Article Text

Abstract
BACKGROUND Dendritic cells (DCs) in the mucosa of the respiratory tract might be involved in the early development of pulmonary allergy or tolerance. To date, little is known about when the first DCs occur in human airways.
METHODS Specimens of the distal trachea from patients who had died from sudden death in the first year of life (n=29) and in older age groups (n=59) as well as from those who had died from respiratory tract infections in the first year of life (n=8) were examined by immunohistochemistry. Transmission electron microscopy was performed in additional samples from two adults.
RESULTS In the sudden death subgroup DCs were absent in 76% of those who died in the first year of life but were present in 53 of the 59 older cases. All infants who had died of respiratory infectious diseases had DCs in the tracheal mucosa.
CONCLUSIONS Mature DCs are not constitutively present in the human tracheobronchial mucosa in the first year of life, but their occurrence seems to be triggered by infectious stimuli. These data support the hypothesis that DCs play a crucial role in immunoregulation in early childhood.
- dendritic cells
- human respiratory mucosa
- infectious diseases
Statistics from Altmetric.com
Dendritic cells (DCs) are involved in antigen uptake, processing, and presentation of antigenic fragments to T cells and play a major role in mucosal defence including that of the respiratory tract.1-5 Many data on DCs have been acquired from animal models and from human blood DCs or precursors of DCs. However, there are few data on DCs in the mucosa of the human respiratory tract in health and disease.6-8 To date, only adult patients have been investigated, the tissues being obtained from thoracotomy samples in various pulmonary diseases—mainly pulmonary carcinomas—or from mucosal biopsy specimens. Limited access to so called “normal” human tissues is not surprising because necroscopic examinations in cases of acute death are rare.
Dendritic cell precursors from the bone marrow and blood are well described.9 ,10 Different stimuli result in a dramatic increase in DCs within the respiratory mucosa in animal experiments, even before that of granulocytes.6 It is not known whether local proliferation or immigration from the blood is the main cause of this phenomenon. DCs have also been discussed in connection with the pathophysiology of asthma and the influence of cortisol on the presence of DCs has been investigated.11 Epidemiological observations suggest an important role in the development of allergy or tolerance during early childhood.12-15 However, as was pointed out in a recent review,16 ,17 there are no data on the presence and development of mature mucosal DCs during this period. The clinical relevance of the development of neonatal immunity to allergic diseases in childhood and in adults is currently under discussion.18 ,19 Because DCs are necessary for the initiation of T cell responses and are able to skew the T cell response to Th1 or Th2, it is thought that they might be involved in early immunoregulation.20-23
The afferent limb of mucosal immune responses in early infancy has not been studied in humans because of difficulties with tissue sampling. The first step was therefore to examine necroscopic material, despite problems with preservation and availability of immunohistochemical markers. This study focused on the presence of DCs in the tracheal mucosa in the first year of life. Infants who had died from sudden death (thus excluding the influence of diseases) were compared with a small group of infants who had died of respiratory tract infections. In addition, children of other ages and adults were included in the study.
Methods
This study was performed retrospectively on tracheal tissue from 96 patients, 88 of whom had died a sudden death, either from traumatic causes (n=65) or sudden infant death (SID) according to the definition of Beckwith24 (n=23). Their ages varied between 6 days and 82 years; 29 were younger than 1 year (23 SID, six traumatic causes of death). Another group comprised eight infants aged 37–294 days who had died of respiratory tract infections as confirmed by histopathological and microbiological examinations.
The distal part of the trachea (at least 1 cm in length) was excised during necropsy and fixed in 4% buffered formaldehyde. The tissue was dehydrated and embedded in paraffin. Immunohistochemical examination was performed using monoclonal antibodies against human HLA-DR and CD1a. Paraffin sections (5 μm thick) were mounted onl-polylysine coated slides and dried overnight at 37°C. The sections were dewaxed and incubated with an anti HLA-DR antibody (mouse anti-human HLA-DR, clone CR3/43, IgG1, kappa, 1:100, Dako, Hamburg, Germany) and with two antibodies against the human CD1a molecule (mouse anti-human CD1a, clone O10, IgG1, 1:50, Immunotech, Hamburg, Germany; clone LN3, IgG1, BioGenex, San Ramon, USA) for 30 minutes at room temperature. After washing with TBS-Tween (0.05 % Tween 20; Serva, Heidelberg, Germany) the sections were incubated with a polyclonal rabbit anti-mouse immunoglobulin (Dako) functioning as a bridging antibody. The APAAP complex (Dako) was then applied for 30 minutes. To increase the staining intensity the incubation with the bridging antibody and the addition of the APAAP complex were repeated once. Fast blue (Sigma; St Louis, MO, USA) served as a substrate for alkaline phosphatase. Counterstaining was not performed. Paraffin sections of human spleens served as external positive controls while paratracheal lymph nodes served as internal controls. Omitting the primary antibody or using isotype matched irrelevant antibodies produced the expected negative results.
The stained sections were examined microscopically by two independent investigators who were blinded to the source of the material. Since macrophages, B cells, and even epithelial cells can also express MHC class II molecules, the DCs were identified by a combination of HLA-DR staining, their dendritic morphology, and their typical localisation under the basement membrane of the epithelium. Either there were variable numbers of DCs within the tracheal mucosa or the DCs were completely absent.
Transmission electron microscopy was performed on tracheal samples from two additional adults (an 81 year old woman and a 40 year old man) following acute death (for details see Paulsen and Tillmann25). In brief, the samples were excised within 6 hours of death and fixed in 3.5% glutaraldehyde (in 0.1 M Sörensen phosphate buffer solution at pH 7.4) at 4°C for 1 week. After dehydration in graded concentrations of ethanol they were embedded in araldite. Ultrathin sections were prepared and contrasted with uranyl acetate and lead citrate. Examination was carried out with a Zeiss TEM 902 electron microscope.
Results
Although the tissue was obtained from necropsy samples, its morphology was adequate and in most cases the epithelium was well preserved (fig 1A). DCs were identified using HLA-DR staining and the specific dendritic morphology (fig 1A and B) and could be followed through the depth of the section. Using the anti-CD1a antibody, no positive cells could be detected within the tracheal mucosa (DCs in sections of identically fixed tumour tissues served as positive controls; not shown). In some cases paratracheal lymph nodes were present and served as internal positive controls for the detection of HLA-DR expression which was of particular interest in cases with no DCs in the mucosa (fig 1C). During respiratory infections a strong staining intensity for HLA-DR was found in all cases. Even the epithelium expressed this MHC class II molecule, indicating a strong inflammation induced activation (fig 1D). DCs were found in all children who had died of respiratory infections in numbers ranging from a few cells in the mucosa of the whole section to more than 10/mm basement membrane in the mucosa of the inflamed tissues. In children who had not died of respiratory tract infection no correlation with the cause of death or their age could be found.

HLA-DR stains of human paraffin sections from (A) a 78 year old woman (traumatic cause of death), (B) a 216 day old male infant (sudden infant death, SID), (C) a 64 day old male infant (SID), and (D) a 76 day old male infant (bronchopneumonia). Dendritic cells (DCs) are HLA-DR+ (blue) and have a dendritic morphology (arrows). They are located under the basement membrane within the epithelium (Ep) and lamina propria (Lp) of the respiratory mucosa (A, B, Ve = vessel). Other HLA-DR+ cells were macrophages and lymphocytes. A paratracheal lymph node (LN) with DCs and other HLA-DR+ cells served as an internal positive control in the case lacking DCs within the mucosa (C). In the case with respiratory infection (D) many DCs were found and the epithelium also expressed HLA-DR. (E) Spleen sections served as external controls. DCs are interspersed in the lymphatic tissue (insert: isotype control). DCs in the draining lymph node were found between HLA-DR stained lymphocytes (F, magnification of C). HLA-DR expression was found on DCs but also on epithelial cells during inflammation (G, magnification of D).
The ultrastructure was not preserved sufficiently. However, using transmission electron microscopy on the samples of the two additional patients, intraepithelial and subepithelial cells were detected with features characteristic of mucosal DCs (fig 2). These included long cell processes and nuclei with multiple invaginations. Some of the cells were found to encircle a lymphocyte with their cell processes or to lie in close contact to lymphocytes. Because of autolytic changes, Birbeck granules could not be demonstrated as proof of Langerhans' cells.

Transmission electron micrograph of a dendritic cell (DC) in close contact with a lymphocyte (L) in an 80 year old man with a traumatic cause of death. Both cells are located in the subepithelium. Cp = cell process of the dendritic cell, Ecm = extracellular matrix of the subepithelial connective tissue, arrows = invaginations of the nucleus.
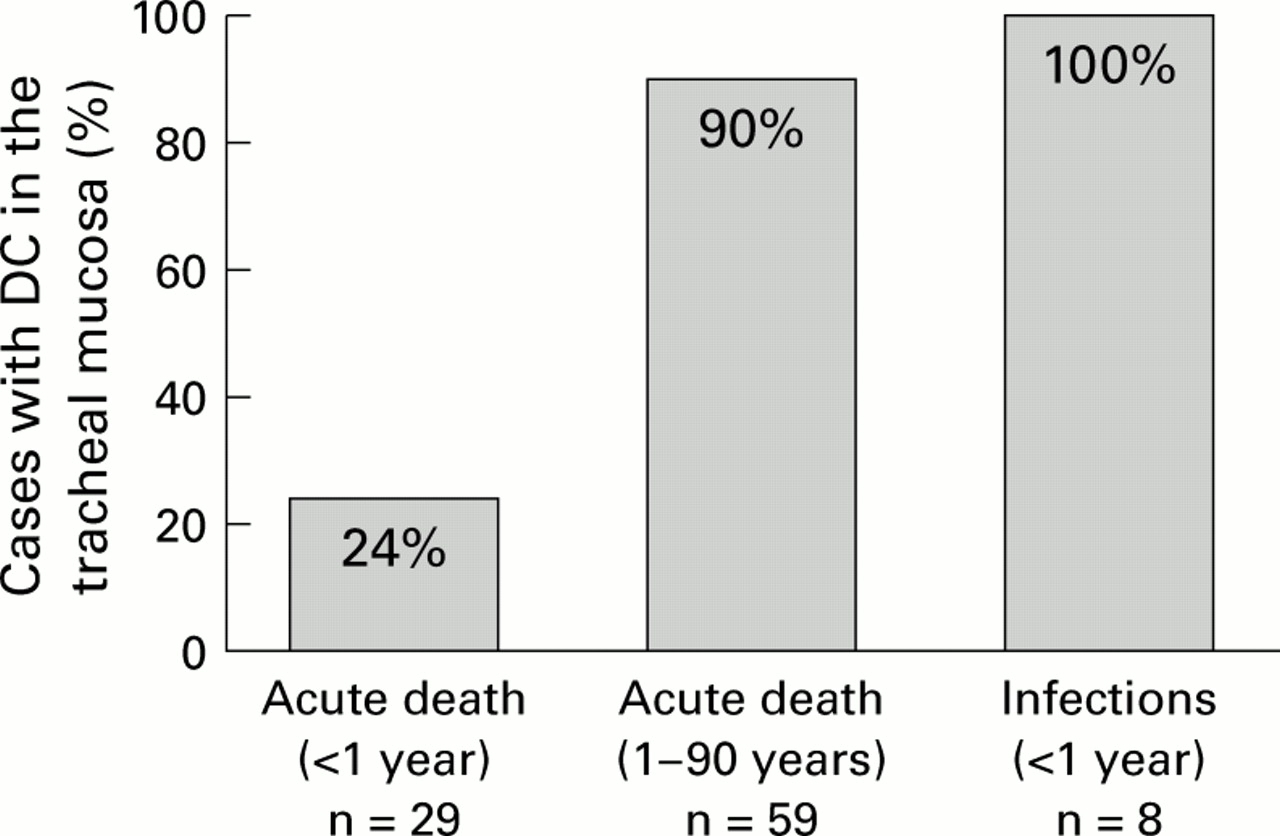
Of those patients who had died an acute death within the first year of life, most had no DCs within the tracheal respiratory mucosa. Only in seven of the 29 cases in this age group (24%) were DCs detected (fig3), all of whom had died of SID. The six children who had died of non-natural causes of death were all negative for DCs. An influence of age on the occurrence of DCs in the first year of life was not evident. In contrast, most of the older patients possessed DCs. In all children who had died of respiratory tract infections DCs were present (fig 4). The histopathological diagnosis as well as cytomegalovirus, influenza, adenovirus infections and a streptococcal infection were noted as the causes of death.
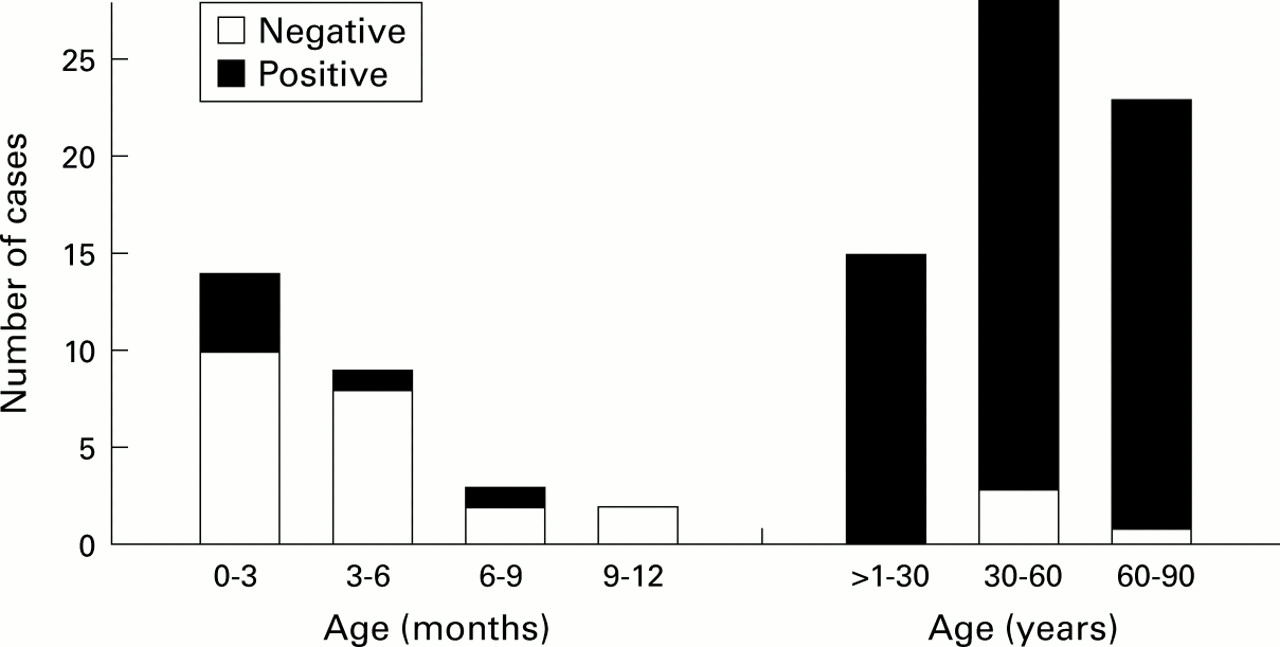
Number of cases with dendritic cells (DCs) in the mucosa in the first year of life and thereafter.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Occurrence of dendritic cells (DCs) in patients who had died acutely and those with respiratory tract infections. The cases of acute death are shown in two groups: <1 year and ⩾1 year of age.
Discussion
In this study human tissue of the respiratory tract was examined retrospectively. It was extremely difficult to collect an adequate number of cases who had died an acute death since necroscopic examination in these cases is rare. The data presented support the findings of other investigators who described DCs in human lung tissue and, for the first time, yield information on the initial occurrence and frequency of mature DCs in the human tracheal mucosa.
Animal data suggest an increase in bone marrow progenitor cells of DC and the immigration of these cells into different lung compartments.26 ,27 Since it has been shown that the number of DCs in the respiratory mucosa increases faster than neutrophil granulocytes after bacterial stimulation, local stimulation and ongoing proliferation might also be involved.28
Interestingly, our data are in line with recent findings in very young mice in whom few DCs were found in the airways. Resistance to pulmonary allergen sensitisation of these mice could be disrupted by co-exposure to pollutant aerosols leading to a marked increase in DC numbers.29
There are few other markers for the detection of DCs within the mucosal tissue on paraffin sections and these have disadvantages. The antibody against CD68 is also present on macrophages and should give results similar to HLA-DR. A subset of DCs has been described as positive for CD1a30 ,31 but others could not detect this molecule on human DCs.32 We found no CD1a+ DCs in our samples using two different antibodies. It could not be excluded that DC precursors were present in the mucosa of the cases lacking mature DCs. The presence and detection of DCs might be related to the post mortem interval but, as the time of death was uncertain in some cases (especially in SID), a correlation could not be determined.
This study extends the knowledge on the first occurrence of DCs in the human respiratory mucosa. The investigation of their distribution in the bronchi, bronchioli, and the interstitium, as well as their presence in the fetal mucosa, is of great interest and will be undertaken in further studies. Although time consuming, prospective studies should be performed with cryopreservation of material to extend the list of immunomarkers which recognise DCs in the human mucosa.
This study has shown that mature DCs are a regular part of the human tracheobronchial mucosa after the first year of life. Recurrent viral infections not resulting in severe injury or death are a common feature at this age and may have influenced the results. Mucosal DCs may complement and later substitute the antigen uptake of BALT which is absent in healthy adults.33 Bronchial DCs might be involved in processes of tolerance or allergy development. However, the mechanisms of such involvement and the influence of environmental stimuli as well as the genetic background of the patient remain to be elucidated. In contrast, many data exist on the development of secondary lymphoid tissue in experimental animals.34 It is known from animal studies that the immigration of DC precursors from the bone marrow is caused by mucosal infectious stimuli.28 ,35 This might also be true for patients suffering from severe infections. Thus DCs are regularly present in patients after the first year of life, which may be of relevance to the clinical concept of inhalation vaccination.
Acknowledgments
This study was supported by the Deutsche Forschungsgemeinschaft (Ts 56/1-1, Pa 240/8-1). The authors would like to thank Dr Peter Jecker for help with the CD1a staining, Karin Westermann and Sonja Seiter for skilful technical assistance, Dagmar Stelte for graphic work, and Sheila Fryk for correction of the English.
References
Narrative Based Medicine, An Interdisciplinary Conference Research, Narrative, and Practice A two day conference—Monday 3rd and Tuesday 4th September 2001 Homerton College, Cambridge, UK
BMJ Publishing Group
For full details contact: BMA/BMJ Conference Unit, Tavistock Square, London, WC1H 9JP Tel: +44 (0)20 7383 6819; fax: +44 (0)20 7383 6663; email: clyders{at}bma.org.uk. www.quality.bmjpg.com