Article Text

Abstract
Background: Human β-defensin (HBD)-1 and -2 are antimicrobial peptides present in the respiratory tract. Recent reports have indicated reduced activity of β-defensins in cystic fibrosis, suggesting that β-defensins may play an important role in the pathological process of chronic respiratory tract infection. Diffuse panbronchiolitis (DPB) is a progressive disease characterised by frequent episodes of superimposed infection, typically caused by Pseudomonas aeruginosa. The aim of this study was to elucidate the role of these antimicrobial peptides in this disease.
Methods: The concentrations of HBD-1 and HBD-2 in plasma and bronchoalveolar lavage (BAL) fluid from 33 patients with DPB and 30 normal adults were measured by radioimmunoassay. Localisation of HBD-2 was investigated immunohistochemically in an open lung biopsy specimen obtained from a patient with DPB.
Results: High concentrations of HBD-1 and HBD-2 were noted in BAL fluid from DPB patients. Increased plasma concentrations of HBD-2, but not HBD-1, were found in patients with DPB compared with control subjects. In patients with DPB the HBD-2 concentration in BAL fluid correlated significantly with the numbers of cells recovered from the BAL fluid (total cells, neutrophils, and lymphocytes) and with the BAL fluid concentration of IL-1β. Synthetic HBD-2, but not HBD-1, had dose dependent bactericidal activity against P aeruginosa. Treatment of 14 patients with macrolides significantly reduced BAL fluid concentrations of HBD-2 but not HBD-1 or plasma concentrations of HBD-1 and HBD-2. Immunohistochemistry of lung tissue showed localisation of HBD-2 in the epithelia of the distal bronchioles.
Conclusions: These results indicate that β-defensins, particularly HBD-2, participate in antimicrobial defence in the respiratory tract in DPB, and that the BAL fluid concentration of HBD-2 may be a useful marker of airway inflammation in patients with DPB.
- human β-defensin-1,2
- diffuse panbronchiolitis
- bronchoalveolar lavage fluid
Statistics from Altmetric.com
Antimicrobial peptides have been identified as key elements in the innate host defence against infection. Defensins are single chain strongly cationic peptides of molecular weight 3000–4500 and are one of the most extensively studied classes of antimicrobial peptides.1,2 They have broad spectrum microbicidal activities against various Gram positive and Gram negative bacteria, mycobacteria, fungi, and certain enveloped viruses. Human defensins are classified as α- and β-defensins based on the arrangements of three intramolecular disulfide bonds. At present the human α-defensin family comprises six members, four of which (human neutrophil peptides (HNP)-1 to -4) are located in the azurophilic granules of the neutrophil and two of which (human defensin-5 and -6) are present in the secretory granules of the intestinal Paneth’s cells. Four β-defensins have also been isolated in humans. Human β-defensin (HBD)-1 was originally isolated from blood filtrate and expressed mainly in the urogenital tract.3–5 HBD-2 and HBD-3 were isolated from psoriatic scale extracts from patients with psoriasis using a whole Escherichia coli affinity column or Staphylococcus aureus affinity column that binds cationic antimicrobial peptides.6,7 HBD-4 was found by analysis of genomic sequences mapping at chromosome 8p23 where all the known α- and β-defensins are clustered.8 The HBD-4 gene is expressed mainly in the testis and, to a lesser degree, in the gastric antrum. Both HBD-1 and HBD-2 genes are also expressed in human trachea and lung and are believed to contribute to antimicrobial defence in the respiratory tract.9–11
Diffuse panbronchiolitis (DPB) is a common and representative disease of chronic respiratory tract infection in Japan, characterised by chronic inflammation localised predominantly in the respiratory bronchioles with inflammatory cells such as mononuclear and plasma cells.12 A high number of neutrophils and activated T lymphocytes are also present in the airspaces.13–16 In addition, Pseudomonas aeruginosa and Haemophilus influenzae are often isolated from the sputum of patients with DPB. The pathogenesis of DPB has been elucidated in recent years, but several aspects remain unclear. Recent reports have indicated that the antimicrobial activity of β-defensins may be decreased in cystic fibrosis, which is also characterised by bacterial colonisation and chronic airway infection.9
We have established a sensitive radioimmunoassay (RIA) for HBD-1 and HBD-2.17,18 In the present study we used this assay to measure the concentration of HBD-1 and HBD-2 in plasma and BAL fluid of patients with DPB to investigate their pathophysiological significance in this disease. We also examined immunohistochemically the localisation of HBD-2 in normal lung tissue and lung tissue from a patient with DPB.
METHODS
Study population
The study population consisted of 33 patients with DPB (17 men) and 30 healthy volunteers (15 men). Healthy volunteers (mean (SE) age 37 (3) years) were younger than patients with DPB (55 (2) years). The diagnosis of DPB was based on new criteria published by the Japanese Ministry of Health and Welfare in 199519 and included the following: (1) productive cough and dyspnoea on exertion; (2) rales and rhonchi on physical examination; (3) diffuse disseminated fine nodular shadows, mainly in the lower lung fields on chest radiograph or computed tomography (CT); (4) two abnormalities of forced expiratory volume in 1 second (FEV1) <70% and arterial oxygen tension (Pao2) <10.7 kPa; (5) cold haemagglutination >64; and (6) the presence or history of chronic sinusitis. BAL was performed in 20 of 33 patients with DPB and 10 healthy volunteers. BAL fluid cultures from 20 patients with DPB yielded P aeruginosa in six cases, H influenzae in three cases, H influenzae and S aureus in two cases, H influenzae and Streptococcus pneumoniae in one case, and eight cultures were negative. Mean vital capacity (VC), FEV1, and Pao2 in patients with DPB were 72.0 (4.1)%, 63.5 (2.6)%, and 9.4 (0.3) kPa, respectively. Plasma and BAL fluid sampling was repeated in 14 patients with DPB after approximately 6 months of continuous treatment with macrolide antibiotics. Nine of these patients were treated with erythromycin (600 mg/day) and five with clarithromycin (200 mg/day). None received other antibiotics or corticosteroids during the course of the study. All healthy volunteers had normal chest radiographs, were free of symptoms, and were not taking any medication. The study had local ethical approval and informed consent was obtained from the subjects.
Blood sampling
Blood samples were anticoagulated with ethylenediaminetetraacetic acid sodium and centrifuged to obtain plasma. The plasma (0.3–100 μl) was subjected to RIA for HBD-1 and HBD-2.
Bronchoalveolar lavage
BAL was performed as described previously.15,16 Briefly, after local anaesthesia of the upper airway with 4% lidocaine, a flexible fibreoptic bronchoscope (Olympus BF, type p-20; Olympus Co, Tokyo, Japan) was wedged into a subsegmental bronchus of the right middle lobe. An aliquot of 50 ml sterile saline at body temperature was instilled through the bronchoscope. The fluid was immediately retrieved by gentle suction using a sterile syringe and the procedure was repeated three times. BAL fluid was passed through two sheets of gauze and centrifuged at 500g for 10 minutes at 4°C. The supernatant was analysed for β-defensins by RIA. The remaining cells were counted with a haemocytometer. An aliquot was diluted to a concentration of 2 × 105 cells/ml and a 0.2 ml cell suspension was spun down onto a glass slide at 1100 rpm for 2 minutes using a cytocentrifuge (Cytospin 2; Shandon Instruments; Sewickley, PA, USA). The prepared slides were dried, fixed, and then stained using the May-Giemsa method. More than 200 cells were identified using a photomicroscope.
β-defensin assay
The concentrations of HBD-1 and HBD-2 were measured by the RIA established in our laboratories.17,18 HBD-1 and HBD-2 were synthesised by the solid phase technique. Synthetic HBD-1 and HBD-2 were used for immunising New Zealand white rabbits by multiple intracutaneous and subcutaneous injections. HBD-1 and HBD-2 were radioiodinated and the 125I-labelled peptide was purified by RP-HPLC on a TSK ODS 120A column (Tosoh Co). The incubation buffer for RIA was 50 mM sodium phosphate (pH 7.4) containing 0.25% BSA treated with N-ethylmaleimide, 80 mM NaCl, 25 mM EDTA sodium, 0.05% NaN3, 0.1% Triton X-100, and 3.1% dextran T-40. The diluted sample or a standard peptide solution (100 μl) was incubated for 24 hours with 100 μl diluted antiserum (final dilution, 1:460 000 and 1:4 200 000, respectively). The tracer solution (16 000–18 000 cpm in 100 μl) was added and the mixture incubated for 24 hours, after which normal rabbit serum and anti-rabbit IgG goat serum were added and the whole stored for 16 hours. Bound and free ligands were separated by centrifugation. All procedures were performed at 4°C. Samples were assayed in duplicate.
IL-1β and IL-8 assays
The concentration of interleukin (IL)-1β was measured using a commercially available enzyme linked immunosorbent assay kit (R&D Systems; Minneapolis, MN, USA). The concentration of IL-8 was measured by another commercially available kit (Toray Fuji Bionics, Tokyo, Japan).
Bactericidal assay
The colony count assay20 was used to study the bactericidal activities of HBD-1 and HBD-2 against P aeruginosa (ATCC39324). The bacteria were cultured overnight at 37°C in trypticase soy broth (TSB) and an aliquot of the culture was transferred to fresh TSB and incubated for 2.5 hours at 37°C to obtain mid logarithmic phase cells. The bacteria, precipitated by centrifugation at 800g for 10 minutes, were washed with phosphate buffered saline (PBS) and counted spectrophotometrically at 620 nm. Fifty thousand colony forming units of the bacteria were incubated in 50 μl of incubation medium (Na+ 95 mEq/l, Cl– 90 mEq/l, K+ 25 mEq/l, osmolarity 210 mOsm/l, and pH 7.1) for 2 hours at 37°C with various concentrations of HBD-1 or HBD-2. In the control experiments the bacteria were exposed to 50 μl of incubation medium. The incubation mixtures were diluted serially, spread on nutrient agar plates, incubated for 18 hours at 37°C, and the number of colonies counted. Bactericidal activity was expressed as the colony reduction ratio, representing the percentage of killed to control bacteria.
Immunohistochemistry
Normal lung tissue was obtained at surgery from a 50 year old man with a lung tumour and open lung biopsy specimens were obtained from a 32 year old patient with DPB. The tissues were fixed with 3.7% formaldehyde in 10 mM PBS (pH 7.2), dehydrated in a graded ethanol series, and embedded in paraffin. The cut sections (3 μm thick) were deparaffinised in xylene, rehydrated in a graded ethanol series, then treated with 3% hydrogen peroxide for 30 minutes to inactivate endogenous peroxidases. Non-specific binding was blocked with normal goat serum when assessing HBD-2 immunoreactivity and with normal horse serum when assessing surfactant apoprotein A (SP-A) immunoreactivity. The preparations were incubated overnight at 4°C in 10 mM PBS with anti-HBD-2 antiserum used at a final dilution of 1/2000 or anti-SP-A antiserum (Dako, Glostrup, Denmark) used at a final dilution of 1/100 ENVISION+ (Dako Corporation, USA) as the second antibody for HBD-2 staining and horse biotinylated antimouse IgG (Vectastain, Vector Laboratories Inc, Burlingame, CA, USA) for SP-A staining. The specimens for SP-A staining were allowed to react for 60 minutes with peroxidase conjugated streptavidine (Gibco BRL, Gaithersburg, MD, USA) diluted 1/200 in 10 mM PBS. The specimens for both HBD-2 and SP-A were stained for 3 minutes at room temperature with 20 mg 3,3′-diaminobenzidine tetrahydrochloride (Sigma Chemical Co, St Louis, MO, USA) and 0.006% hydrogen peroxide in 50 mM Tris buffer solution (pH 7.2) and counterstained with haematoxylin. Control studies were performed with normal rabbit serum or anti-HBD-2 antiserum that had been pre-absorbed with 100 μg authentic HBD-2.
Immunoelectron microscopy
Normal lung tissue was fixed with 4% paraformaldehyde in 0.1 M PBS, followed by immunoperoxidase staining with anti-HBD-2 antibody as described above. After colour development the cells were fixed with 2% osmium tetroxide for 1 hour at 4°C. The cells were then dehydrated through a graded series of ethanol and embedded in Epon 812 by placing Epon filled capsules over the tissue sections. After polymerisation, the capsules were removed from the glass slide by heating on a hot plate and a thin layer (1–2 mm) of Epon containing the tissues was cut from each block with a dental drill equipped with a thin blade. The Epon plates were cut into small rectangular pieces (approximately 1 × 2 mm) which were placed onto the top of new Epon blocks so that sections could be cut from the surface. Ultrathin sections were stained with lead citrate for 1 minute and examined with a JEM-100S microscope.
Statistical analysis
Data were expressed as mean (SE). Differences between groups were examined using the Mann-Whitney U test. Correlations between two groups were determined using the Spearman rank correlation analysis. A paired samples Wilcoxon test was used to compare BAL fluid and plasma levels of defensins before and after treatment. A p value of <0.05 indicated statistical significance.
RESULTS
Concentrations of β-defensins in plasma and BAL fluid
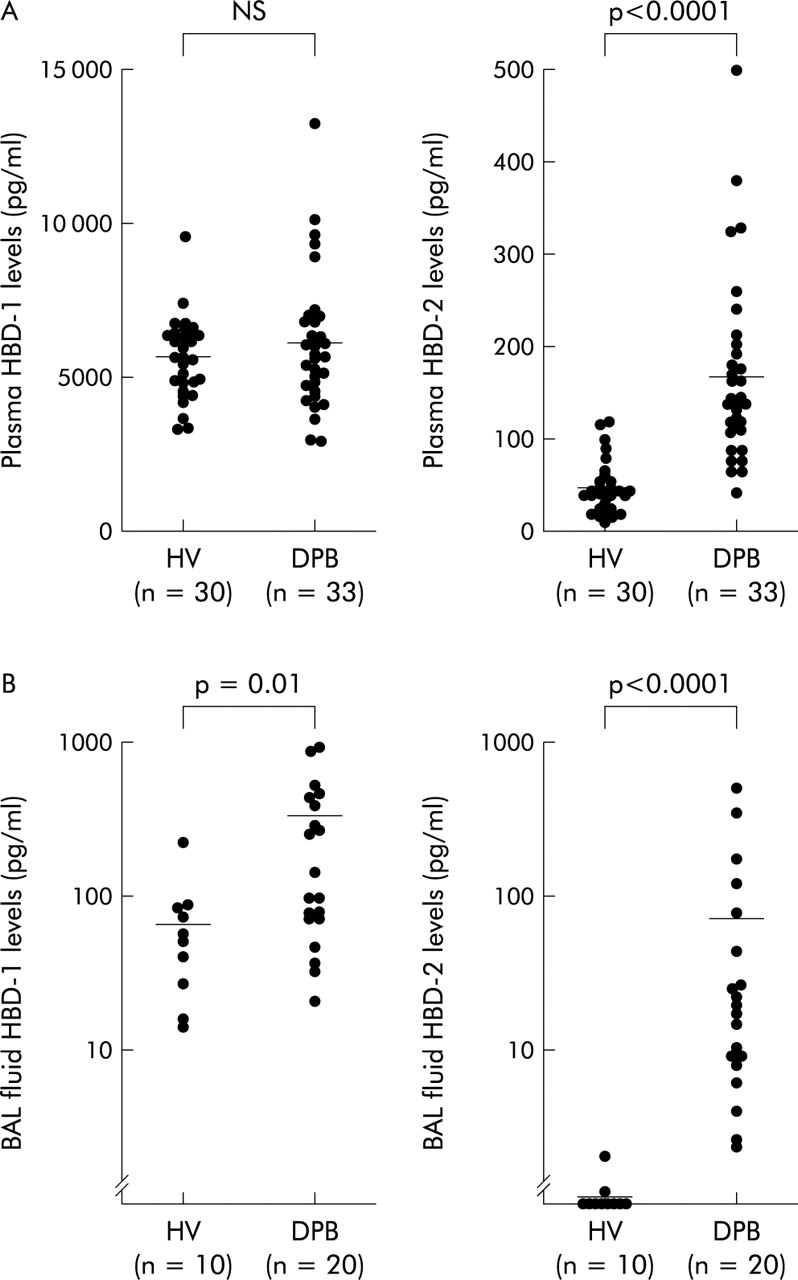
The mean (SE) plasma concentration of HBD-1 in patients with DPB (6166 (418) pg/ml) was not significantly different from that in normal subjects (5654 (241) pg/ml, fig 1A). The plasma concentration of HBD-2 was 3.5-fold higher in patients with DPB (167.6 (18.8) pg/ml) than in normal subjects (47.6 (5.2) pg/ml, p<0.0001; fig 1A). The concentration of HBD-1 in the BAL fluid of patients with DPB was 264.1 (61.0) pg/ml, which was 3.8-fold higher than the concentration in normal subjects (69.2 (20.0) pg/ml, p=0.01, fig 1B). HBD-2 was also detected in the BAL fluid of normal subjects (0.3 (0.2) pg/ml) and at significantly higher concentrations in patients with DPB (71.5 (28.7) pg/ml, p<0.0001; fig 1B). There was a positive correlation between the concentrations of HBD-2 in the plasma and BAL fluid of patients with DPB (r=0.72; p=0.001; fig 2A), while the BAL fluid concentration of HBD-1 did not correlate with the plasma concentration of HBD-1 in these patients.

(A) Individual plasma concentrations and (B) BAL fluid concentrations of HBD-1 (left) and HBD-2 (right) in patients with diffuse panbronchiolitis (DPB) and normal healthy volunteers (HV). The horizontal bars represent mean values.
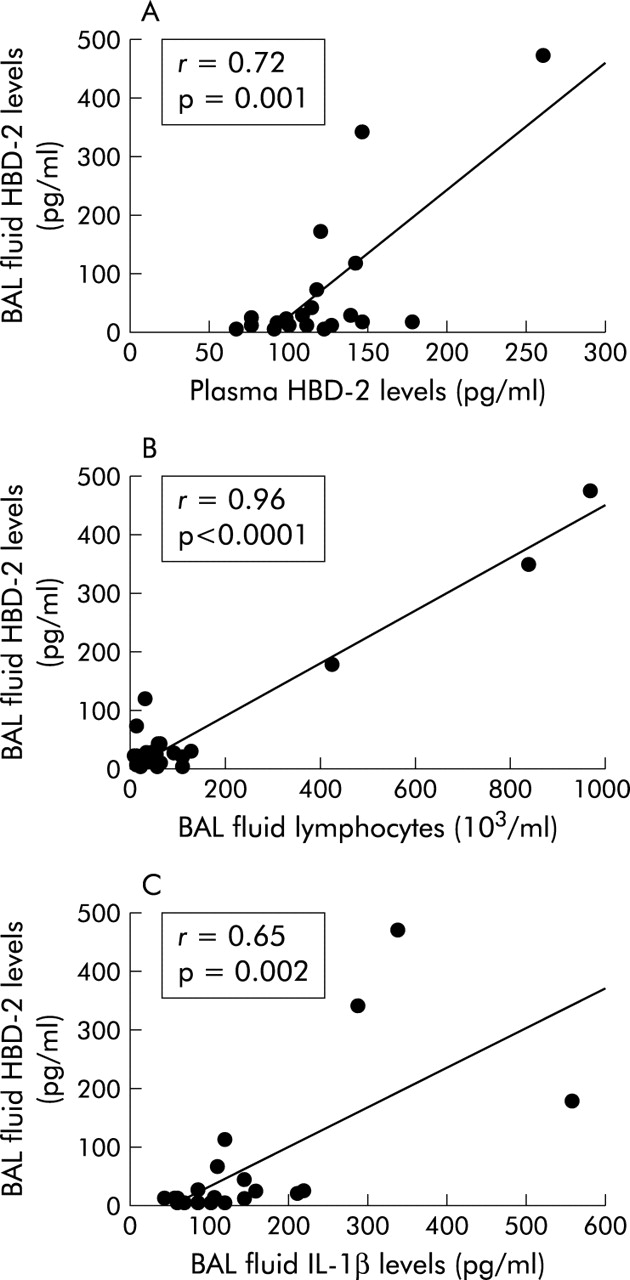
Correlation between (A) HBD-2 concentrations in BAL fluid and plasma of patients with DPB, (B) HBD-2 concentrations and number of lymphocytes in BAL fluid in patients with DPB, and (C) HBD-2 and IL-1β concentrations in BAL fluid of patients with DPB.
Correlation between β-defensins and BAL fluid cells
The total number of cells recovered from BAL fluid of patients with DPB was higher than in the control subjects (table 1). In addition, the relative numbers of neutrophils and lymphocytes in BAL fluid were also significantly higher in DPB patients than in controls. There was a significant correlation between the BAL fluid concentration of HBD-2 and the relative number of neutrophils in BAL fluid in patients with DPB (r=0.85; p<0.0001). The BAL fluid concentration of HBD-2 also correlated with the relative number of lymphocytes in the BAL fluid of patients with DPB (r=0.96; p<0.0001; fig 2B). There was no significant correlation between concentrations of HBD-1 and numbers of cells recovered from the BAL fluid of DPB patients, nor was there a significant relationship between BAL fluid levels of β-defensins in these patients and various laboratory parameters (%VC, FEV1%, FEV1, or Pao2). There was no significant relationship between the type of microorganisms recovered from BAL fluid and the BAL fluid concentrations of HBD-2 in patients with DPB.
BAL fluid cell composition in control subjects and patients with DPB
BAL fluid concentrations of IL-1β and IL-8
The mean (SE) concentrations of IL-1β and IL-8 in BAL fluid of patients with DPB were 155.0 (27.3) pg/ml and 120.7 (56.3) pg/ml, respectively, which were significantly higher than those of control subjects (13.8 (3.6) pg/ml and 3.5 (1.3) pg/ml, respectively; p<0.005). There was a positive correlation between HBD-2 and IL-1β concentrations in BAL fluid (r=0.65; p=0.002; fig 2C), while BAL fluid concentrations of HBD-1 did not correlate with concentrations of IL-1β in the BAL fluid of DPB patients. Neither HBD-1 nor HBD-2 concentrations correlated with IL-8 concentrations in the BAL fluid of patients with DPB.
Effect of treatment with macrolides on the concentration of β-defensins in DPB
The effects of treatment are shown in table 2. Treatment of 14 patients with macrolides significantly improved the results of lung function tests and Pao2. The numbers of total cells, neutrophils, and lymphocytes in BAL fluid also diminished significantly after treatment. Treatment significantly reduced the BAL fluid concentration of HBD-2 but not HBD-1. On the other hand, plasma HBD-1 and HBD-2 concentrations did not change after treatment.
Effect of treatment on lung function tests, Pao2, BAL fluid and plasma variables in patients with DPB
Bactericidal activity of β-defensins
Synthetic HBD-2 (but not HBD-1) exhibited a dose-dependent bactericidal activity against P aeruginosa under conditions that mimicked the bronchial airway surface liquid (fig 3). The concentration for 50% colony reduction of HBD-2 was 17.3 μg/ml. HBD-1 showed no bactericidal activity against P aeruginosa under the experimental conditions.

Bactericidal activities of HBD-1 and HBD-2 against P aeruginosa ATCC39324. The bacteria were incubated with various concentrations of HBD-1 (○) or HBD-2 (•). Bactericidal activity is expressed as the percentage of killed to control bacteria.
Immunohistochemistry of HBD-2
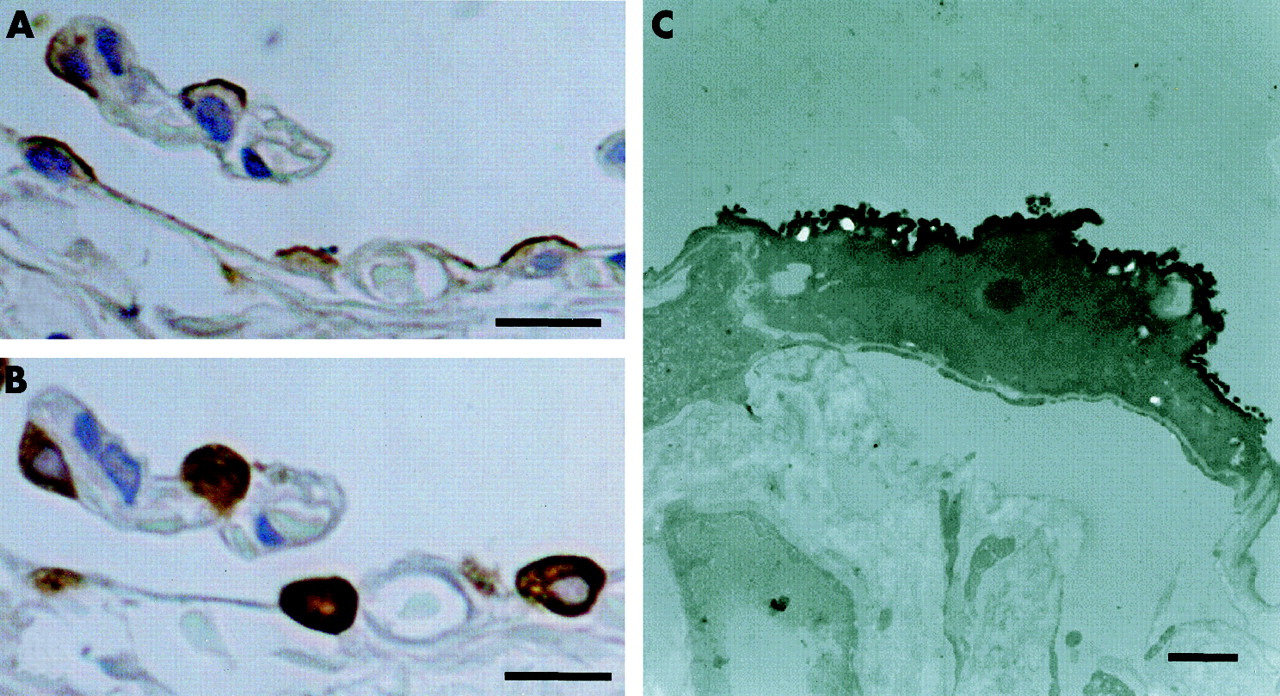
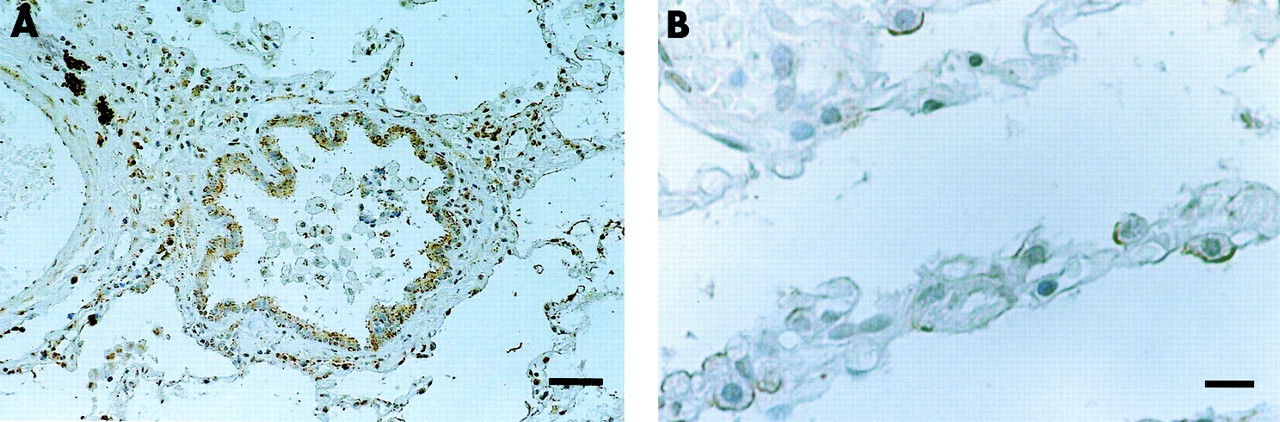
The apical site of alveolar type II cells was immunoreactive for HBD-2 (fig 4A). HBD-2 positive cells in the lung also reacted with SP-A antibody, an immunohistological marker of alveolar type II cells and Clara cells, in serially cut sections (fig 4B). No HBD-2 immunoreactivity was detected in the lung when normal rabbit serum or antiserum absorbed with excessive synthetic HBD-2 was used. Using immunoelectron microscopy, alveolar type II cells that protruded into the alveolar lumen with apical microvilli were immunoreactive for HBD-2 (fig 4C). Immunohistochemistry of open lung biopsy specimens from a patient with DPB showed localisation of HBD-2 in the epithelia of the distal bronchioles and alveolar type II cells (fig 5).

Immunoreactivities for (A) HBD-2 and (B) SP-A in serially cut sections of normal lung tissue. Note the co-localisation of HBD-2 and SP-A in alveolar type II cells. Bar=10 μm. (C) Immunoelectron micrograph showing alveolar type II cells immunoreactive for HBD-2. Bar=1 μm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunohistochemical staining of HBD-2 in open lung biopsy specimen obtained from a patient with DPB. Note the positive staining of (A) bronchiolar epithelial cells and (B) alveolar type II cells. Bar=100 μm (A); 10 μm (B).
DISCUSSION
This study has shown increased BAL fluid concentrations of β-defensins in patients with DPB. In plasma the concentration of HBD-2, but not HBD-1, was also higher in DPB patients than in control subjects. In addition, the plasma concentration of HBD-2 correlated well with the BAL fluid concentration of HBD-2. This is consistent with our previous results showing the presence of high levels of HBD-2, but not HBD-1, in plasma and BAL fluid in patients with Mycobacterium avium intracellulare infection.21 These findings suggest that the increased production of HBD-2 in the pulmonary lesions of patients with DPB may cause high levels of HBD-2 in the plasma. HBD-1 is produced mainly in the kidney and is present in ng/ml concentrations in plasma at a level significantly higher than that in the BAL fluid in both groups of subjects. This may be the reason why the plasma concentration of HBD-1 did not increase in patients with DPB.
Previous in vitro studies have shown that HBD-2 gene expression in primary cultures of airway epithelium is increased following contact with microorganisms or inflammatory mediators such as lipopolysaccharide, tumour necrosis factor-α, and IL-1β, whereas HBD-1 was not.22 The mucoid phenotype of P aeruginosa also induced HBD-2 mRNA in cultured respiratory epithelial cells.23 In our study, high concentrations of IL-1β were detected in the BAL fluid of patients with DPB and a positive correlation was found between IL-1β and HBD-2 concentrations in BAL fluid. These findings confirm that HBD-2 may also be regulated by IL-1β in vivo as well as in vitro, but there was no significant relationship between microorganisms recovered from the BAL fluid of DPB patients and the level of HBD-2 in the BAL fluid.
IL-8 is a potent neutrophil chemotactic factor that enhances transendothelial migration of leucocytes.24 We have previously shown that IL-8 is a major determinant of neutrophil accumulation in the airway lumen in DPB, and its concentration correlates with the concentration of human α-defensins localised in the azurophil granules of neutrophils in BAL fluid of DPB patients.25 In the present study, although a positive correlation was found between the concentration of HBD-2 and the number of neutrophils in the BAL fluid of patients with DPB, there was no significant correlation between IL-8 and HBD-2 concentrations in the BAL fluid of these subjects. It therefore appears that HBD-2 is not directly regulated by IL-8. Patients with DPB are known to have an increased number of lymphocytes in BAL fluid.15 In our study a positive correlation was also found between the concentration of HBD-2 and the number of lymphocytes in the BAL fluid of DPB patients. In this regard, Yang et al26 reported that HBD-2 has chemotactic activity for immature dendritic cells and memory T lymphocytes. β-defensins, particularly HBD-2, may therefore contribute to lymphocyte accumulation in the airway lumen in DPB, although macrophage inflammatory peptide (MIP)-1α is also important for lymphocyte accumulation in the lung.27
The antimicrobial activities of defensins decreased in the presence of high NaCl concentrations. In human bronchial airway surface liquid the Na+ concentration is 80–90 mEq/l, Cl– 70–80 mEq/l, K+ 25–30 mEq/l, osmolarity 250–270 mOsm/l, and pH about 7.0.28 We have reported that both synthetic HBD-1 and HBD-2 showed antimicrobial activity against E coli under conditions nearly the same as in the bronchial airway surface liquid.17,18P aeruginosa is the major pathogen that causes chronic respiratory infection in DPB patients.12 We therefore studied the antimicrobial activities of β-defensins against P aeruginosa under conditions resembling those in the bronchial airway surface liquid. Synthetic HBD-2 showed a dose-dependent antimicrobial activity against P aeruginosa (fig 3) but HBD-1 had no antimicrobial activity against P aeruginosa under the experimental conditions. Considering the antimicrobial activity, HBD-2 seems to function in airway mucosal defence; however, we could not explain the persistent bacterial colonisation of the airway in patients with DPB. An increased level of NaCl has been observed in the airway surface fluid in cystic fibrosis, and it is presumed that the antimicrobial activity of β-defensins decreases as a result of its increased NaCl concentration.9 Although the NaCl concentration in airway surface fluid has not been reported in patients with DPB, it may be also increased in a similar way to cystic fibrosis.
Previous studies of in situ hybridisation histochemistry showed that HBD-2 mRNA is localised in the epithelia of the conducting airway from the proximal bronchi to the distal bronchioles and submucosal glands.10 In our study HBD-2 was detected immunohistochemically in alveolar type II cells in normal lung. No HBD-2 immunoreactivity was seen in the epithelium of the distal bronchioles in normal human lung. Immunohistochemistry of open lung biopsy specimens from a patient with DPB showed localisation of HBD-2 in the epithelia of the distal bronchioles, where HBD-2 mRNA was detected by in situ hybridisation.10 While our immunohistochemical study demonstrated the augmentation of HBD-2 synthesis in the epithelium of the distal bronchioles in DPB, HBD-2 immunoreactivity in alveolar type II cells in the lungs of these patients did not differ from that in normal lungs. These results do not exclude the possibility that HBD-2 peptide is not synthesised in alveolar type II cells but is deposited on alveolar type II cells. Further studies using cultured alveolar type II cells are needed to confirm HBD-2 gene expression in alveolar type II cells.
The use of low dose, long term macrolide antibiotics has been established as an effective treatment for DPB.29 In the present study a significant reduction in the BAL fluid concentration of HBD-2 was observed in DPB patients after treatment with macrolide antibiotics. This reduction may be due to an improvement in the chronic inflammation in the respiratory tract. Alternatively, the reduction might be due to a direct inhibition of HBD-2 synthesis in respiratory epithelial cells induced by macrolide antibiotics. Whichever the mechanism, our results suggest that the BAL fluid concentration of HBD-2 reflects the degree of airway inflammation in DPB.
In conclusion, we have demonstrated the presence of high BAL fluid levels of β-defensins in the respiratory tract of patients with DPB. Our results also suggest that β-defensins, particularly HBD-2, may participate in antimicrobial defence in the respiratory tract in these patients, and that the BAL fluid concentration of HBD-2 may be a useful marker of airway inflammation in this disease.
Acknowledgments
The authors thank S Tajiri for the excellent technical assistance. This study was supported in part by the 21st COE Program and a research grant from the Ministry of Education, Science, Sports, and Culture of Japan.
REFERENCES
Linked Articles
- Airwaves