Article Text

Abstract
Background: Exposure of patients with atopic asthma to allergens produces a long term increase in exhaled nitric oxide (FENO), probably reflecting inducible NO synthase (NOS) expression. In contrast, bradykinin (BK) rapidly reduces FENO. It is unknown whether BK suppresses increased FENO production after allergen exposure in asthma, and whether it modulates FENO via NOS inhibition.
Methods: Levels of FENO in response to aerosolised BK were studied before (day 3) and 48 hours after (day 10) randomised diluent (diluent/placebo/BK (Dil/P/BK)), allergen (allergen/placebo/BK (All/P/BK), and allergen/l-NMMA/BK (All/L/BK)) challenges (day 8) in 10 atopic, steroid naïve, mild asthmatic patients with dual responses to inhaled house dust mite extract. To determine whether BK modulates FENO via NOS inhibition, subjects performed pre- and post-allergen BK challenges after pretreatment with the NOS inhibitor l-NMMA in the All/L/BK period.
Results: Allergen induced a fall in FENO during the early asthmatic reaction (EAR) expressed as AUC0–1 (ANOVA, p=0.04), which was followed by a rise in FENO during the late asthmatic reaction (LAR) expressed as AUC1–48 (ANOVA, p=0.008). In the Dil/P/BK period, FENO levels after BK on pre- and post-diluent days were lower than FENO levels after placebo (difference 23.5 ppb (95% CI 6.2 to 40.9) and 22.5 ppb (95% CI 7.3 to 37.7), respectively; p<0.05). Despite the long lasting increase in FENO following allergen challenge in the LAR, BK suppressed FENO levels at 48 hours after allergen challenge in the All/P/BK period, lowering the increased FENO (difference from placebo 54.3 ppb (95% CI 23.8 to 84.8); p=0.003) to the baseline level on the pre-allergen day (p=0.51). FENO levels were lower after l-NMMA than after placebo on pre-allergen (difference 10.85 ppb (95% CI 1.3 to 20.4); p=0.03) and post-allergen (difference 36.2 ppb (95% CI 5.5 to 66.9); p=0.03) days in the All/L/BK and All/P/BK periods, respectively. l-NMMA did not significantly potentiate the pre- and post-allergen reduction in BK induced FENO.
Conclusions: Bradykinin suppresses the allergen induced increase in exhaled NO in asthma; this is not potentiated by l-NMMA. Bradykinin and l-NMMA may follow a common pathway in reducing increased NO production before and after experimental allergen exposure. Reinforcement of this endogenous protective mechanism should be considered as a therapeutic target in asthma.
- asthma
- exhaled nitric oxide
- allergen induced exacerbation
- bradykinin
Statistics from Altmetric.com
Nitric oxide (NO) is generated from l-arginine by the enzyme NO synthase (NOS), of which constitutive (cNOS) and inducible (iNOS) isoforms have been described functionally.1 Both are present in the airways.2 cNOS (endothelial NOS (eNOS) and neuronal NOS (nNOS)) release small amounts of NO (in the picomolar range) within seconds in response to increases in cytosolic Ca2+. Several agonists, including bradykinin, activate cNOS upon receptor stimulation.3 In contrast, the expression of iNOS is induced by proinflammatory cytokines, a process that results in a protracted and Ca2+ independent release of high levels of NO (in the nanomolar range)4 which is inhibited by corticosteroids.5,6 NO formed by cNOS leads to cGMP dependent airway smooth muscle relaxation, whereas NO released by iNOS is associated with proinflammatory effects.7,8 Both enzyme isoforms are likely to play a role in asthma.7,8 NO is detectable in the exhaled air of healthy volunteers9 and is increased in patients with atopic asthma.10 It has recently been shown that the increased levels of NO in the exhaled air of asthmatic patients are related to airway eosinophilic inflammation11 and to the increased expression of iNOS.6 Exhaled NO (FENO) is inhibited by treatment with inhaled corticosteroids5 and NOS inhibitors,12 suggesting that it is sensitive to steroids and produced by the enzymatic NOS pathway.
In most sensitised asthmatics, allergen exposure induces an early (EAR) and late (LAR) asthmatic response. The EAR, with early onset (10 minutes after exposure) and short duration (1–2 hours), is followed by the LAR (3–10 hours) which is characterised by bronchoconstriction associated with airway cellular inflammation.13 Interestingly, it has been shown that the LAR is associated with increased levels of NO in the exhaled air,14 suggesting that FENO may reflect allergic inflammation in asthmatic airways.
Bradykinin is an autacoid involved in the pathophysiology of asthma.15 Bradykinin induced hyperresponsiveness is increased by NOS inhibitors in asthma,16 suggesting that bradykinin releases Ca2+ dependent cNOS derived NO in the airways.16 Conversely, inhalation of bradykinin,17 as well as of prostaglandin E2 (PGE2),18 reduces FENO in stable asthmatics, which suggests that bradykinin has a role in modulating FENO in the absence of disease activity. However, it is unknown whether bradykinin can regulate increased FENO levels during late phase inflammation induced by allergen in asthma and whether it can modulate FENO via NOS inhibition.
We measured the effect of bradykinin inhalation on NO levels in the exhaled air of asthmatic patients, both at baseline and after experimental allergen exposure in the absence or presence of the NOS inhibitor l-NMMA. This study is part of a larger project for which the study design, methods, and results for other outcome parameters have already been published.16
METHODS
Subjects
Ten non-smoking patients (five men) with mild intermittent asthma and house dust mite atopy,19 as described elsewhere,16 participated in the study (table 1⇓). All subjects were symptom free at the time of the study and were not on regular medication. They had normal lung function and were hyperresponsive to inhaled histamine.20 Inhaled short acting β2 agonists were allowed on demand until 12 hours before testing. The subjects had documented EAR and LAR to inhaled house dust mite extract in the screening period.20 The study was approved by the ethics committee of the Leiden University Medical Centre and all participants gave written informed consent.
Characteristics of participants
Study design
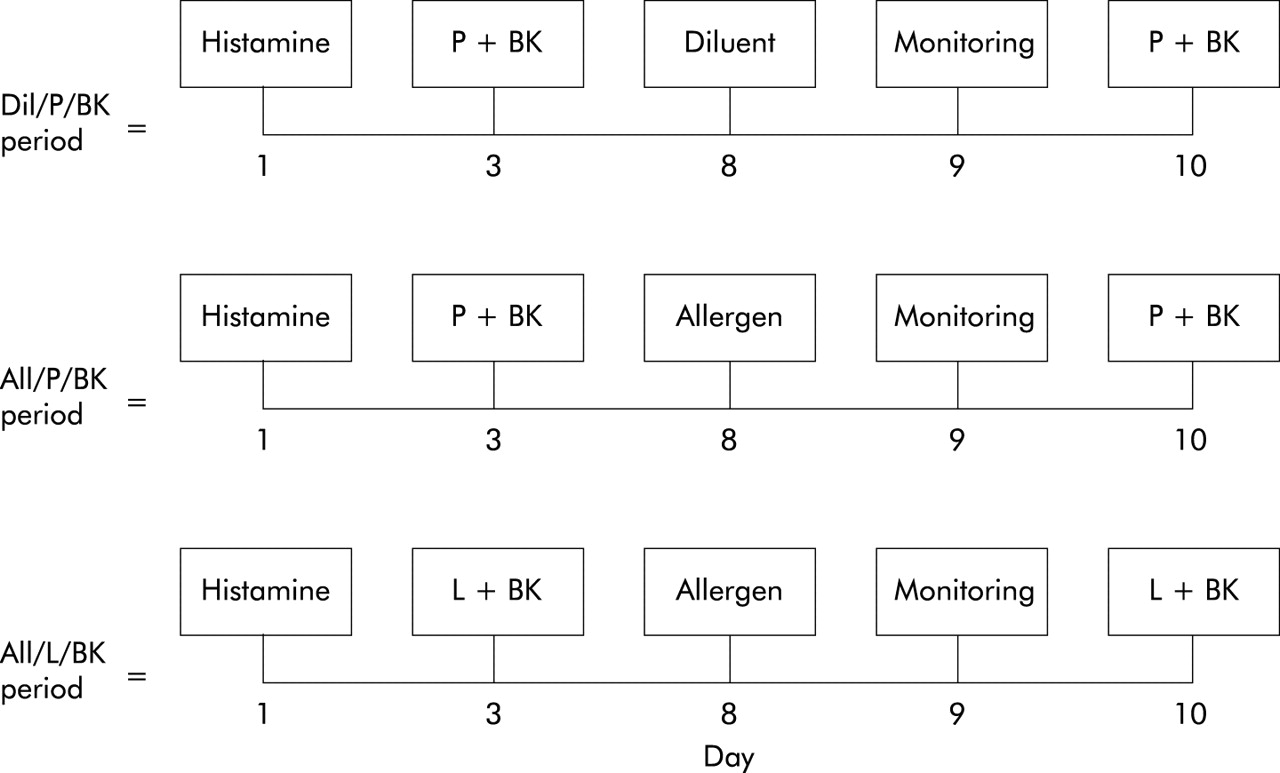
The screening period for the selection criteria has been described previously.16 The study had a randomised, placebo controlled, crossover design and consisted of three periods (diluent/placebo/bradykinin (Dil/P/BK), allergen/placebo/bradykinin (All/P/BK), and allergen/l-NMMA/bradykinin (All/L/BK)) of five study days each, separated by a washout interval of 2–4 weeks. During each study period the participants pretreated with placebo (Dil/P/BK and All/P/BK periods) or l-NMMA (All/L/BK period) underwent a bradykinin challenge before (day 3) and 48 hours after (day 10) the diluent (Dil/P/BK period) or allergen challenge (All/P/BK and All/L/BK periods, day 8; fig 1⇓). On day 9 the participants underwent spirometric tests and FENO monitoring (fig 1⇓). On day 1 of each study period a control PC20 histamine measurement was performed.

Study design. Dil=diluent; All=allergen; P=placebo; BK=bradykinin; L=NG-monomethyl-l-arginine.
FENO was measured (2 minutes after spirometry) at baseline, after placebo or l-NMMA, and at the end of bradykinin (after 5 minutes and every 10 minutes up to 1 hour) and allergen/diluent challenge (every 10 minutes in the first hour, every 30 minutes in the second hour, every 60 minutes up to 10 hours, and at 24 hours), when a fall in forced expiratory volume in 1 second (FEV1) of ⩾20% was achieved or after three consecutive diluent inhalations.
Inhalation challenges
Freshly prepared NG-monomethyl-l-arginine (l-NMMA; Clinalfa AG, Läufelfingen, Switzerland: 10 mg in 3 ml 0.9% saline) or placebo (3 ml 0.9% saline), bradykinin (Clinalfa AG, Läufelfingen, Switzerland: 0.0024–5.0 mg/ml), diluent, and allergen extract of Dermatophagoides pteronyssinus (SQ 503; Vivodiagnost, ALK, Benelux) were inhaled as described previously.16 FEV1 measurements for bradykinin and allergen or diluent challenges were performed as previously reported.16
Measurements of exhaled NO
FENO measurements were performed, according to the present recommendation,21 using a Sievers NOA 270B chemiluminescence analyser (Sievers, Boulder, CO, USA) as previously described.22 Subjects performed a slow vital capacity manoeuvre with a constant expiratory flow of 100 ml/s using online visual monitoring. An expiratory resistance of 5 cm H2O was applied to prevent bias of the measurement with nasal NO. Plateau levels of NO were determined and expressed as parts per billion (ppb). Subjects inspired “NO free” air (<1 ppb) during measurements. Three successive recordings were made at 1 minute intervals and the mean was used in the analysis.
Statistical analysis
The difference in FENO after allergen challenge was expressed in absolute terms as the area under the time-response curve (AUC) from 0 to 48 hours (total AUC0–48/48=mean FENO), from 0 to 1 hour (AUC0–1) in the EAR, and from 1 to 48 hours (AUC1–48) in the LAR. The effect of bradykinin or l-NMMA on FENO was calculated in terms of the difference between treatments or the difference from placebo with 95% confidence intervals for these differences.
Values in the text and figures are expressed as mean (SE) or 95% confidence intervals (95% CI). In order to measure the effect of allergen and diluent on FENO we compared AUC0–1, AUC1–48, and total AUC0–48/48 using one way analysis of variance (ANOVA). Repeated measures ANOVA was applied to test whether there were any differences in baseline FENO on different days and in FENO between treatments. Two tailed paired Student’s t tests were applied to explore the differences. Repeated measures ANOVA of PC20 histamine and FEV1 values not described in the previous report16 were performed. In all cases a p value of <0.05 was considered significant.
Statistical analyses were performed using Statistica for Windows (StatSoft Inc; Tulsa, OK, USA). A power calculation was based on the standard deviation (SD) of the differences between the bradykinin induced reduction in FENO during placebo pretreatment in the Dil/P/BK (day 3) and All/P/BK (day 3) periods. Based on this SD (13.6% change in FENO), the current sample size allows us to detect a difference of 13.6% in FENO (1 SD) between repeated bradykinin challenges (power 0.8, α 0.05).
RESULTS
Airway responses
PC20 histamine was similar in each study phase (p=0.28). Airway responses to placebo of challenge with l-NMMA (0.9% saline) or l-NMMA (10 mg in 3 ml) plus diluent or allergen challenge have been reported previously.16 At 24 and 48 hours after allergen challenge FEV1 was similar to baseline in both the All/P/BK (p=0.57) and All/L/BK periods (p=0.23). Bradykinin challenge before and after diluent or allergen provoked a similar percentage reduction in FEV1 in the Dil/P/BK period (26 (2)% and 25 (2)%), All/P/BK period (27 (2)% and 28 (2)%), and All/L/BK period (26 (2)% and 28 (2)%), respectively (p=0.44).
Exhaled NO
NO was detectable in the exhaled air of all asthmatic subjects. Baseline FENO values in the Dil/P/BK period (day 3: 37 (11) ppb; day 8: 38 (12) ppb), All/P/BK period (day 3: 35 (9) ppb; day 8: 33 (8) ppb), and All/L/BK period (day 3: 37 (10) ppb; day 8: 31 (8) ppb) were not different (MANOVA, p=0.48).
There was no change in FENO up to 48 hours after diluent challenge (day 8), whereas allergen challenge on day 8 reduced FENO in the first hour after the end of the challenge in the All/P/BK and All/L/BK periods (fig 2⇓), as shown by the significant differences between AUC0–1 for FENO in the All/P/BK period (19.9 ppb/h (95% CI 9.2 to 30.7)), the All/L/BK period (17.9 ppb/h (95% CI 8.5 to 27.4)), and the Dil/P/BK period (38.1 ppb/h (95% CI 9.6 to 66.6); p=0.04). FENO was increased in the LAR after allergen challenge in the All/P/BK and All/L/BK periods (fig 2⇓). AUC1–48 for FENO on day 8 was significantly different in the All/P/BK (3282 ppb/h (95% CI 1693 to 4870)), All/L/BK (2579 ppb/h (95% CI 1467 to 3690)), and Dil/P/BK periods (1781 ppb/h (95% CI 471 to 3091); p=0.008). The differences in total AUC0–48/48 for FENO, interpreted as the mean FENO, on day 8 in the All/P/BK (68.8 ppb (95% CI 35 to 102)), All/L/BK (54.1 ppb (95% CI 31 to 77)), and Dil/P/BK periods (37.8 ppb (95% CI 10 to 65)) were significant (p=0.009). The differences in all the AUCs for FENO between diluent and the two allergen challenges are shown in table 2⇓.
Mean (95% CI) differences in AUC0–1, AUC1–48, and total AUC/48 for FENO between diluent (Dil/P/BK) and allergen (All/P/BK and All/L/BK) challenges
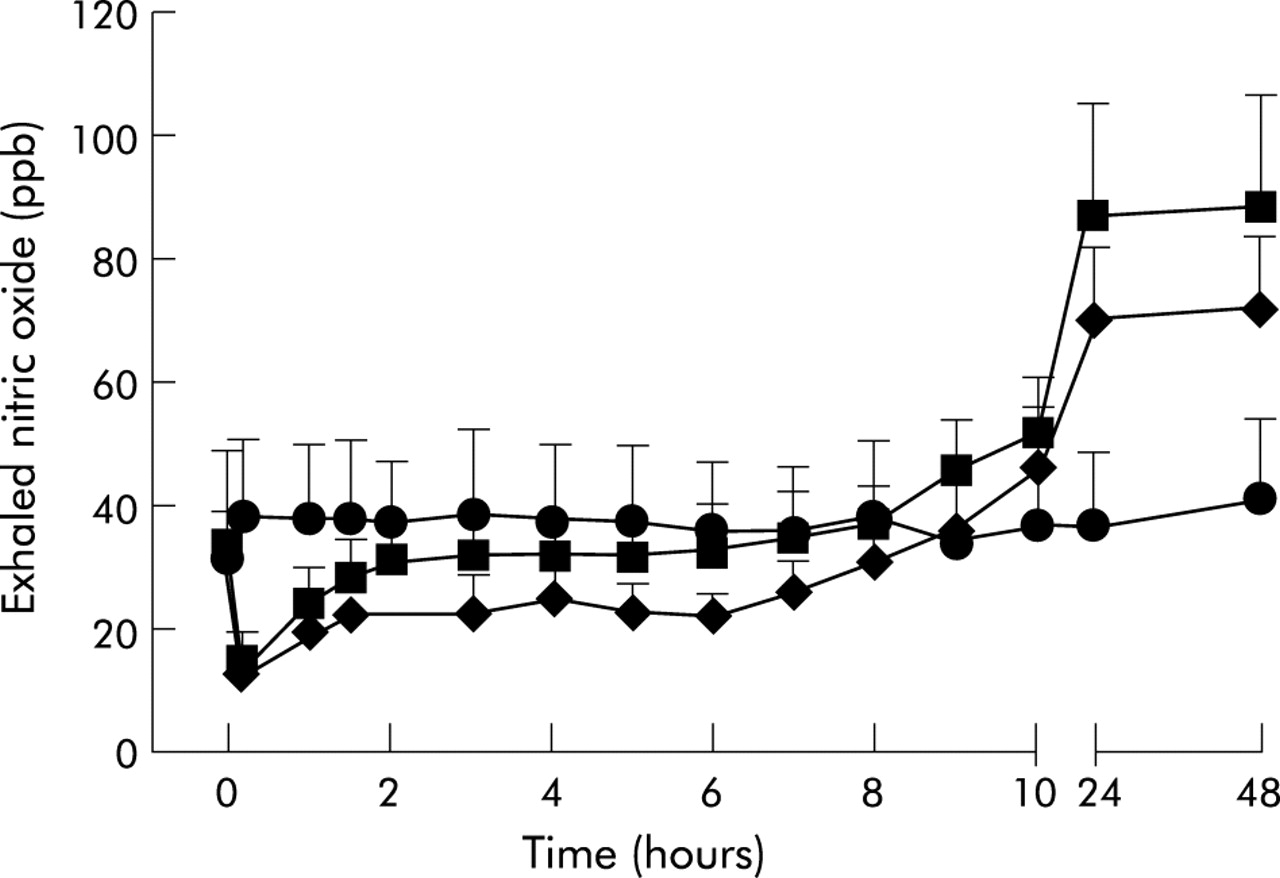
Time course of the change in exhaled nitric oxide levels after diluent (circles) and allergen (allergen/placebo/bradykinin: squares; allergen/l-NMMA/bradykinin: rhombus) challenges in 10 atopic patients with mild asthma. Mean (SE) values are shown.
In the Dil/P/BK period FENO values were significantly lower after bradykinin than after placebo (fig 3⇓) on day 3 (difference 23.5 ppb (95% CI 6.2 to 40.9); p=0.01) and day 10 (difference 22.5 ppb (95% CI 7.3 to 37.7); p=0.008). Both FENO values after bradykinin in the Dil/P/BK period were not significantly different from the corresponding FENO value on the pre-allergen day in the All/P/BK period (p=0.15). Forty eight hours after allergen challenge in the All/P/BK period bradykinin completely suppressed the increased FENO levels (fig 3⇓), as shown by the reduction in the allergen induced increase in FENO (difference from placebo: 54.3 ppb (95% CI 23.8 to 84.8); p=0.003) to the baseline level of the pre-allergen day (p=0.51). The difference between the reductions in FENO induced by bradykinin from the respective placebo 48 hours after allergen in the All/P/BK period and after diluent in the Dil/P/BK period was statistically significant (31.8 ppb (95% CI 7.1 to 56.5); p=0.017).

Effect of bradykinin challenge on exhaled nitric oxide in (A) the diluent/placebo/bradykinin period, (B) the allergen/placebo/bradykinin period, and (C) the allergen/l-NMMA/bradykinin period. Exhaled nitric oxide levels were measured before pre-treatment (open columns), after pretreatment (with placebo or l-NMMA: shaded columns), and 5 minutes after bradykinin challenge (filled columns) on days 3 and 10 of each study period. On day 8 diluent (diluent/placebo/bradykinin) and allergen (allergen/placebo/bradykinin and allergen/l-NMMA/bradykinin) challenges were performed. Mean (SE) values are shown. * p<0.05 v respective placebo or l-NMMA.
In the Dil/P/BK and All/P/BK periods, neither placebo nor l-NMMA (0.9% saline) changed baseline FENO on any study days (fig 3⇑). In the All/L/BK period the differences between the level of FENO after l-NMMA and placebo before and after allergen were significant (All/L/BK: 10.85 ppb (95% CI 1.3 to 20.4), p=0.029; All/P/BK: 36.2 ppb (95% CI 5.5 to 66.9), p=0.026).
After pretreatment with l-NMMA, bradykinin significantly reduced FENO on pre-allergen (difference from l-NMMA: 14.1 ppb (95% CI 6.8 to 21.4); p=0.002) and post-allergen (difference from l-NMMA: 29.7 ppb (95% CI 16 to 43.5); p=0.0008) days (fig 3⇑). The differences between the level of FENO after placebo plus bradykinin in the All/P/BK period and after l-NMMA plus bradykinin in the All/L/BK period on pre- and post-allergen days (1.96 ppb (95% CI −2.1 to 6), p=0.3; and 9.4 ppb (95% CI −0.5 to 19.5), p=0.06, respectively) were not statistically significant. The difference between the baseline values of FENO (48 hours after allergen) on the post-allergen day (day 10) in the All/P/BK and All/L/BK periods was 15.8 ppb (95% CI −5 to 36.4); p=0.11), which explains the slight difference between FENO after placebo plus bradykinin in the All/P/BK period and after l-NMMA plus bradykinin in the All/L/BK period on the post-allergen day.
On all bradykinin treatment days the recovery time (up to 60 minutes) was characterised by a complete spontaneous recovery in FEV1 values to baseline levels, whereas FENO levels remained significantly lower than baseline (p<0.05, fig 4⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Time course of the change in exhaled nitric oxide (rhombus) and forced expiratory volume in one second (FEV1; squares) during the recovery period after diluent in (A) the diluent/placebo/bradykinin period and after allergen in (B) the allergen/placebo/bradykinin period and (C) the allergen/l-NMMA/bradykinin period on the day of bradykinin challenge (day 10) in 10 atopic patients with mild asthma. Mean (SE) values are shown.
DISCUSSION
This study shows, for the first time, that the allergen induced increase in FENO in atopic asthma is suppressed by bradykinin, and that the NOS inhibitor l-NMMA significantly reduces pre- and post-allergen FENO levels and does not potentiate the bradykinin induced reduction in FENO. Taken together, our results indicate that bradykinin, via NOS inhibition, modulates the prolonged increase in FENO induced by allergen exposure. Our findings suggest that bradykinin inhibits excessive NO production resulting from an overexpression of the NOS pathway in asthmatic airways during an exacerbation.
The bradykinin induced fall in FENO levels in stable asthmatics has recently been reported by Kharitonov et al.17. We have shown that bradykinin suppresses increased levels of FENO 48 hours after allergen challenge, which suggests that it has a role in modulating exacerbations of allergic inflammation. Furthermore, the finding that FENO levels were still significantly reduced 60 minutes after the end of the pre-allergen and post-allergen bradykinin challenge, when FEV1 had completely recovered, suggests that the rapid suppression of FENO by bradykinin has a relatively long duration and is not dependent on airway calibre. One possible explanation for the action of bradykinin is vasodilation15 with potentially enhanced NO trapping by haemoglobin23 and/or altered diffusion of NO to the airway lumen due to mucosal swelling and gland secretion15 with subsequent airway obstruction; however, the short term effect of vasodilation and the reduction in FENO even after recovery of the obstruction limit the validity of this hypothesis.
In this study we also examined whether bradykinin negatively modulates FENO through inhibition of NOS. l-NMMA has previously been found to reduce FENO levels significantly in stable asthma,12 so we studied the effect of l-NMMA on the bradykinin induced reduction in FENO. The lack of significant potentiation by l-NMMA on the bradykinin induced reduction in FENO suggests that bradykinin and l-NMMA share a common pathway in inhibiting FENO production in the airways of patients with asthma. It has recently been found that prostaglandins also reduce FENO in asthmatic subjects.18 Bradykinin stimulates PGE2 release from airway epithelial cells, either by the constitutive isoform of cyclo-oxygenase enzyme (COX-1) or by the inducible isoform (COX-2),24 and PGE2 negatively modulates the induction of iNOS expression at the transcriptional level.25 It is therefore possible that the reduction in FENO induced by bradykinin is a result of the inhibition of iNOS expression through the release of PGE2. However, the rapid reduction in FENO observed after bradykinin inhalation does not favour such a delayed mechanism, but suggests a possible direct or indirect influence of bradykinin on NOS at the post-transcriptional level. It has previously been shown that high levels of NO can downregulate NOS pathways by negative feedback,26 and also that augmented production of FENO in the lower respiratory tract of eNOS deficient mice is associated with pulmonary iNOS overexpression, indicating a counter-regulation between the two NOS isoforms.27 Furthermore, a recent study in human mesangial cells showed a negative post-transcriptional regulation of iNOS by the NO/cGMP pathway.28 On the basis of the present results we believe that bradykinin is not only an activator of the Ca2+ dependent cNOS/cGMP pathway which modulates airway hyperresponsiveness,29 but also may downregulate excessive NO release, probably derived by iNOS, through negative feedback in allergic asthma.
In line with this hypothesis, we have previously shown that iNOS (but not eNOS or nNOS) immunostaining is higher after allergen exposure in the epithelium of bronchial biopsy sections of subjects with asthma,16 which suggests two major findings on the basis of the present data: (1) iNOS may be responsible for the increase in FENO after allergen; (2) bradykinin should affect iNOS activity by reducing FENO in asthmatic patients and, in particular, by suppressing the increase in FENO after exposure to allergen.
In the present study, allergen inhalation led to two opposing changes in FENO levels in atopic mild asthmatics. Exhaled NO tended to decrease during the EAR and exhibited a prolonged increase during and after the LAR. These findings are consistent with previous data showing the effect of allergen on FENO levels in atopic asthmatic patients with LAR.14 In the early phase we found a reduction in FENO levels compared with baseline, which confirms previous observations.30 In previous studies acute airways obstruction has been found to have a partial effect on FENO levels.31,32 It is therefore possible that bronchoconstriction could, at least partially, lead to a fall in FENO levels during the early phase of the bronchial allergic reaction, possibly as a result of limiting NO diffusion or increased airflow velocity in constricted airways. We also postulate an indirect effect of allergen on FENO levels during the early phase reaction via the release of proinflammatory mediators such as kinins or prostaglandins.17,18
Exhaled NO appeared to increase 9–48 hours after allergen inhalation, confirming a previous report.14 In the latter study the authors observed a decline in FENO by 27 hours. The longer duration of increased FENO in our study could be a reflection of a possibly higher level of allergic inflammation induced by allergen exposure. Several lines of evidence suggest that FENO reflects iNOS expression in asthma,5,6 which recognises that FENO is a non-invasive marker of airway inflammation in asthma.33 The delayed and long term increase in FENO during the LAR could be explained by the action of proinflammatory cytokines released from inflammatory cells recruited to the airways after allergen exposure, and of reactive oxygen species with subsequent upregulation of iNOS expression.34,35 Furthermore, an additional explanation could be derived from the recent evidence that airway acidification converts nitrite (NO2−) to NO gas in quantities consistent with those observed in expired air during an asthma exacerbation,36 or after the LAR to allergen.
To avoid differences in FENO as a result of smoking or medication use, we selected a homogeneous group of non-smoking, atopic, mild intermittent asthmatic subjects who were not using any steroid medication before or during the experiments. It has recently been shown that repeated forced vital capacity (FVC) manoeuvres significantly reduce FENO levels in healthy and asthmatic subjects by 13% and 10%, respectively.37 Even though our FENO measurements were performed 2 minutes after an FVC manoeuvre, we do not consider that it had any effect because no effect on the FENO measurement was seen following diluent.
NO released by iNOS is likely to exert proinflammatory activities such as induction of eosinophil chemotaxis and activation of the Th2 driven immune response,8,11 as observed during the late phase response to inhaled allergen. The bradykinin induced suppression of raised FENO levels during the late phase suggests that bradykinin can modulate allergic inflammation exacerbations in the airways by the inhibition of proinflammatory NOS. On the basis of our study, a clinically relevant scenario could be outlined in patients with asthma where an acute inflammatory mediator (bradykinin) could have a modulatory role in limiting the effect of a detrimental long lasting inflammatory pathway (iNOS). The reinforcement of such endogenous protective mechanisms should be considered as a potential therapeutic target in asthma.
Acknowledgments
This study was supported by a Research Fellowship of the European Respiratory Society (ERS).
REFERENCES
Linked Articles
- Airwaves