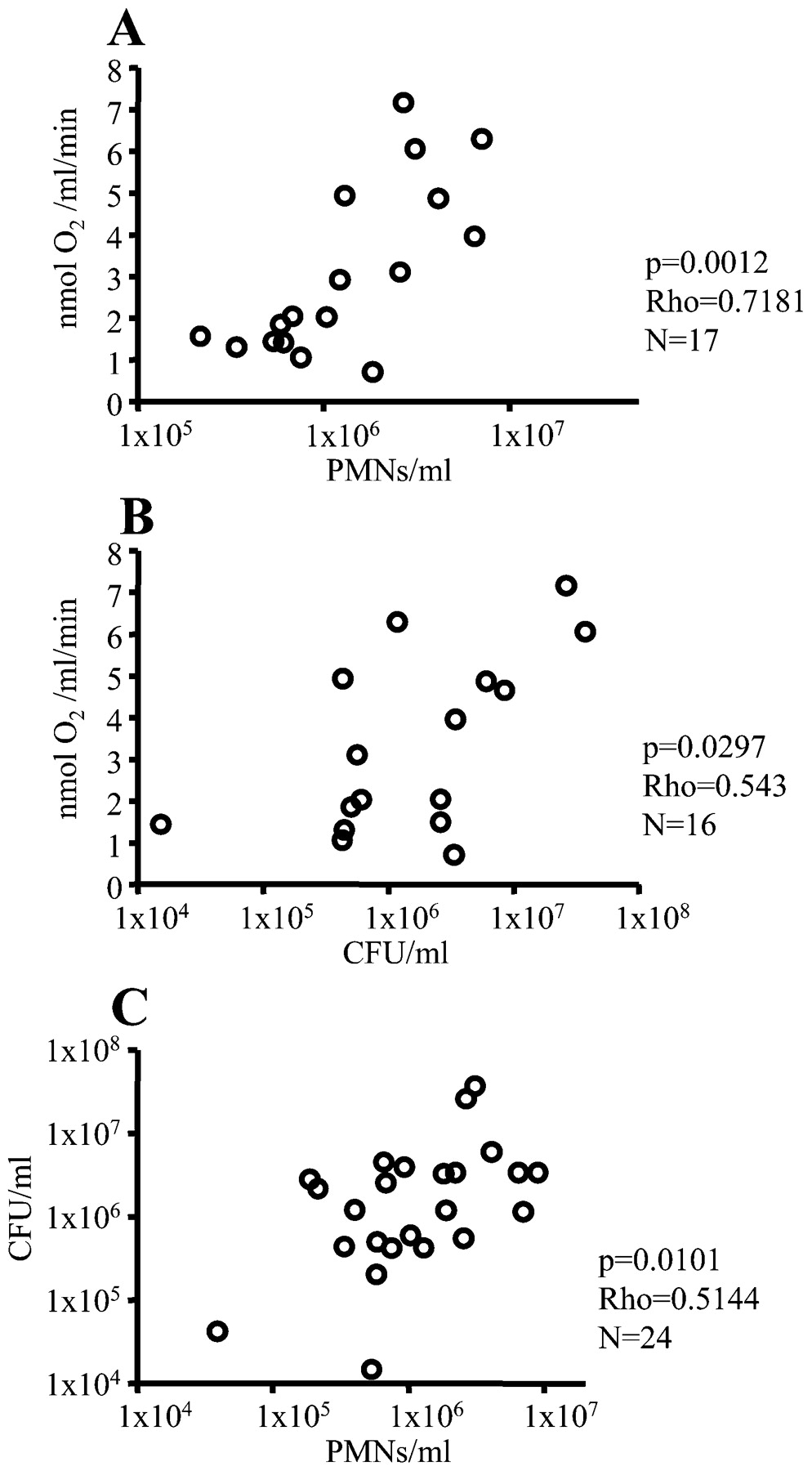
Correlations between the O2 consumption rate, the concentration of polymorphonuclear leucocytes (PMNs) and colony-forming units (CFUs) in diluted sputum from patients with cystic fibrosis with chronic P aeruginosa lung infection. The O2 consumption rate was determined in the reaction chamber, the concentration of PMNs was estimated by flow cytometry, and the CFUs were counted by plating of serial dilutions. (A) O2 consumption rate vs the concentration of PMNs. (B) O2 consumption rate vs the concentration of CFUs. (C) The concentration of PMNs vs the concentration of CFUs. Statistical analysis was performed by Spearman rank correlation test.
The concentration of dissolved O2
Measurement of dissolved O2 in sputum and during phagocytosis is described in online supplementary fig 2.
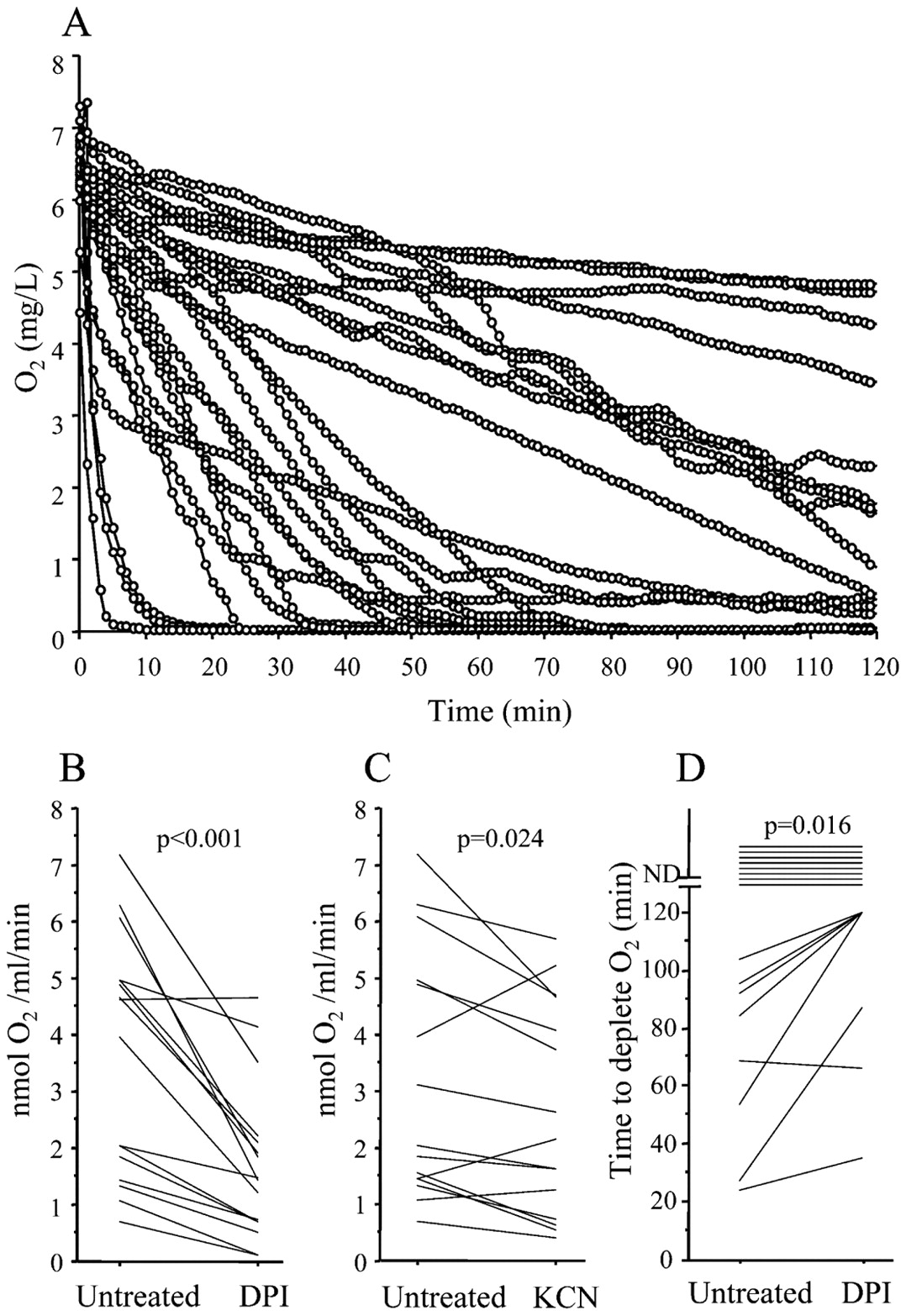
O2 consumption and reactive oxygen species generation in diluted sputum from patients with cystic fibrosis with chronic P aeruginosa lung infection. (A) The decreasing O2 concentration as a function of incubation time measured in the reaction chamber (n = 23). (B) Reduced O2 consumption rate in sputum treated with 50 μM diphenylene iodonium (DPI) (n = 16). (C) Reduced O2 consumption rate in sputum treated with 2 mM KCN (n = 15). (D) Postponing of O2 depletion in sputum treated with 50 μM DPI. In B–D, data were analysed by Wilcoxon signed rank test.
Visualiaation of O-2 production
Hydroethidine (HE) (D7008, Sigma, St Louis, Missouri, USA) at 1 μM was added to 100 μl of sputum and the samples were incubated for 15 min at 37°C before examination with a fluorescence microscope (BX40, Olympus, Ballerup, Denmark). To verify the origin of O-2, sputum was pretreated for 10 min with 50 μM DPI before addition of HE. The interaction between PMNs and bacteria in the sputum samples was visualised by adding HE (1 μM) together with the green permeable DNA dye SYTO 9 (Molecular Probes, Eugene, Oregon, USA) (10 μM), and the samples were incubated for 15 min at 37°C before examination.
Statistical methods
Statistical significance was evaluated by two-way analysis of variance (ANOVA) with Bonferroni post-tests for observations with parametric data. Non-parametric data were analysed by Wilcoxon signed rank test and Spearman rank correlation test. A p value <0.05 was considered statistically significant. The tests were performed with Prism 4.0c (GraphPad Software, La Jolla, California, USA).
Results
O2 consumption by PMNs and P aeruginosa during phagocytosis
Estimations of the O2 consumption during phagocytosis were initiated at atmospheric O2 concentration. Accelerated initial O2 consumption with a reduced O2 consumption rate at low O2 concentrations was seen when mixing PMNs with PAO1 (fig 1A). Nevertheless, the O2 concentration decreased below the level of detection (0.01 mg O2/l). This was considered as an indication of anaerobic conditions, since similar recordings were obtained in buffer equilibrated in an anaerobic bench with growing strict anaerobes as control. Low O2 consumption by unstimulated PMNs was also seen (fig 1A). Evidence of phagocytosis was provided by the reduced number of surviving P aeruginosa in the presence of PMNs (online supplementary fig 1) and by the green fluorescent protein (GFP)-tagged P aeruginosa being ingested by or attached to the PMNs in the reaction chamber (online supplementary fig 2).
The O2 consumption of P aeruginosa resulted in anaerobic conditions within 2 h (fig 1B) and indicated that P aeruginosa may consume a considerable part of the O2 during phagocytosis.
Inhibition of the respiratory burst with DPI during phagocytosis of P aeruginosa by PMNs
Treatment with 50 μM DPI reduced the chemiluminescence, which predominantly reflects O-2 production during phagocytosis, to the level of unstimulated PMNs for up to 50 min (online supplementary fig 3A). Thus, inhibition of the respiratory burst with 50 μM DPI during phagocytosis resulted in reduced O2 consumption and prevention of anaerobic conditions (fig 1A). Since treatment with 50 μM DPI did not affect bacterial O2 consumption (fig 1B), growth, and survival after phagocytosis (online supplementary fig 1) significantly, 50 μM DPI was selected for estimating the O2 consumption by the respiratory burst of the PMNs.
Inhibition of the respiratory chain with KCN during phagocytosis of P aeruginosa by PMNs
Unexpectedly, the LEC was increased (online supplementary fig 3B) without affecting the O2 concentration (fig 1A) during phagocytosis inhibited with KCN. This could not be ascribed to P aeruginosa since the bacteria ceased to consume O2 shortly after addition of 2 mM KCN (fig 1B). Instead, the surprisingly high O2 consumption and LEC during KCN treatment may result from a higher amount of O2 being available for the respiratory burst due to the bacteria failing to consume O2. In addition, KCN repression of aerobic respiration may decrease bacterial motility and facilitate capture of P aeruginosa by the PMNs, resulting in a stronger respiratory burst. Though KCN treatment may overestimate the O2 consumption by the respiratory burst, it was decided to use 2 mM KCN for the study of sputum. Lower concentrations may be insufficient to block the pseudomonal cyanide-independent terminal oxidase.12
O2 consumption by the respiratory burst in sputum from patients with CF with chronic P aeruginosa lung infection
To ensure similar initial O2 concentrations, the expectorated sputum was diluted 10 times in Krebs–Ringer buffer equilibrated in ambient air. O2 consumption was observed in all sputum samples by the decreasing O2 concentration, and anaerobic conditions were established in 10 of the 23 sputum samples (fig 2A). The chemiluminescence was inhibited >95% after 10 min of treatment with DPI whereas KCN treatment had no effect (online supplementary fig 4). Thus, the sputum was pretreated with DPI and KCN for 10 min to estimate the O2 consumption rate during the following 30 min before the O2 was depleted. The O2 consumption rate was reduced by DPI treatment, indicating an active respiratory burst in the sputum samples, and resulted in a mean inhibition of the O2 consumption rate by 56% (SD 20%) (fig 2B), suggesting that the respiratory burst consumed the major part of the O2. Active O2 consumption by the respiratory chain was also observed by the reduced O2 consumption rate caused by addition of KCN (fig 2C). The consequence of inhibiting the respiratory burst with DPI was a delayed generation of anaerobic conditions (fig 2D) whereas inhibition of the respiratory chain with KCN was insufficient to delay the depletion of O2 significantly. DPI treatment allowed us to detect comparable levels of the O2 consumption rate by the burst per PMN in CF sputum and in the phagocytosis experiment using normal PMNs and PAO1. In the sputum we found 1.7 nmol O2/106 PMNs/min (SD 1.6 nmol O2/106 PMNs/min) and in the experimental phagocytosis assay we found 1.5 nmol O2/106 PMNs/min (SD 0.4 nmol O2/106 PMNs/min). These data make it unlikely that the O2 consumption rate of sputum PMNs is a CF-specific phenomenon.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
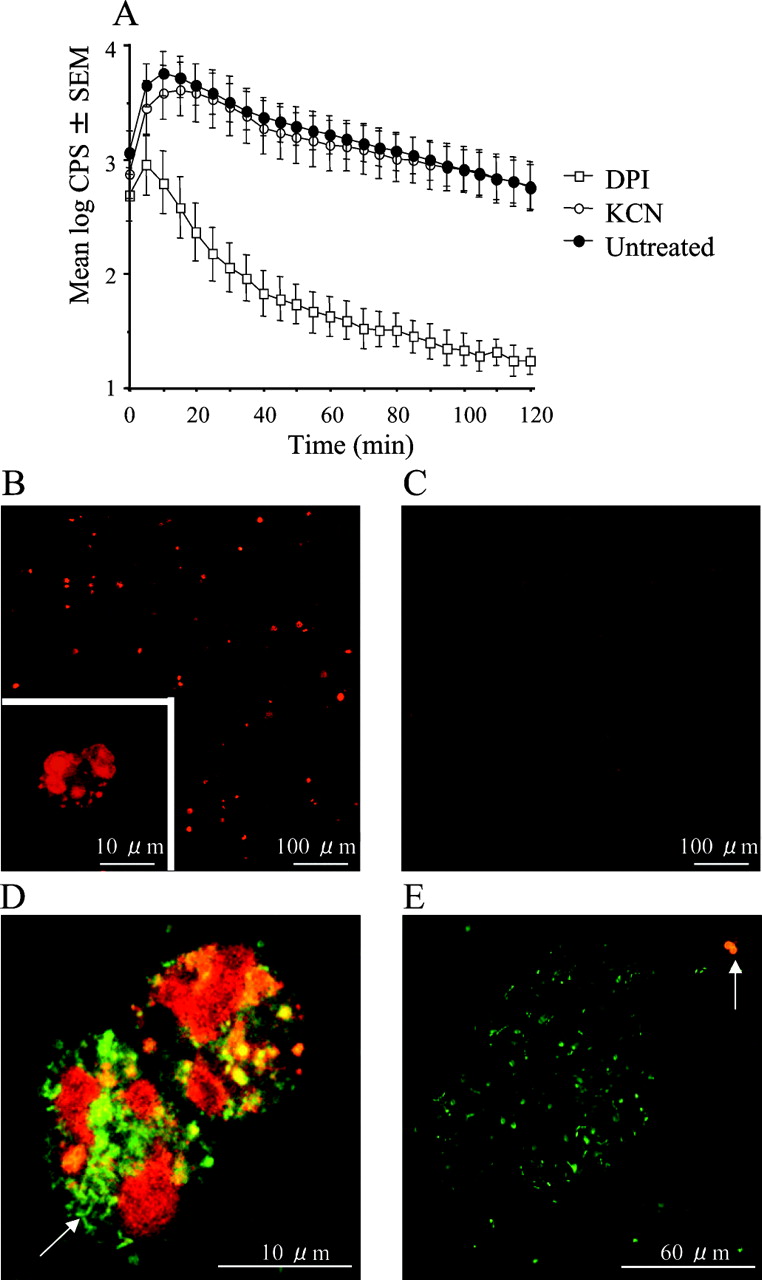
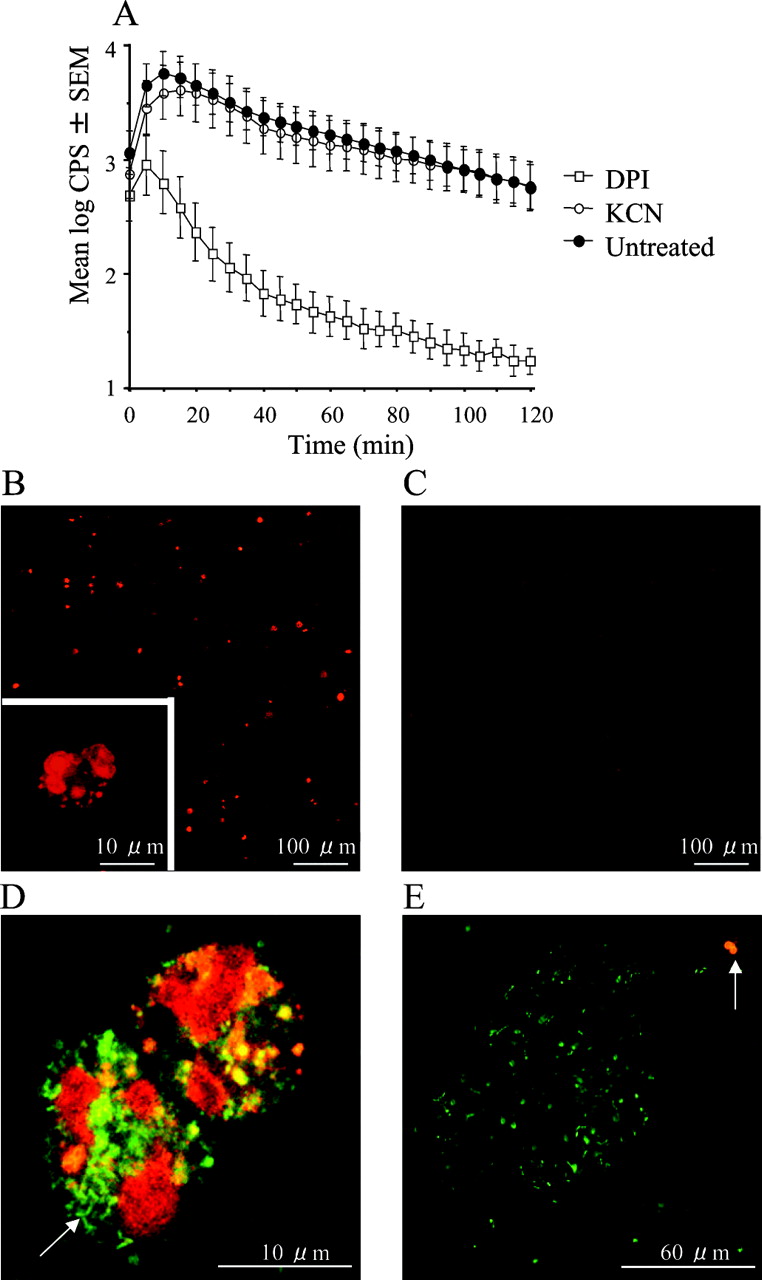
Reactive oxygen species (ROS) generation in sputum from patients with cystic fibrosis with chronic P aeruginosa lung infection. (A) The effect of treatment with KCN and inhibition with diphenylene iodonium (DPI) (p<0.001) on ROS generation shown as counts per second (CPS) from luminol-enhanced chemiluminescence (n = 10). Statistical analysis was performed by two-way analysis of variance (n = 10). (B–E) Visualisation of O-2 production by fluorescence microscopy of sputum stained with hydroethidine. (B) Untreated sputum. (C) Sputum treated with 50 μM DPI to inhibit the phagocyte NADPH oxidase. (D) Polymorphonuclear leucocytes (PMNs) with ingested planktonic bacteria (arrow) with green fluorescence from SYTO 9. (E) A bursting PMN (arrow) engaged with a biofilm with bacteria emitting green fluorescence from SYTO 9.
PMNs with respiratory burst in sputum from patients with CF with chronic P aeruginosa lung infection
The mean fraction of PMNs constituted 90% (7%) (SD) of the host cells in the sputum samples. Therefore, the concentration of PMNs was compared with the O2 consumption rate in the diluted sputum samples. A positive correlation was observed, supporting the involvement of PMNs in the O2 consumption (fig 3A). Positive correlations were also observed between the concentration of colony-forming units (CFUs) and the O2 consumption rate (fig 3B) and between the concentration of PMNs and the concentration of CFUs (fig 3C). No significant correlation was observed between the O2 consumption rate and forced expiratory volume in 1 s (FEV1) or forced vital capacity (FVC).
To avoid unintended effects of the Krebs–Ringer buffer, the respiratory burst in undiluted sputum samples was estimated by LEC. As in diluted sputum, DPI treatment resulted in >95% inhibition after 10 min in undiluted sputum (fig 4A), indicating an active respiratory burst that persisted during the 2 h of observation. Furthermore, the LEC was resistant to KCN, suggesting an origin that may function independently of the respiratory chain such as the Phox of the PMNs.8 To verify further the PMNs as the source of ROS in sputum, O-2 production was visualised by staining with HE, which is oxidised by O-2 to generate 2-hydroxyethidium that emits intensified red fluorescence when complexing with DNA.13 In the untreated sputum, several fluorescent objects were seen at low magnification. Higher magnification revealed these objects to be PMNs (fig 4B). In DPI-treated sputum, only faint fluorescence was seen (fig 4C). By applying the validated HE to fresh, expectorated sputum together with the permeable DNA stain, SYTO 9, we found bursting PMNs with ingested planktonic bacteria (fig 4D) and bursting PMNs failing to engulf bacteria from a biofilm (fig 4E).
Discussion
PMNs in bronchoalveolar lavage fluids from chronically infected patients with CF and in sputum from healthy individuals have the potential to mount a respiratory burst upon stimulation.14 15 In contrast, the presence of PMNs with an active respiratory burst was demonstrated in fresh sputum from patients with CF with chronic P aeruginosa lung infection in this study. Both the inhibition of O2 consumption and ROS generation with DPI as well as the correlation of the PMN concentration to the O2 consumption rate are indications of an ongoing respiratory burst by the PMNs in sputum from chronically infected patients with CF. PMNs with an active respiratory burst were not demonstrated directly in the lungs of the patients in this study. Therefore, our findings do not necessarily reflect the situation of all PMNs in the endobronchial mucus of infected CF lungs since the pO2 may differ at various sites in the sputum plugs. However, the rapid, high chemiluminescence, observed in undiluted sputum ex vivo, may exclude the possibility that adding Krebs–Ringer buffer, equilibrated to ambient air, causes the oxidative burst. In this context, the initial, high O2 consumption and ROS generation seen in untreated sputum samples within 30 min after expectoration suggest that the respiratory burst of the PMNs was already active in the mucus in the lungs. Thus, our results indicate that the respiratory burst of the PMNs is the main contributor to the intensive O2 depletion inside the mucus of the bronchi.3 The respiratory burst of the PMNs in the sputum may be activated by direct contact with P aeruginosa5 16 and by the binary signalling from the engagement of the integrins and binding to inflammatory cytokines.17 Interestingly, we have recently found levels of tumour necrosis factor α (TNFα) with the potential to augment the respiratory burst in sputum from several infected patients with CF attending the Copenhagen CF Centre, as in the patients in the present study.18 In addition, comparable levels of TNFα in sputum from infected patients with CF are reported from other CF centres.19 Besides TNFα, platelet-activating factor, leukotriene B4, interleukin-8, lipopolysaccharide, antibodies and alginate have been suggested to be major triggers for the induction of the respiratory burst of airway PMNs.1 5 18 19 In contrast, the respiratory burst of the PMNs may be repressed by quorum sensing-regulated secretion of rhamnolipid, by exoprotease from P aeruginosa, by antibiotic treatment and by the high levels of apoptotic or necrotic cells in the sputum samples.10 11 20 21 22 These opposing mechanisms may contribute to the variation observed in the correlation between the concentration of PMNs and the O2 consumption in the sputum. Our results imply that at least in some parts of the CF mucus O2 is present, which allows PMNs to create a respiratory burst. This is in accordance with the steep pO2 gradients in CF mucus.3 Furthermore, based on the substantial fraction of PMNs able to create a significant respiratory burst found in CF sputum in this study, our previous demonstrations of the toxic effects of P aeruginosa rhamnolipid on PMNs may not apply to the entire population of PMNs in the sputum.10 11
The demonstration of PMNs with an active respiratory burst in sputum from chronically infected patients with CF may have been anticipated due to the ROS lesions in the infected CF lungs.5 23 However, it could not have been taken for granted. In fact, the ROS in the CF lungs may be derived from sources other than the Phox of the PMNs, including epithelial cells, alveolar macrophages, and from mitochondrial respiration.23 24 25 Due to the predominance of PMNs in the sputum samples the contribution of other cells to O2 metabolism was regarded to have low impact on the results in this study. In contrast to the Phox, mitochondrial activity is unlikely to influence significantly our observations in the sputum samples. PMNs contain only few and poorly developed mitochondria with a low activity of the respiratory chain.26 In addition, sputum from chronically infected patients with CF may contain P aeruginosa producing pyocyanine, which may also cause O-2 formation.27 Thus, O-2 formation by pyocyanine may be a reason why DPI failed to block ROS generation completely in the sputum samples.
Previously, faster O2 depletion has been achieved when PMNs were added to P aeruginosa growing in experimental biofilms due to increased O2 consumption of both the bacteria and the PMNs.16 However, in the present study the ability of the PMNs per se to create anaerobic conditions by an active respiratory burst was observed in the KCN-resistant O2 depletion during the phagocytosis of PAO1 and in the sputum samples. This may, however, be expected, since PMNs are able to produce a substantial part of the energy needed for the respiratory burst by anaerobic glycolysis resulting in l-lactate formation.28 Accordingly, our suggestion of an active respiratory burst by the PMNs in the CF lungs is supported by the increased glucose uptake in PMNs in CF lungs and the high concentration of l-lactate in sputum from patients with CF with chronic P aeruginosa lung infection.29 30
The demonstration of PMNs with an active respiratory burst in sputum samples from patients with CF with chronic P aeruginosa lung infection verifies the frequent assumption that the PMNs induce ROS lesions in the lungs. Sufficient O2 to produce O-2 by the respiratory burst of the PMNs in sputum may be supplied by the daily inhalation of 8000 litres of O2-rich air.23 The result of the fast dismutation of O-2 is H2O2, which may lead to the formation of myeloperoxidase-derived oxidising species found in CF airways.5 Considering the short half-lives of the ROS generated by PMNs, the tissue adjacant to the mucus is most likely to be exposed to PMN-derived ROS. Therefore, ROS generated in the mucus may be a reason for the affected bronchi in patients with CF with chronic P aeruginosa lung infection.31 However, it is less obvious how ROS generated in the mucus may reach more distant lung tissue. Since the included patients with CF show a broad range of FEV1 values, one may suggest that the O2 consumption rate might be linked to pulmonary function in these patients. However, no significant correlation was observed between the O2 consumption rate and FEV1 or FVC. This may be due to the condition of the infected patients, which has been stabilised for years by elective antibiotic treatment, resulting in a slower decline of lung function and fewer excerbations. Thus, the lung function of the included patients may primarily result from slowly accumulated deterioations of the lung tissue rather than the ongoing inflammation in the endobronchial mucus. Our results provide arguments to control the extent of ROS by antibiotic treatment, since O2 consumption was mainly caused by ROS formation of the PMNs and was correlated to the concentration of P aeruginosa in the sputum samples. The latter correlation probably results from a balanced accumulation of PMNs in response to the concentration of P aeruginosa in the sputum samples.
As indicated in fresh sputum samples, the ROS from the respiratory burst of the PMNs in the mucus may reach the bacteria, due to their ingestion by PMNs, with a respiratory burst and the intimate surrounding of the endobronchial biofilms by the PMNs.4 Though the killing of P aeruginosa by PMNs was apparently independent of ROS formation in this study, the surviving bacteria may experience ROS lesions, which may affect virulence. In particular, PMN-derived ROS may cause mutations in the mucA gene, leading to formation of the crucial mucoid phenotypes.32 In fact, mutations in the mucA gene were found in the majority of the mucoid isolates from 91 Scandinavian patients with chronic P aeruginosa infection.33 In addition, the PMN-derived ROS may induce highly mutagenic lesions in the bacterial DNA that are associated with resistance to antibiotics and accumulation of hypermutable isolates.34
The accelerated O2 depletion by the PMNs probably affects the O2 distribution in the endobronchial mucus. Though the precise O2 profile of the endobronchial mucus has not been determined, the activity of the PMNs is sensitive to the local O2 tension. In particular, the accumulation of PMNs in the lung secretions may be increased due to the postponing of apoptosis by hypoxia.35
Sputum from chronically infected patients with CF may also contain biomarkers for the bacterial response to O2 depletion such as slow growth and cyanide secretion.12 36
This study demonstrates an ongoing respiratory burst of the PMNs in sputum from patients with CF with chronic P aeruginosa lung infection that may induce environmental changes in the lungs with the potential to generate virulent and resistant bacterial phenotypes, to facilitate colonisation by anaerobic bacteria and to cause oxidative lesions in the lung tissue of the patients.
Acknowledgments
The skilful assistance of Marjan Yousefi, Louise Falk-Lauritsen and Lars Kolpen is gratefully appreciated and recognised as being crucial for this study.
REFERENCES
Footnotes
▸ Additional figures are published online only at http://thorax.bmj.com/content/vol65/issue1
Funding None.
Competing interests None.
Provenance and Peer review Not commissioned; externally peer reviewed.