Article Text

Abstract
Background: Magnolol, a compound isolated from the cortex of Magnolia officinalis, has been found to possess anti-allergic and anti-asthmatic activity.
Methods: The effect of magnolol on ionic currents was studied in cultured smooth muscle cells of human trachea with the aid of the patch clamp technique.
Results: In whole cell current recordings magnolol reversibly increased the amplitude of K+ outward currents. The increase in outward current caused by magnolol was sensitive to inhibition by iberiotoxin (200 nM) or paxilline (1 μM) but not by glibenclamide (10 μM). In inside out patches, magnolol added to the bath did not modify single channel conductance but effectively enhanced the activity of large conductance Ca2+ activated K+ (BKCa) channels. Magnolol increased the probability of these channel openings in a concentration dependent manner with an EC50 value of 1.5 μM. The magnolol stimulated increase in the probability of channels opening was independent of internal Ca2+. The application of magnolol also shifted the activation curve of BKCa channels to less positive membrane potentials. The change in the kinetic behaviour of BKCa channels caused by magnolol in these cells is the result of an increase in dissociation and gating constants.
Conclusions: These results provide evidence that, in addition to the presence of antioxidative activity, magnolol is potent in stimulating BKCa channel activity in tracheal smooth muscle cells. The direct stimulation of these BKCa channels by magnolol may contribute to the underlying mechanism by which it acts as an anti-asthmatic compound.
- magnolol
- Ca2+ activated K+ current
- large conductance Ca2+ activated K+ channel
- human tracheal smooth muscle cells
Statistics from Altmetric.com
- magnolol
- Ca2+ activated K+ current
- large conductance Ca2+ activated K+ channel
- human tracheal smooth muscle cells
Magnolol (5,5′-di-2-propenyl-1,1′-biphenyl-2,2′-diol), which was originally extracted from the Chinese medicinal herb Magnolia officinalis, is used as an anti-allergic and anti-asthmatic compound with a wide spectrum of pharmacological activities. Saiboku-to, a herbal medicine for bronchial asthma, consists of a number of different herbs, one of which is Magnolia officinalis.1 It has been reported that this compound inhibits the respiratory burst in formylmethionyl-leucyl-phenylalanine activated neutrophils,2 suppresses the overproduction of nitric oxide and tumour necrosis factor (TNF)-α in lipopolysaccharide activated macrophages,3 protects transplanted organs from lipid peroxidation,4 and prevents cortical neurons from chemical hypoxia in rats.5 Several studies have shown that magnolol exerts an antioxidative effect on neutrophil adhesion in vitro and in septic rats.6–8
Furthermore, this compound has previously been reported to decrease the release of serotonin, dopamine, and norepinephrine in the rat hypothalamus,9,10 to suppress catecholamine release from bovine adrenal chromaffin cells,11 to reduce the increased production of prostaglandin E2 caused by chemical hypoxia in neurons12 and the release of leukotrienes from A23187 stimulated polymorphonuclear leucocytes,13 and to suppress the activity of leukotriene synthase in rat basophilic leukaemic cells.14 Magnolol was also found to interact with γ-aminobutyric acid receptors in rat brain.15 However, to our knowledge, none of the studies have thus far demonstrated the underlying mechanism of action of magnolol on ionic currents in tracheal smooth muscle cells, even though there previous reports have shown that this compound can alleviate the symptoms of asthmatic attacks.1,13,16
Large conductance Ca2+ activated K+ (BKCa) channels play an important role in controlling the excitability of nerve, muscle, and other cells by stabilising the cell membrane at negative potentials.17 These channels, the gating of which can be controlled by intracellular Ca2+ levels and/or membrane depolarisation, are known to be present in virtually every type of smooth muscle cell.17,18 The activity of these channels is important in relaxation of airway smooth muscle.19
Cultured smooth muscle cells of human trachea were used in the present study. This cell line is known to possess many biochemical characteristics of tracheal smooth muscle cells.20 The objective was to examine the effect of magnolol on K+ outward currents in these tracheal smooth myocytes and to determine whether magnolol affects the activity and kinetic properties of BKCa channels expressed in these cells. The results clearly indicate that magnolol can effectively enhance the activity of BKCa channels and these effects may lead to a decrease in the excitability of airway myocytes.
MATERIALS AND METHODS
Cell culture
Smooth muscle cell line (HTSMC), originally derived from human trachea, was obtained from Cell Applications Inc (San Diego, CA, USA).20 Cells were maintained in smooth muscle cell growth medium (Cell Applications Inc) and equilibrated in a humidified atmosphere of 5% CO2/95% air at 37°C. The cells were subcultured weekly after detachment by using culture medium containing 1% trypsin. The experiments were performed after the cells reached confluence (usually 5–7 days).
Electrophysiological measurements
Immediately before each experiment cells were dissociated and an aliquot of the cell suspension was transferred to a recording chamber mounted on the stage of an inverted microscope (Diaphot 200; Nikon, Tokyo, Japan). Cells were bathed at room temperature (20–25°C) in normal Tyrode's solution containing 1.8 mM CaCl2. Patch pipettes were prepared from Kimax capillary tubes (Vineland, NJ, USA) using a vertical two step electrode puller (PP-83; Narishige, Tokyo, Japan) and the tips were fire polished with a microforge (MF-83; Narishige). The pipette had a resistance of 3–5 MΩ when immersed in normal Tyrode's solution. A programmable stimulator (SMP-311; Biologic, Claix, France) was used for the digital generation of rectangular or linear voltage pulses. Ionic currents were recorded in either whole cell or inside out configuration of the patch clamp technique by using a patch clamp amplifier (RK-400; Biologic).21,22 All potentials were corrected for liquid junction potential, a value that develops at the tip of the pipette when the composition of the pipette solution was different from that of the bath.
Data recording and analysis
The signals consisting of voltage and current tracings were monitored with a digital storage oscilloscope (model 1602; Gould, Valley View, OH, USA) and an LCD projector (AV600; Delta, Taipei, Taiwan). The data were simultaneously recorded using a digital audiotape recorder (model ZA5ES; Sony, Tokyo, Japan). Currents were low pass filtered at 1 or 3 kHz. A Digidata 1320A interface (Axon Instruments, Foster City, CA, USA) was used for the analogue-to-digital/digital-to-analogue conversion. This interface device was connected to a Pentium III grade laptop computer (Slimnote VX3; Lemel, Taipei, Taiwan) through a USB port and was then controlled with the aid of pCLAMP 8.02 software package (Axon Instruments). Ionic currents were analysed off line with the aid of the Clampfit subroutine in the pCLAMP software or the Origin 6.0 software (Microcal Software Inc, Northampton, MA, USA).
To calculate the percentage increase in BKCa channel activity resulting from magnolol, the potential was held at the level of +30 mV and the bath solution contained 0.1 μM Ca2+. The probability of channel openings measured after application of magnolol (30 μM) was taken as 100%. The concentration dependent relation of magnolol on channel activity was fitted to a Hill function: where [C] is the concentration of magnolol, EC50 and n are the half maximal concentration of magnolol required to increase the probability of channel openings and the Hill coefficient, respectively, and Emax is the magnolol induced maximal increase in channel activity. The EC50 value of magnolol and its 95% confidence limits were calculated with the Origin program (Microcal Software Inc) using non-linear regression methods to obtain the best fit dose-response curve.
where [C] is the concentration of magnolol, EC50 and n are the half maximal concentration of magnolol required to increase the probability of channel openings and the Hill coefficient, respectively, and Emax is the magnolol induced maximal increase in channel activity. The EC50 value of magnolol and its 95% confidence limits were calculated with the Origin program (Microcal Software Inc) using non-linear regression methods to obtain the best fit dose-response curve.
Single channel currents of the BKCa channel were analysed with Fetchan and Pstat subroutines in the pCLAMP software (Axon Instruments). Multigaussian adjustments of the amplitude distributions between channels were used to determine single channel currents. Functional independence between channels was then verified by comparing the observed stationary probabilities with the values calculated according to the binomial law.
To determine the effect of magnolol on the activation curve of BKCa channels, the ramp pulses from 0 to +80 mV with a duration of 1 second were delivered from the Clampex subroutine of pCLAMP software. The activation curve was calculated by averaging current traces in response to 20 voltage ramps and dividing each point of the mean current by the unitary amplitude of each potential after the leakage component was corrected.22,23 The number of active channels in the patch (N) was counted at the end of each experiment when the solution with 100 μM Ca2+ was perfused This number was used to normalise the probability of channel openings at each potential. The activation curves obtained before and after the addition of magnolol were fitted with Boltzmann function of the form: where n is the maximal relative N • P0, b is the slope factor of the voltage dependent activation (that is, the change in the potential required to result in an e-fold increase in the activation), and a is the voltage at which there is half maximal activation.
where n is the maximal relative N • P0, b is the slope factor of the voltage dependent activation (that is, the change in the potential required to result in an e-fold increase in the activation), and a is the voltage at which there is half maximal activation.
Open or closed lifetime distributions were fitted with logarithmically scaled bin width using the method of McManus et al.22,24 To estimate all transition rates between states, single channel data were idealised and converted to an ASCII format—that is, a dwell time file format—using the SKM program of the QuB software suite that was downloaded from the web site (http://www.qub.buffalo.edu). This type of format was readily read by the maximum interval likelihood (MIL) program in the QuB suite.25,26 The data were then used to determine single channel kinetic parameters by means of a maximum likelihood algorithm.25,26 The highest log likelihood for the observed activity of BKCa channels in this study was obtained with the gating scheme: C1 ↔ C2 ↔ O. The single channel data were modelled on the basis of this simplified scheme. Simulated single channel data were also obtained using the determined transition rates and SIMU program. K and L values represent the equilibrium dissociation constant that equals the transition rate constant for C1 ← C2 divided by that for C1 → C2, and the equilibrium gating constant that equals opening rate constant divided by closing rate constant, respectively. This type of kinetic analysis was based on the assumption that the channel can be described by a finite state Markovian model and that the channels in the patch are mutually independent.
All values are reported as mean (SE). The paired or unpaired Student's t test and one way ANOVA with the least significance difference method for multiple comparisons were used for the statistical evaluation of differences in the mean values. Differences between values were considered significant at a value of p<0.05.
Drugs and solutions
Magnolol (5,5′-di-2-propenyl-1,1′-biphenyl-2,-2′-dol), an amphipathic molecule, is composed of two phenol rings with one polar hydroxyl and one non-polar hydrocarbon domain on each phenol ring (fig 1).27 Because of its unique property, magnolol can freely permeate through cell membranes. Niflumic acid was purchased from Sigma Chemical (St Louis, MO, USA); 18α-glycyrrhetinic acid and baicalein were obtained from Aldrich Chemical (Milwaukee, WI, USA); iberiotoxin, glibenclamide, apamin, and sodium nitroprusside were obtained from Research Biochemicals (Natick, MA, USA); paxilline was purchased from Alomone (Jerusalem, Israel); and magnolol was a kind gift of Professor Chien-Chich Chen, National Institute of Chinese Medicine, Taipei City, Taiwan. All other chemicals were commercially available and of reagent grade.

Chemical structure of magnolol (5,5`-di-2-propenyl-1, 1`-biphenyl-2,2`-diol).
The composition of normal Tyrode's solution was 136.5 mM NaCl, 5.4 mM KCl, 1.8 mM CaCl2, 0.53 mM MgCl2, 5.5 mM glucose, and 5.5 mM HEPES NaOH buffer, pH 7.4. To record K+ currents the patch pipette was filled with a solution consisting of 140 mM KCl, 1 mM MgCl2, 3 mM Na2ATP, 0.1 mM Na2GTP, 0.1 mM EGTA, and 5 mM HEPES KOH buffer, pH 7.2. In the single channel recording, high K+ bathing solution contained 145 mM KCl, 0.53 mM MgCl2, and 5 mM HEPES KOH, pH 7.4, and the pipette solution contained 145 mM KCl, 2 mM MgCl2, and 5 mM HEPES KOH, pH 7.2. The value of free Ca2+ concentration was calculated assuming a dissociation constant for EGTA and Ca2+ (at pH 7.2) of 0.1 μM.28
RESULTS
Effect of magnolol on whole cell K+ outward current (IK) in cultured smooth muscle cells of human trachea
In the patch clamp technique the whole cell configuration was used to investigate the effect of magnolol on macroscopic ionic currents in these cells. The cells were bathed in normal Tyrode's solution containing 1.8 mM CaCl2 and the pipette solution contained a low concentration (0.1 mM) of EGTA and 3 mM ATP. As shown in fig 2, when the cell was held at –40 mV and various potentials ranging from –50 to +50 mV with 20 mV increments were applied, a family of K+ outward currents were elicited and the current/voltage (I/V) relationship for these currents had an outward rectification. Within 1 minute of exposing cells to magnolol (3 μM) the amplitude of the outward currents was greatly increased throughout the entire range of voltage clamp steps. For example, when the depolarising pulses from –40 to +50 mV were applied, magnolol (3 μM) significantly increased current amplitude from 1105 (150) pA to 2794 (280) pA (n=8). This stimulatory effect was readily reversed when magnolol was washed out. Figure 2B shows the mean I/V relationships for these currents in the absence and presence of magnolol (3 μM).

Stimulatory effect of magnolol on K+ outward current (IK) in cultured smooth muscle cells of human trachea (HTSMC). (A) Superimposed current traces in the absence (left) and presence (right) of 3 μM magnolol. The upper part in A indicates the voltage protocol. The cells, bathed in normal Tyrode's solution containing 1.8 mM CaCl2, were held at –40 mV and voltage pulses from –50 to +50 mV in 20 mV increments were applied at 0.05 Hz. Arrows indicate zero current level. (B) Mean (SE) relationship between current and voltage of outward currents measured at the end of voltage pulses in control samples, after application of 3 μM magnolol, and after washout of magnolol (n=7–12 for each point).
To examine the nature of IK stimulated by magnolol, the values of reversal potential obtained during cell exposures to magnolol (3 μM) were measured at different concentrations of extracellular K+. The data from each cell were then pooled and plotted as a function of extracellular K+. The line of best fit through the mean data revealed a slope of 58 mV per tenfold increase in extracellular K+. The outward current measured after application of magnolol therefore indicates that the K+ selective channel exhibits Nernstian behaviour.
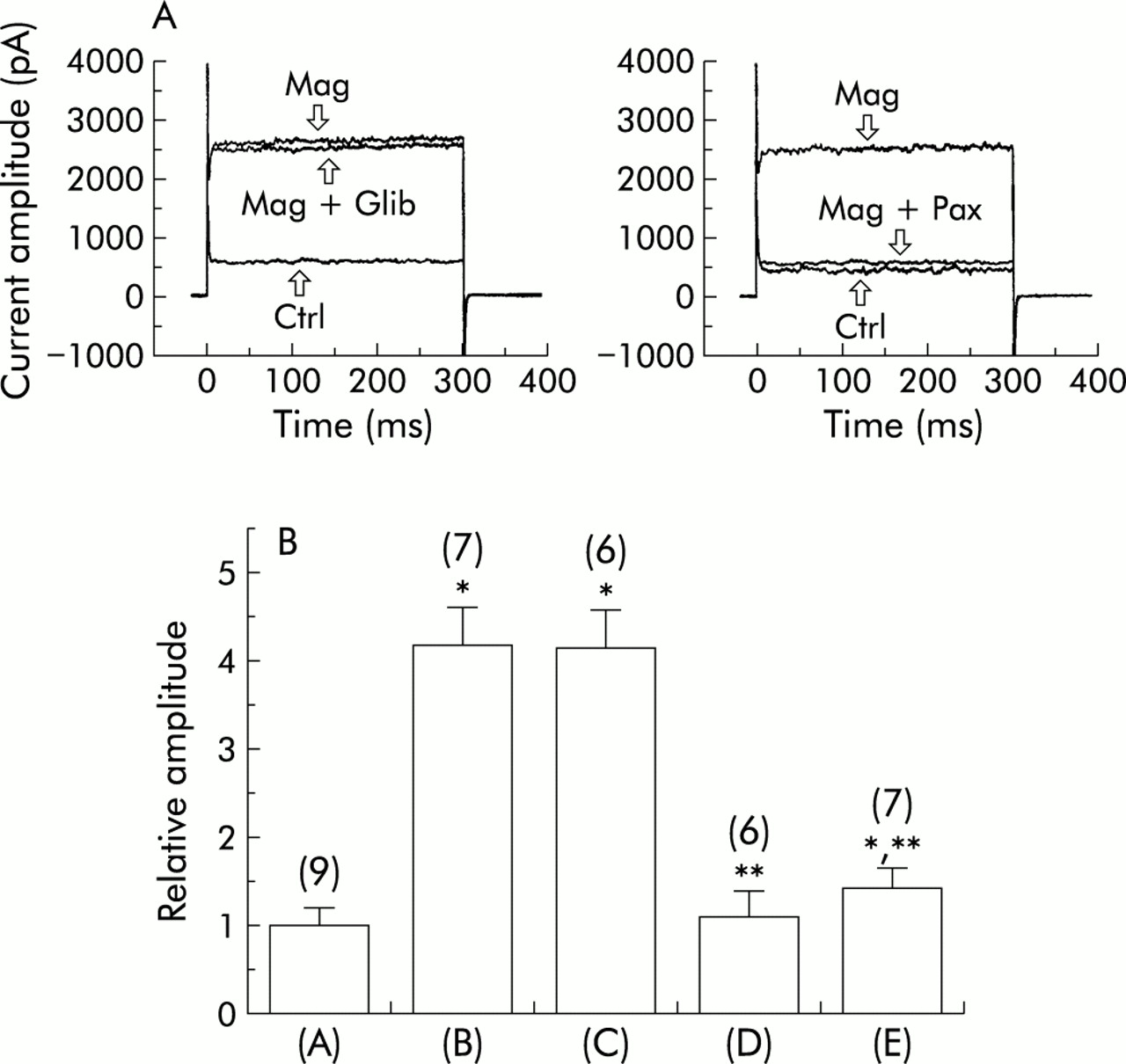
The effects of various K+ channel blockers including glibenclamide, iberiotoxin, paxilline, and apamin on magnolol stimulated IK in these cells were also examined and compared. As shown in fig 3, glibenclamide (10 μM) did not affect the magnolol mediated increase in the amplitude of IK significantly. However, both iberiotoxin (200 nM) and paxilline (1 μM) was able to suppress the increase in IK caused by magnolol (10 μM). Paxilline, an indole alkaloid isolated from Penicillium paxilli, was reported to be a potent inhibitor of BKCa channels.29 These results indicate that the observed increase in outward current by magnolol is due to its stimulation of IK which is sensitive to inhibition by iberiotoxin or paxilline.

Effect of various K+ channel blockers on magnolol stimulated IK in cultured smooth muscle cells of human trachea. (A) Original current traces showing the effect of glibenclamide (Glib; 10 μM) or paxilline (Pax; 1 μM) on the increased amplitude of IK caused by magnolol (Mag; 3 μM). Each cell was depolarised from –40 to +30 mV with a duration of 300 ms. (B) Effect of various compounds on magnolol induced increase in IK in these cells. The magnitude of IK in the control was considered to be 1.0 and the relative amplitude of IK after application of each agent was plotted. The numbers in parentheses denote the number of cells examined. Values are mean (SE). A = control; B = magnolol (3 μM); C = magnolol (3 μM) + glibenclamide (10 μM); D = magnolol (3 μM) + paxilline (1 μM); E = magnolol (3 μM) + iberiotoxin (200 nM). *Significantly different from controls. **Significantly different from magnolol alone.
Stimulatory effect of magnolol on BKCa channel activity in cultured smooth muscle cells of human trachea
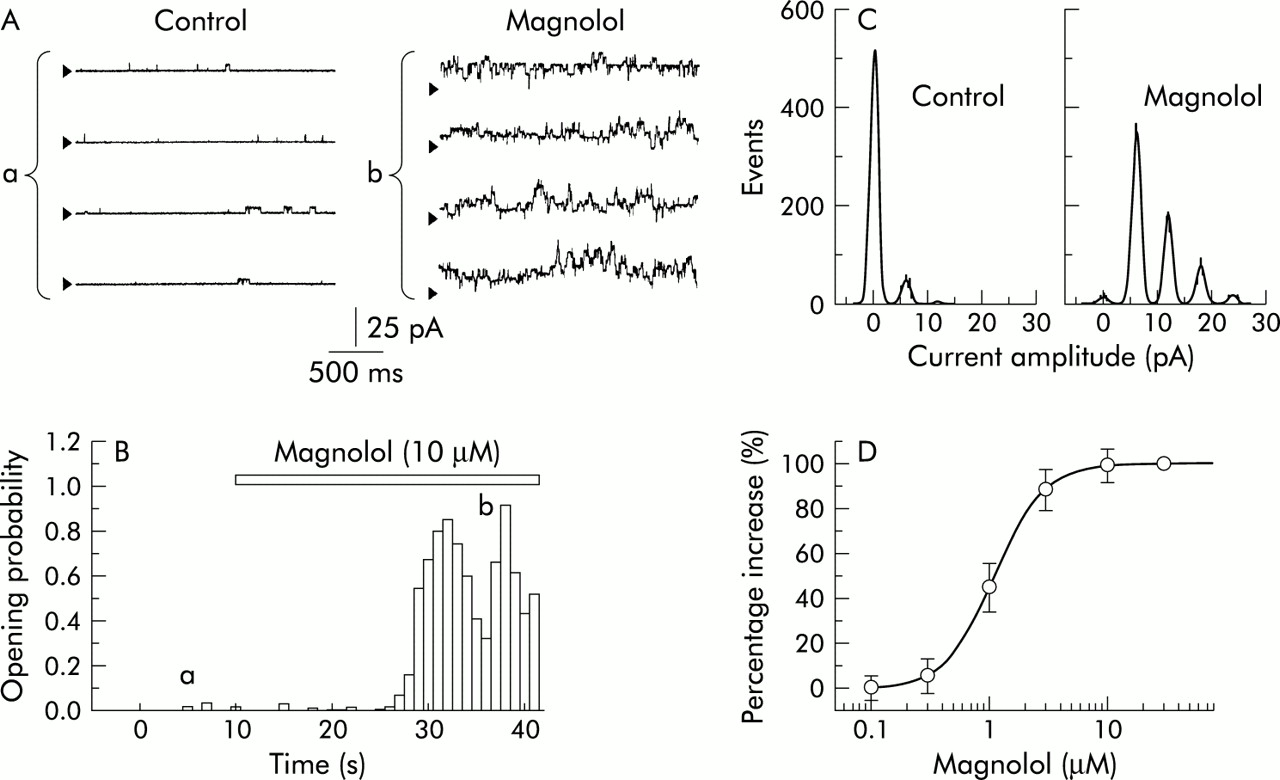
Since macroscopic IK stimulated by the presence of magnolol appears to be a large noisy voltage dependent, Ca2+ sensitive K+ current, the next series of experiments were designed to investigate whether magnolol interacts with the BKCa channel to stimulate Ca2+ activated K+ currents. Unless otherwise specified, all experiments of single channel recordings performed in this study were conducted with a symmetrical K+ concentration (145 mM) and an inside out configuration, and the bath medium contained 0.1 μM Ca2+. To provide 0.1 μM free Ca2+ in the bath solution, 0.5 mM CaCl2 and 1 mM EGTA were added.28 As shown in fig 4A, when the potential was held at +30 mV, application of magnolol (10 μM) to the cytosolic surface of an inside out patch greatly enhanced single channel activity which occurred in rapid open-close transitions. When membrane patches were exposed to magnolol (10 μM) the opening probability of BKCa channels was significantly increased to 0.622 (0.022) from a control of 0.013 (0.005) (n=8). Channel activity was reduced to 0.018 (0.007) (n=6) after washout of magnolol. However, there was no significant difference in the amplitude of single channel current between the absence and presence of magnolol (10 μM; fig 4C).

Stimulatory effect of magnolol on BKCa channels in cultured smooth muscle cells of human trachea. The experiments were conducted in a symmetrical K+ concentration (145 mM) and bath solution contained 0.1 μM Ca2+. In an inside out configuration, channel activity was measured at a holding potential of +30 mV. (A) Original current traces recorded before and after application of 10 μM magnolol to intracellular face of channels in a detached patch. The original current traces labelled “a” and “b” in (A) correspond to those labelled “a” and “b” in (B). Arrowheads shown in each current trace indicate zero current level. Channel openings are shown as an upward deflection. (B) Time course of change in the probability (1 second bin width) of channel openings before and during bath application of magnolol (10 μM). Horizontal bar indicates the addition of magnolol (10 μM) into the bath. (C) Amplitude histograms measured in the absence (left) and presence (right) of magnolol (10 μM). All data points shown in the amplitude histograms were fitted by one or more Gaussian distributions using the method of maximum likelihood. (D) Concentration-response relationship for magnolol stimulated increase in channel activity. The channel activity obtained in the presence of 30 μM magnolol was considered to be 100%. The smooth curve was fitted with the Hill equation. The EC50 value and maximal opening probability of BKCa channels were 1.5 μM and 99%, respectively. The Hill coefficient was 2.2. Each point represents the mean (SE) value (n=6–9).
The relationship between the concentration of magnolol and the opening probability of BKCa channels was further examined using a holding potential of +30 mV. As shown in fig 4D, magnolol (0.1–30 μM) effectively increased the probability of channel openings in a concentration-dependent manner. The EC50 value for magnolol stimulated channel activity was 1.5 μM. In addition, the Hill coefficient was found to be 2.2, suggesting that there was positive cooperation for the stimulation of BKCa channels.
Lack of effect of magnolol on single channel conductance of BKCa channels
The effect of magnolol on the activity of BKCa channels at various membrane potentials was also examined (fig 5). The plots of single channel amplitude as a function of holding potential were constructed. Figure 5B illustrates the I/V relationship of BKCa channels in the absence and presence of magnolol (10 μM). The single channel conductance of BKCa channels calculated from a linear I/V relationship in control solutions (in the absence of magnolol) was 196 (8) pS (n=12) with a reversal potential of 0 (3) mV (n=12). This value for single channel conductance of BKCa channels was similar to that reported previously in tracheal smooth muscle cells,30 but was not significantly different from that measured after application of magnolol (197 (8) pS; n=10). Thus, it is clear that the presence of magnolol effectively enhances the activity of BKCa channels but produces no significant change in single channel conductance in these cells.
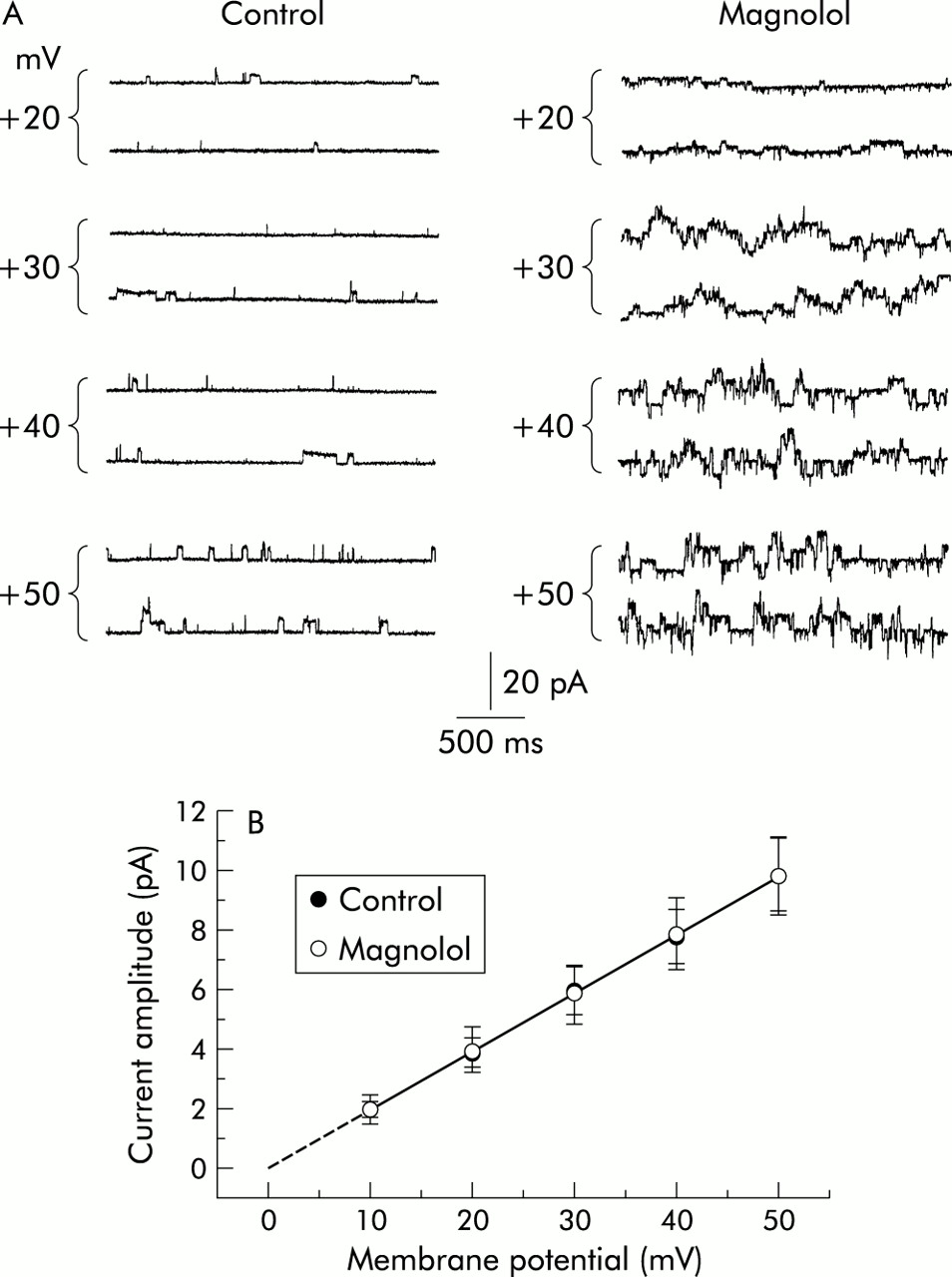
Effect of magnolol on the I/V relationship of BKCa channels in cultured smooth muscle cells of human trachea. (A) Examples of BKCa channels in the absence (left) and presence (right) of magnolol (10 μM) measured from an inside out patch at various membrane potentials. Magnolol was applied to the bath medium. The number shown at the beginning of each current trace indicates the voltage applied to the patch pipette. Channel opening gives an upward deflection in current. (B) I/V relationship of BKCa channels in the absence and presence of magnolol (10 μM). Values are mean (SE) (n=4–8).
Effect of magnolol on voltage dependent activation of BKCa channels from cultured smooth muscle cells of human trachea
Figure 6A shows the activation curves of BKCa channels in the absence and presence of 1 and 3 μM magnolol. In these experiments the activation curve of BKCa channels was obtained with the aid of voltage ramp protocols.22,23 The ramp pulses were applied from 0 to +80 mV with a duration of 1 second. The plots of the probability of channel openings as a function of membrane potential were constructed and fitted with the Boltzmann function as described in the Methods section. In the control n=0.37 (0.04), a=49.1 (1.5) mV, and b=9.7 (0.6) mV (n=6); in the presence of 1 and 3 μM magnolol n=0.55 (0.05) and 1.02 (0.08), a=43.5 (1.1) and 35.2 (1.6) mV, and b=9.6 (0.7) and 9.5 (0.7) mV, respectively (n=6). Thus, application of magnolol not only produced an increase in the maximal opening probability of BKCa channels, but also significantly shifted the activation curve to the less positive potential. In contrast, magnolol had no effect on the slope (b value) of the activation curve. These data clearly indicate that the magnolol induced change in the activity of BKCa channels in these tracheal smooth muscle cells is dependent on the level of membrane potential.

Voltage and Ca2+ dependence of magnolol induced increase in the probability of channel openings in cultured smooth muscle cells of human trachea. (A) Effect of magnolol on the activation curve of BKCa channels. The experiments were conducted with a symmetrical K+ concentration (145 mM). In an inside out configuration, holding potential was set +40 mV and the bath medium contained 0.1 μM Ca2+. The activation curves of the BKCa channels were obtained when ramp pulses from 0 to +80 mV with a duration of 1 second were applied at a rate of 0.05 Hz. The smooth lines showed Boltzmann fits of the data yielding a value for half maximal activation (a) of 49.1 mV for the control and 43.5 and 35.2 mV when the membrane patch was exposed to 1 and 3 μM magnolol, respectively. (B) Stimulation of BKCa channels by magnolol at various concentrations of internal Ca2+ in the bath before (control) and during exposure to 3 μM magnolol. *Significantly different from controls.
Effect of internal Ca2+ concentration on magnolol stimulated activity of BKCa channels in cultured smooth muscle cells of human trachea
To determine whether the magnolol induced increase in channel activity involves the level of internal Ca2+ concentration, another series of experiments was conducted with various concentrations of Ca2+ in the bath and the probability of channel openings before and after application of magnolol was examined. As shown in fig 6B, at a given concentration of magnolol (3 μM) the magnitude of the magnolol induced increase in channel activity was increased as the internal Ca2+ concentration was increased. However, the observed effect of magnolol on channel activity was unaffected by a change in the internal Ca2+ concentration. At a holding potential of +30 mV the presence of magnolol (3 μM) at internal Ca2+ concentrations of 0.01, 0.1, and 1 μM enhanced channel activity to a similar extent (about a fourfold increase).
Effect of magnolol on kinetic behaviour of BKCa channels
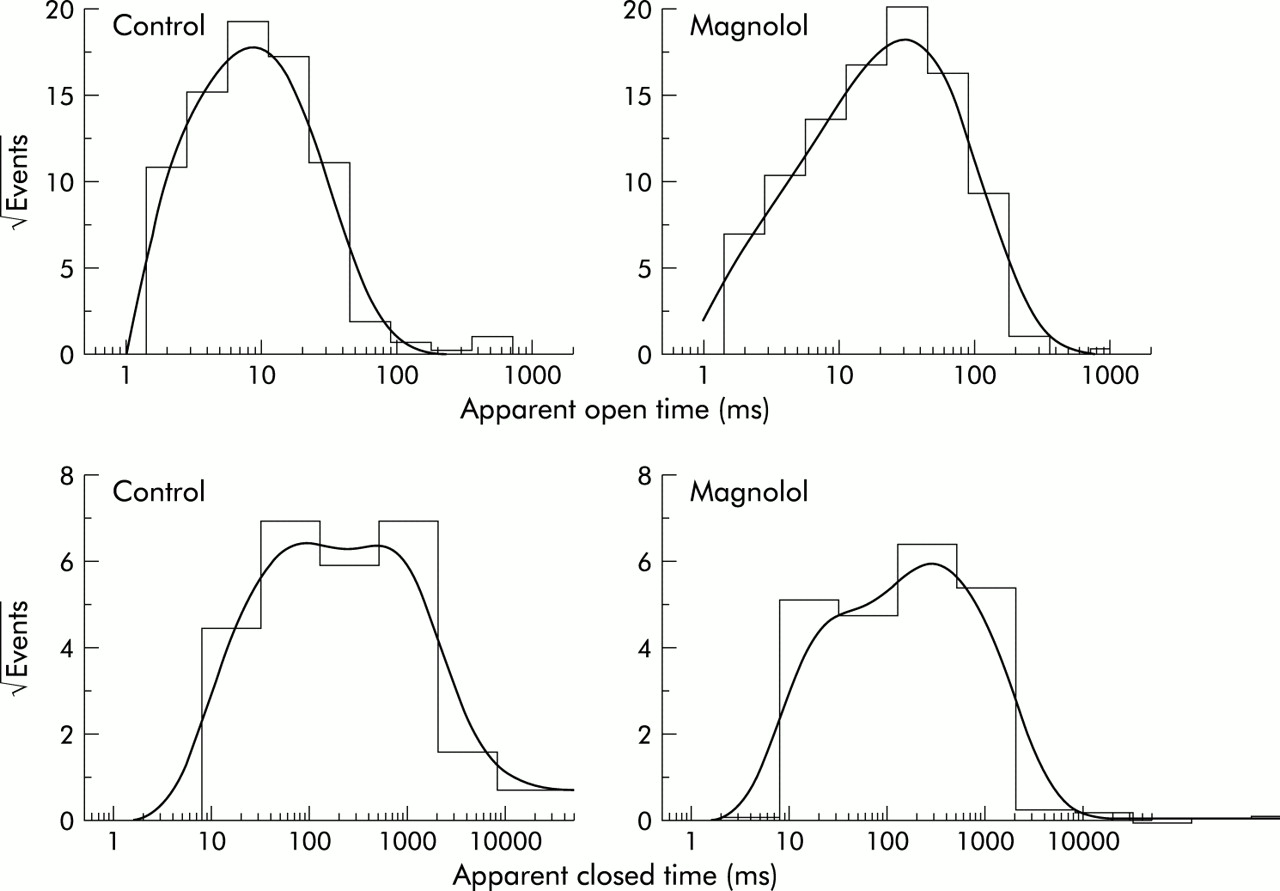
Because magnolol produced no change in single channel conductance, the effect of magnolol on the gating of these channels was also analysed. As shown in fig 7, in a detached patch of control cells—that is, in the absence of magnolol—open and closed time histograms at the level of +30 mV can be fitted by a one or two exponential curve. In addition, the time constant for the open time histogram was 8.9 (1.2) ms, whereas those for the fast and slow components of the closed time histogram were 43.4 (4.5) and 721.0 (13.5) ms, respectively (n=7). Magnolol (3 μM) added to the bath increased the time constant of the open state to 32.2 (4.3) ms (n=6) and decreased the mean closed times to 16.6 (3.2) and 419.6 (10.8) ms (n = 6). Thus, magnolol produced an increase in channel open time and a decrease in channel closed time. The presence of magnolol may cause the channel to spend more time in the open state with a longer open time. Changes in mean open and closed times caused by magnolol might explain its stimulatory effect on the activity of BKCa channels.

Effect of magnolol on mean open and closed times of BKCa channels in cultured smooth muscle cells of human trachea. Inside out configuration was performed in these experiments and the potential was held at +30 mV to reduce the number of channel openings. Cells were bathed in a symmetrical K+ solution (145 mM) and the bath medium contained 0.1 μM Ca2+. The open (upper) and closed time (lower) histograms after application of magnolol (3 μM) are shown on the right. Of note, the abscissa and ordinate show the logarithm of apparent open or close times (ms) and the square root of the number of events (n0.5), respectively. Control data were obtained from measuring 995 channel openings with a total record time of 3 minutes, whereas those after application of magnolol were measured from 984 channel openings with a total record time of 2 minutes.
In order to obtain more information on the channel kinetics, single channel currents in the absence and presence of magnolol were idealised and modelled using the kinetic scheme described in the Methods section. This gating scheme is composed of one open and two closed states. Transition rates between states were derived from the maximum likelihood estimation.25,26,29 Figure 8 shows examples of simulated single channel data that were generated using the transition rates of BKCa channels in the absence and presence of magnolol. The equilibrium dissociation constants (K values) obtained in the absence and presence of magnolol (10 μM) were estimated to be 0.44 (0.03) and 1.37 (0.04), respectively (n=6), a difference of about threefold. However, the equilibrium gating constants (L values) in the absence and presence of magnolol (10 μM) were 0.184 (0.003) and 3.09 (0.005), respectively (n=6), which differ by about 17-fold. These results suggest that the difference in the gating constant in the absence and presence of magnolol is greater than in the dissociation constant.

Simulated single channel currents in the absence (upper) and presence of 3 and 10 μM magnolol. The kinetic models used to analyse the observed data measured at a holding potential of +30 mV are shown in the upper part of each simulated current. Each horizontal arrow pointing to the left represents the binding of magnolol or closing of a channel, whereas each arrow to the right represents the dissociation of magnolol or opening of a channel. The units are μM/s or /s. O=open state; C1 and C2=first and second closed states.
Comparison between effect of magnolol and those of baicalein, glycyrrhetinic acid, niflumic acid and Evans blue
The effects of magnolol, baicalein, glycyrrhetinic acid, glibenclamide, and paxilline on the activity of BKCa channels in cultured smooth muscle cells of human trachea were also examined and compared. As shown in fig 9, neither baicalein (10 μM) nor glycyrrhetinic acid (10 μM) applied intracellularly had any significant effect on the probability of channel openings. These compounds are known to be other phenolic compounds contained in Saiboko-to, a herbal medicine used to treat bronchial asthma.1,13,16 However, both niflumic acid (30 μM) and Evans blue (30 μM) enhanced the channel activity significantly. These two compounds have previously been reported to stimulate BKCa channel activity in vascular endothelial cells.29,31 In addition, paxilline effectively suppressed the increased activity of BKCa channels caused by magnolol which is consistent with the finding in whole cell current recordings (fig 3). Thus, it is clear that magnolol is more potent than niflumic acid or Evans blue in stimulating the activity of BKCa channels in these cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comparison between the effect of magnolol and those of baicalein, glycyrrhetinic acid, niflumic acid, and Evans blue on the activity of BKCa channels in cultured smooth muscle cells of human trachea. Inside out configuration was performed in this series of experiments. The membrane potential held at each detached patch was +30 mV and the bath medium contained 0.1 μM Ca2+. The channel activity in the absence of each agent was considered to be 1.0 and the relative opening probability after application of each agent was plotted. The numbers in parentheses denote the number of cells examined. Values are mean (SE).*Significantly different from control group. **Significantly different from magnolol alone.
DISCUSSION
The major findings in the present study are as follows: (a) in cultured smooth muscle cells of human trachea magnolol can reversibly increase the amplitude of K+ outward current; (b) magnolol stimulates the activity of BKCa channels in a concentration and voltage dependent manner but does not modify single channel conductance of these channels; (c) the magnolol induced increase in the probability of channel openings is accompanied by an increase of the time constant in the open state and a decrease in the time constant in the closed state; and (d) magnolol can modify both dissociation and gating constants of these channels. The stimulatory effect of this compound on BKCa channel activity would be expected to affect cell excitability if similar results are present in airway myocytes in vivo.18,19,30
The single channel conductance of BKCa channels measured with the use of a symmetrical K+ concentration (145 mM) on both sides of the membrane was 196 (8) pS (n=12). This value is similar to those of typical BKCa channels reported previously in swine tracheal smooth myocytes.30 It was also found that, in these cells, the channel activity stimulated by magnolol is sensitive to inhibition by paxilline. However, the density of BKCa channels reported here in cultured smooth muscle cells of human trachea appears to be lower than that in freshly dissociated airway myocytes, since we did not clearly observe a unique type of spontaneous transient outward K+ current as shown previously.30,32 On the other hand, because there is a lower density of BKCa channels, single channel properties may be easily identified and examined in these smooth muscle cells of human trachea.
It seems likely that magnolol did not exert its effect on BKCa channel activity via an increase in the affinity of Ca2+ ions or the Ca2+ binding site in the membrane because magnolol stimulated channel activity was found to be independent of internal Ca2+. The EC50 value of magnolol required for the stimulation of BKCa channels was 1.5 μM, a value that is lower than those used to produce the antioxidative actions.5–8 BKCa channels expressed in airway smooth myocytes may therefore be a relevant “target” for the action of magnolol. Regardless of the mechanism of its actions, findings from the present study demonstrating the stimulatory effect of magnolol on BKCa channels should be noted with caution in relation to its increasing use as an antioxidant agent.4,6–8
Previous reports have shown that magnolol can reduce the level of intracellular cyclic AMP in rat neutrophils.2 It was believed that the magnolol mediated inhibition of the respiratory burst in formylmethionyl-leucyl-phenylalanine activated neutrophils appeared to involve the attenuation of tyrosine kinase phosphorylation or mitogen activated protein kinase activation.2 The inhibition of phorbol ester induced neutrophil aggregation by magnolol is found to be associated with the inhibition of the activity of protein kinase C.33 Previous studies have also shown that magnolol can stimulate Ca2+ release from internal stores and Ca2+ influx across plasma membrane in rat neutrophils.34 However, in our experimental conditions, the effect of magnolol on ionic currents was observed within only 2 minutes of application of this compound. Furthermore, in an inside out configuration magnolol added to the bath is effective in stimulating the activity of BKCa channels. It is thus possible that the magnolol stimulated increase in BKCa channel activity in these cells is direct and does not require cytosolic diffusible substances inside the cells. Magnolol mediated increases in the activity of BKCa channels observed in these airway smooth muscle cells are also unlikely to result from an increase in intracellular Ca2+ that is induced by Ca2+ release from internal stores, Ca2+ entry from the cell exterior, or both.35
It seems to be uncertain whether the presence of magnolol affects the production of nitric oxide. A previous report suggested that the antiarrhythmic effect of magnolol during acute coronary occlusion might involve an increase in the activity of nitric oxide synthase.36 On the other hand, in lipopolysaccharide activated macrophages magnolol was found to suppress the production of nitric oxide.3 However, in our study, when tracheal smooth muscle cells were preincubated with sodium nitroprusside, magnolol mediated stimulation of outward currents was not significantly affected (data not shown). The magnolol induced effect on the generation of nitric oxide does not therefore appear to be primarily responsible for the stimulation of BKCa channels observed in the present study.
In addition to magnolol, both baicalein and glycyrrhetinic acid are contained in Saiboku-to which has been found to possess antiasthmatic activity.1,13 However, in our study neither baicalein nor glycyrrhetinic acid affected the activity of BKCa channels. Magnolol thus appears to be an important ingredient present in Saiboko-to that can effectively stimulate BKCa channels. Moreover, it will be of interest to determine whether magnolol contained in Saiboko-to1 decreases the release of leukotrienes through a mechanism of stimulatory action on BKCa channel activity.13
The increase in IK caused by magnolol is apparently not a result of an increase in single channel amplitude of BKCa channels because no significant difference was found in single channel conductance in the absence or presence of magnolol. However, this compound increased the activity of BKCa channels in tracheal smooth muscle cells in a voltage dependent fashion and possibly by acting at a site that is accessible from the intracellular side of the channel. We also found that magnolol produced an increase in mean open time and a decrease in mean closed time. These findings can account for the increased probability of channel openings. The concentration-response curve of channel activation by magnolol had a Hill coefficient of 2.2, suggesting that more than one magnolol molecule may bind to the channel, although the stoichiometry of magnolol binding to the channel is currently unknown. More interestingly, on the basis of the simple gating scheme described in the Methods section, the mechanism of action of magnolol on BKCa channel activity was also deduced. The results showed that the stimulatory actions of magnolol appeared to lie not only in the binding step, but also in the gating of channels because both the equilibrium dissociation constant and the gating constant were significantly altered after application of magnolol. However, we cannot rule out the possibility that the binding of magnolol shifts the channel to longer open state(s). In addition, how these binding events are functionally coupled to the opening of channel pores has yet to be elucidated.
In summary, the data reported here provide evidence that, in addition to stimulating steroidogenesis in adrenal cells,37 magnolol is potent in stimulating the activity of BKCa channels in cultured smooth muscle cells of human trachea. The experiments did not determine whether the modulatory effect on the BKCa channel is the primary mechanism by which magnolol reduces asthmatic symptoms or whether it is secondary to effects mediated through other cell types. However, the present finding will be of great help in the study of the underlying mechanisms through which magnolol interacts with BKCa channels expressed in smooth muscle cells. The potent effect of magnolol on the stimulation of BKCa channels seems to make it a promising compound to induce relaxation of airway smooth muscle.
Acknowledgments
The authors wish to thank Yen-Hua Hung for technical assistance. The present work was aided by grants from National Science Council (NSC-89–2320-B-075B-016), Kaohsiung-Veterans General Hospital (VGHKS90-06 and VGHKS90-73), and VTY Joint Research Program, Tsou's Foundation (VTY87-S3-17), Taiwan, ROC.