Article Text

Abstract
Background: The airway cilia of patients with primary ciliary dyskinesia (PCD) exhibit several anomalies when studied by transmission electron microscopy, but little is known about the ultrastructural organisation of ciliary membranes in these patients. Freeze fracture replication of airway epithelium from patients with PCD provides a means of achieving high resolution views of cell membrane structure. Ciliary necklaces are a specialised structural feature of ciliary membranes thought to serve as a timing mechanism for ciliary beat, and their characterisation in the cilia of patients with PCD may contribute new insights into the pathophysiology of this syndrome.
Methods: The nasal epithelium of three patients with PCD was freeze fractured and replicated with platinum and carbon shadowing. The resultant preparations were examined by transmission electron microscopy and the ciliary necklaces were compared with similar preparations of nasal biopsy specimens from normal healthy subjects.
Results: The ciliary necklaces of the three patients with PCD were normal with no overt differences from those of healthy individuals.
Conclusions: The defective ciliary motility observed in patients with PCD does not appear to result from membrane dysfunction associated with overt disorganisation of ciliary necklace structure.
- primary ciliary dyskinesia
- freeze fracture
- ciliary necklace
Statistics from Altmetric.com
Sections of airway cilia from patients with primary ciliary dyskinesia (PCD) exhibit several anomalies when viewed by transmission electron microscopy.1–3 However, little is known about the ultrastructural organisation of ciliary membranes in these individuals. PCD is a rare genetic disorder characterised by hereditary structural defects in the cilia which affects one in 20 000 individuals.4 It was initially described by Siewert in 1904 as a triad of symptoms characterised by situs inversus, sinusitis, and bronchiectasis.5 At present PCD is characterised clinically by recurrent sinopulmonary infection, bronchiectasis, subfertility, and situs inversus4 and confirmed by laboratory documentation of ciliary dyskinesia and ultrastructural defects.
A normal cilium exhibits a core axoneme comprising nine peripheral doublets of microtubules surrounding a central pair of microtubules. The orientation of the central microtubular pair reflects the coordinated movement of the cilia. There are also several microtubule associated proteins (MAPs) which play important structural and functional roles. Dynein arms were among the first MAPs identified.6 Outer dynein arms are thought to regulate beat frequency and inner arms to regulate beat waveform.6 Radial spokes comprise another type of MAP that connect each doublet to the central microtubular pair.
Among the ultrastructural abnormalities observed in the cilia of patients with PCD are absent or reduced inner and outer dynein arms,1 absent radial spokes,2 an absent central microtubular pair replaced by a peripheral microtubule doublet (transposition),3 and ciliary aplasia with defects in ciliary orientation.7
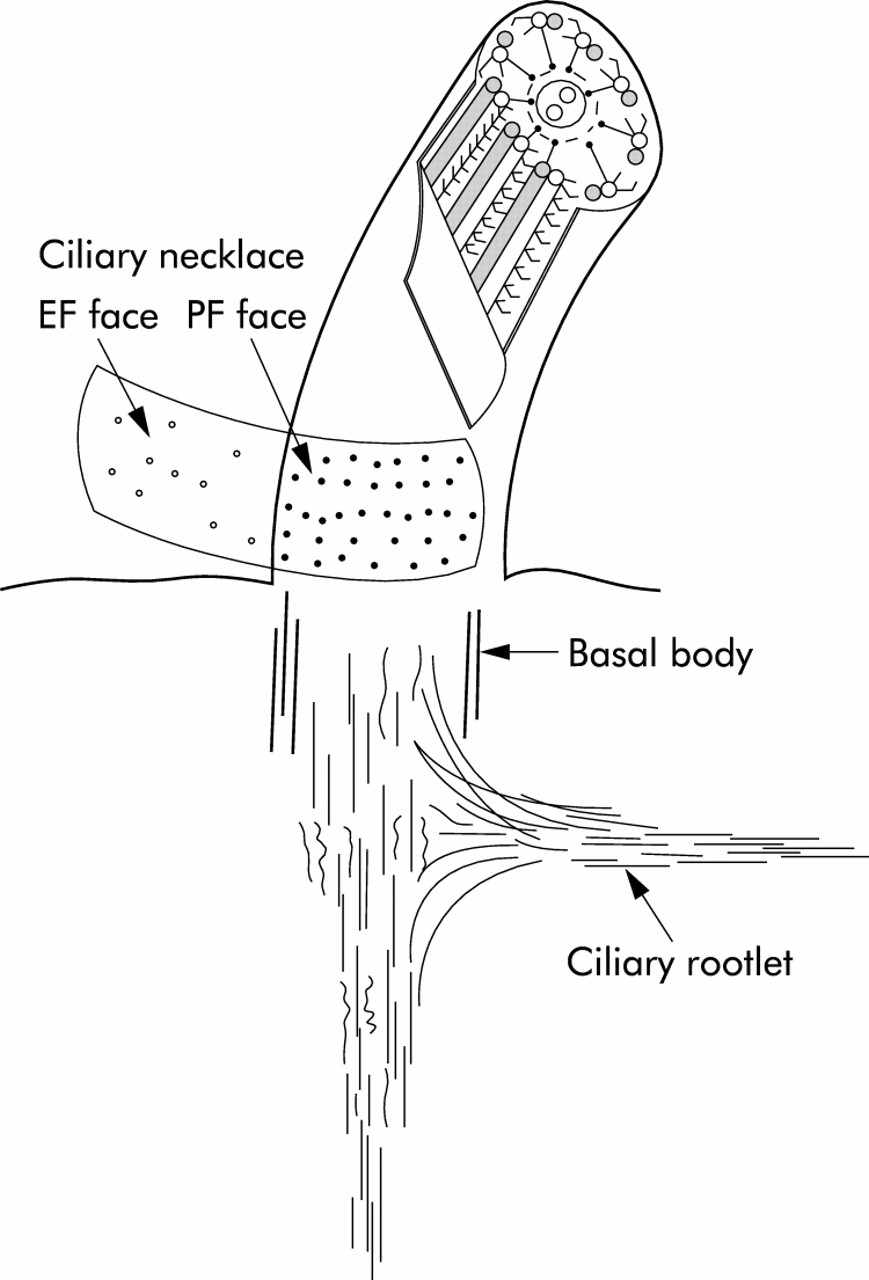
The ciliary necklace is a complex of particles found at the base of each cilium (fig 1). The particles are generally arranged in scalloped rows about 30 nm apart with each scallop corresponding to a peripheral microtubule.8 The number of necklace strands varies among species, with 4–6 strands usually present in human cilia.8 The ciliary necklace has been proposed as a membrane component that functions in energy transduction or timing of ciliary beat.8

Diagram of a normal cilium illustrating the EF face and PF face in the region of the ciliary necklace. Note the location of the strands at the base of the cilium and the lower density of particles in the EF face.
Nasal biopsy specimens from three well documented patients with PCD were freeze fractured to study the ultrastructural organisation of the ciliary membrane. To our knowledge, this is the first report characterising ciliary necklace organisation in the airway epithelium of patients with PCD.
METHODS
Samples of nasal epithelium from three clinically and laboratory documented patients with PCD were obtained under direct vision by gentle curettage of the inferior nasal turbinate. The specimens were fixed immediately in 2% paraformaldehyde/2% glutaraldehyde/0.5% tannic acid or 2% paraformaldehyde/2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.2. Following fixation, the samples were rinsed for 1 hour in 0.1 M phosphate buffer containing 0.2 M sucrose and then incubated in 0.1 M phosphate buffer containing 25% glycerol. Samples were mounted between double replica specimen mounts, frozen in liquid nitrogen cooled freon, and stored in liquid nitrogen until processed.
Freeze fracture was performed in a Balzers BAF 400T unit at a stage temperature of –100°C. The fractured samples were shadowed by platinum and carbon evaporation from an angle of 30° and carbon evaporation from 90°. The replicas were cleaned in 5% sodium dichromate in 50% sulfuric acid and rinsed in several washes of distilled water. They were retrieved on copper grids and viewed in a Zeiss EM 900 transmission electron microscope at an accelerating voltage of 50 kV.
Both patients and volunteers gave informed consent before participating in the study under a protocol approved by an institutional review board on the protection of the rights of human subjects.
RESULTS
Electron microscopic examination of the freeze fracture preparations revealed that the ciliary necklaces present in the cilia of biopsy specimens from patients with PCD were comparable in organisation to those of normal healthy volunteers (fig 2). The ciliary necklaces of both patients and healthy volunteers usually consisted of five rows of particles, extending distally from the base of each cilium approximately 0.25 μm. No deviation in this arrangement was seen in the epithelial cilia preparations from patients with PCD.

{kind=link}
{kind=link}
Transmission electron micrograph of freeze fracture preparation of airway epithelium from nasal biopsy specimens showing that ciliary necklaces (arrows) from (A) healthy controls and (B) patients with PCD are comparable in appearance. The necklaces consist of 4–5 evenly spaced strands of particles and no disruption of this pattern is evident in the preparation of epithelium from the patient with PCD.
Previous studies from this laboratory have shown that disruption of the ciliary necklaces is evident in mycoplasmal infection and following in vivo exposure to sulfur dioxide, both of which contribute to ciliary dysfunction.9,10
DISCUSSION
The suggestion that ciliary necklaces function as a timing device for ciliary beat is appropriate as ciliostasis has been observed in studies where the ciliary necklaces appear to be disorganised.9,10 Generally characterised in normal subjects by rows of membrane associated particles, this is the first study of ciliary necklace organisation in patients with PCD.
An early report by Lessner et al11 studied the distribution of intramembrane particles (IMPs) on different fracture faces of cilia from patients with PCD but did not provide any specific characteristics of ciliary necklace organisation in these subjects. They found a higher density of IMPs on the EF faces than on the PF faces of cilia of patients with PCD, a pattern which contrasts with the distribution seen in normal subjects. They further concluded that the higher IMP density on the EF face was probably due to a defective microtubular system in these patients.11
In this study we have shown that ciliary necklaces present in preparations of nasal epithelial tissue from three patients with PCD appear to have normal ultrastructural organisation. Abnormal organisation of the ciliary necklace particles has been previously reported with experimental Mycoplasma pneumoniae infection9 and experimental exposure to sulfur dioxide exposure.10 In the mycoplasma study only remnants of the necklaces remained or the necklaces were completely absent in infected tissues.9 Following exposure to sulfur dioxide, remnants of the necklaces were also observed at the bases of the cilia in addition to distal displacement of the particles along the ciliary membrane.10 The ciliary necklaces of patients with PCD lacked these abnormalities, so we conclude that the ciliostasis and other irregularities of ciliary motility seen in patients with PCD are not directly associated with structural abnormalities of the ciliary necklace.
The functional decrements in ciliary activity and mucociliary clearance associated with upper respiratory infection and/or irritant exposure, together with the observed anomalies of ciliary necklace organisation, suggest a significant role for necklace proteins in maintaining normal mucociliary function. Because of this link between structure and function, it is plausible to hypothesise that necklace organisation may have a role in the pathophysiology of PCD, but our findings suggest that a direct structure/function anomaly does not occur.
This study provides the first characterisation of ciliary necklace organisation of epithelial cells from patients with PCD. Although organisation of the ciliary necklace in patients with PCD was found to be the same as that of healthy volunteers, this does not preclude membrane or membrane-axoneme dysfunction in the pathophysiology of PCD.
Acknowledgments
This research was supported in part by grants HL56395, HL34322, and HL60280 from the National Heart, Lung, and Blood Institute. The US Environmental Protection Agency through its office of Research and Development partially funded and collaborated in the research described here under Cooperative Agreement #CR824915 to Philip A Bromberg. This work has not been subjected to Agency review and therefore does not necessarily reflect the views of the Agency, and no official endorsements should be inferred.