Article Text

Abstract
BACKGROUND In the host defence of the lung neutrophils (PMN) play a central role. Apart from antimicrobial properties, recent data indicate that PMN also exert anti-inflammatory effects by stimulation and release of cytokine antagonists such as interleukin-1 receptor antagonist (IL-1ra).
METHODS Cytokine release from lipopolysaccharide stimulated whole blood was studied in 18 patients with community acquired pneumonia (CAP) and severe co-morbidities at admission and after 24 hours. Release of IL-1ra, interleukin-1β (IL-1β), tumour necrosis factor α (TNFα), soluble TNF receptor type I (sTNF-RI), and IL-8 was determined by ELISA.
RESULTS The mean (SD) leucocyte level at admission was 12.5 (4.1)/nl. There was a significant correlation between the release of anti-inflammatory cytokines such as IL-1ra and sTNF-RI and the leucocyte count at admission and after 24 hours. Additional in vitro experiments showed that co-incubation of peripheral blood mononuclear cells with autologous PMN led to a marked dose dependent increase in IL-1ra and sTNF-RI release.
CONCLUSION These results indicate that PMN may be responsible for the increase in anti-inflammatory cytokines in CAP. Strategies to increase neutrophil counts may exert beneficial effects, not only by augmenting the antimicrobial activity but also by modulating the inflammatory cytokine response.
- community acquired pneumonia
- neutrophils
- anti-inflammatory cytokines
Statistics from Altmetric.com
Community acquired pneumonia (CAP) remains a common cause of morbidity and mortality with an estimated incidence of 12 cases per 1000 persons per year. It is the sixth leading cause of death in the USA1 and in the UK it causes 10 times as many deaths as all other infectious diseases combined.2 Despite the development of many new antibiotics, the incidence and morbidity of pneumonia has not declined over the past 30 years.
An important risk factor for the development of pneumonia is the presence of severe underlying co-morbidities such as cardiac, cerebrovascular, liver, chronic lung or end stage renal disease, or diabetes mellitus. The mortality of CAP is particularly high in elderly patients; 90% of the deaths attributable to pneumonia occur in patients over 65 years.3
The recruitment and activation of leucocytes characterises the host defence of the lung, in which polymorphonuclear granulocytes (PMN) play a central role. Watanakunakorn et al 4 observed that, in pneumococcal pneumonia, a low or normal leucocyte count is associated with a higher fatality rate.
Regulation of the inflammatory response in bacterial pneumonia is dependent on complex interactions between immune cells and both pro-inflammatory and anti-inflammatory cytokines. Interleukin 1β (IL-1β) and tumour necrosis factor alpha (TNFα) are important pro-inflammatory cytokines and early response mediators which regulate a number of cellular functions and dictate events leading to the initiation, maintenance, and repair of tissue injury. Interleukin 1 receptor antagonist (IL-1ra) leads to a competitive occupation of the IL-1 receptor without agonist activity. It therefore has an inhibitory effect on IL-1 dependent inflammation. Soluble tumour necrosis factor receptor type I (sTNF-RI) represents a truncated form of the TNF receptor which has been shed off the cell surface. It neutralises the biological activity of TNF by competing with cell surface receptors for TNF binding.
The relationship between PMN and cytokines may influence the course of pneumonia in two ways:
- (1)
- Neutrophils are traditionally seen as end stage cells which mainly exert effector functions by releasing the content of their granules. Obviously, phagocyte dysfunction and insufficient recruitment of neutrophils to the inflamed site is associated with impaired clearance of pathogens. This can lead to an overactivation of the inflammatory cascade, with a predominance of pro-inflammatory cytokines followed by increased tissue damage and systemic inflammatory response syndrome (SIRS).5
- (2)
- Recently, apart from antimicrobial effects, the immunoregulatory activity of neutrophils has been established—for example, neutrophils can be driven to acquire dendritic cell characteristics6 and release anti-inflammatory cytokines such as IL-1ra.7 These immunoregulatory properties of neutrophils may contribute to the beneficial effect of the phagocyte response in bacterial pneumonia.
To determine whether PMN have a role in the anti-inflammatory cytokine response in patients with pulmonary infections we have examined the concentration of pro-inflammatory and anti-inflammatory cytokines in 18 patients admitted to hospital with CAP and correlated the results with leucocyte counts at admission and after 24 hours. We hypothesised that pneumonia patients with leucocytosis may show a stronger anti-inflammatory cytokine response than those without leucocytosis, and that this difference in cytokine levels may be related to leucocyte counts. To assess the effects of neutrophils on anti-inflammatory cytokine release more directly, additional in vitro experiments were performed with different mixtures of peripheral blood mononuclear cells (PBMC) and PMN.
Methods
PATIENTS
Eighteen patients (12 men) admitted with a diagnosis of community acquired pneumonia and severe co-morbidity were included in the study. Pneumonia was defined as the presence of new shadowing on the chest radiograph in conjunction with an appropriate clinical history and clinical and laboratory signs of infection (fever >38°C, purulent sputum, increased erythrocyte sedimentation rate, C reactive protein) for which no other cause was found. The mean age of the patients was 68 years (range 38–92) and they had at least one of the following co-morbidities: coronary artery disease (n=11), chronic renal failure (n=8), chronic heart failure (n=5), diabetes mellitus (n=6), chronic obstructive pulmonary disease (n=3), liver cirrhosis (n=2), lung fibrosis (n=1), bronchial asthma (n=1), Parkinson's disease (n=1), cerebral vascular disease (n=2).
All patients were treated by the same investigators and received similar supportive care and antimicrobial therapy. Informed written consent was obtained from all patients and the study was approved by the ethic committee of the Medical University Lübeck.
Severity of disease was assessed by the modified clinical pulmonary infection score (CPIS) as described originally by Puginet al.8 This score, with a range of 1–12 points, considers the following six criteria: (1) body temperature, (2) blood leucocyte count, (3) quality and quantity of bronchial secretion, (4) degree of oxygenation as described by the quotient Po 2/Fio 2, (5) infiltrates on the chest radiograph, and (6) findings of pathogenic bacteria in culture of respiratory secretions. Furthermore, we defined risk categories as described by Fine et al 9 which are based on age, co-morbidity, and abnormal physical and laboratory findings at presentation. This score enables patients to be stratified into five risk categories with respect to 30 day mortality.
WHOLE BLOOD CYTOKINE ASSAY
At admission and after 24 hours venous blood was drawn for whole blood cytokine measurements, which were performed as described previously.10 Briefly, 5 ml heparinised blood was diluted 1:10 with RPMI 1640 medium complemented with 200 mM glutamine and 10000E penicillin/streptomycin (Biochrom, Germany) and immediately stimulated for 24 hours with 1 μg/ml lipopolysaccharide (LPS) fromE coli serotype O26:B6 (Sigma, St Louis, MO, USA). Preliminary experiments with lipoteichoic acid (LTA; Sigma), a cell wall constituent of Gram positive bacteria, showed comparable results with regard to cytokine stimulation (data not shown). Samples were incubated in PPN tubes at 37°C and 5% CO2. Controls without LPS were performed in every experiment. Cell free supernatants were detached after 24 hours and stored at –80°C until the assay was performed.
Levels of the following cytokines were measured in the supernatants by commercially available ELISA kits according to the manufacturers' instructions: TNFα (lower detection limit 15.6 pg/ml; BioSource International, Camarillo, USA); sTNF-receptor p55 (lower detection limit 7.8 pg/ml; R&D Systems; Minneapolis, USA); IL-1β (lower detection limit 3.9 pg/ml; Bender MedSystems, Vienna, Austria); IL-1ra (lower detection limit 46.9 pg/ml; R&D Systems), and IL-8 (lower detection limit 16 pg/ml; Bender MedSystems).
IN VITRO EXPERIMENTS
To determine the contribution of neutrophils (PMN) to the release of anti-inflammatory cytokines, additional in vitro studies with different proportions of mononuclear cells (PBMC) and PMN were performed. Peripheral venous blood was drawn from five healthy volunteers with a mean (SD) age of 25 (1) years for PMN and PBMC isolation and culture. Cells were isolated using Ficoll-Hypaque density gradient centrifugation at 600g for 20 minutes as described previously.11 PBMC were suspended in RPMI 1640 medium with 5% autologous serum to a concentration of 106 cells/ml. PMN were stored in RPMI medium with 5% autologous serum at 4°C. The final preparation contained more than 98% of each cell type. Isolated cells were 99% viable as measured by trypan blue dye exclusion.
Aliquots of PBMC (500 μl each) were placed in the wells of a 24-well plate, immediately stimulated with 1 μg/ml LPS fromE coli serotype O26:B6, and incubated for 24 hours (37°C, 5% CO2). After four hours PMN (500 μl each) were added to the PBMC in the following proportions: 50% PBMC and 50% PMN, 33% PBMC and 67% PMN, 11% PBMC and 89% PMN. Each experiment also included controls without addition of PMN. Cell viability was >96% after 24 hours. Cell free supernatants were removed after 4, 8, 12 and 24 hours and stored at –80°C until assayed for sTNF-receptor p55 and IL-1ra by commercial ELISA kits as described above. Preliminary experiments showed that differences in cytokine release are not due to the differences in absolute cell concentrations or to an influence of toxic PMN mediators like oxygen radicals (data not shown).
STATISTICAL ANALYSIS
Correlations between leucocyte levels and cytokine concentrations or clinical and laboratory parameters of inflammation were tested using the Spearman rank correlation. The Wilcoxon signed rank test was used for paired samples in the in vitro experiments. Calculations were carried out with Statistica for Windows, Version 5. A value of p<0.05 was considered statistically significant.
Results
CLINICAL CHARACTERISTICS OF STUDY POPULATION
The mean (SD) white blood cell count of the study population was 12.5 (4.1) × 103/mm3 with a total PMN count of 10.5 (2.5) × 103/mm3. Microbiological findings were Streptococcus pneumoniae(n=9), Haemophilus influenzae (n=2), none (n=7). Mean body temperature was 38.4 (1.0)°C and the CPIS and Fine severity scores were 5.3 (1.6) and 103 (31), respectively. The C reactive protein (CRP) level was 169 (113) mg/l and Po 2/Fio 2 was 237 (74). Six patients were admitted to the intensive care unit and one patient died. Chest radiography showed lobar infiltrates in 11 cases and patchy infiltrates in seven.
There was no correlation between the demographic and clinical parameters and the leucocyte count. In particular, the risk categories and the severity of pneumonia as assessed by the Fine and CPIS scores were not dependent on the number of circulating leucocytes. Moreover, there was no apparent relationship between leucocyte counts on admission and the aetiology of the infection or the clinical outcome.
CYTOKINE CONCENTRATIONS
The cytokine antagonists IL-1ra and sTNF-RI showed a significant correlation with the leucocyte count at admission (IL-1ra:r=0.57, p<0.02; sTNF-RI:r=0.51, p<0.05) which remained unchanged after 24 hours (IL-1ra: r=0.58, p<0.02; sTNF-RI: r=0.54, p<0.04). TNFα levels also correlated strongly with the leucocyte count at admission (r=0.71; p<0.01) but after 24 hours no further relationship could be observed (r=0.16, p=0.55, NS, fig 1).
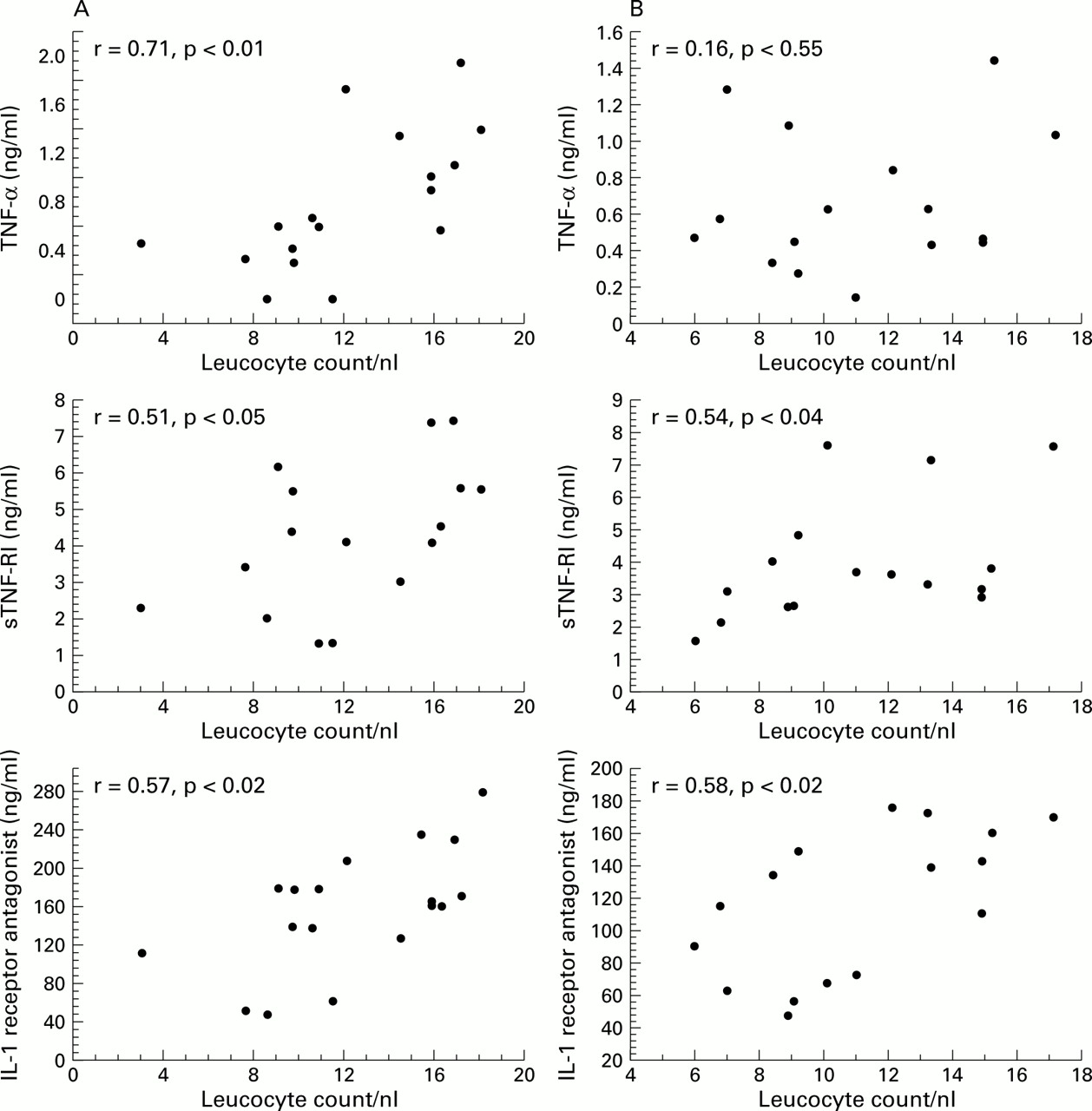
Correlations between cytokines and leucocytes (A) at admission and (B) 24 hours after admission.
At admission there was a correlation between TNFα and sTNF-RI levels (r=0.49, p<0.05) and between TNFα and IL-1ra levels (r=0.63, p<0.01). Furthermore, IL-1ra and sTNF-RI were correlated with CRP levels (IL-1ra: r=0.55, p<0.02; sTNF-RI:r=0.54, p<0.05), whereas only a trend was found for the correlation between TNFα and CRP levels (r=0.44, p=0.08). In contrast, IL-1β showed no relation to markers of inflammation but was the only cytokine which was significantly associated with the severity of disease as assessed by the CPIS score (r=0.48, p<0.05).
PMN AND ANTI-INFLAMMATORY CYTOKINES: IN VITRO FINDINGS
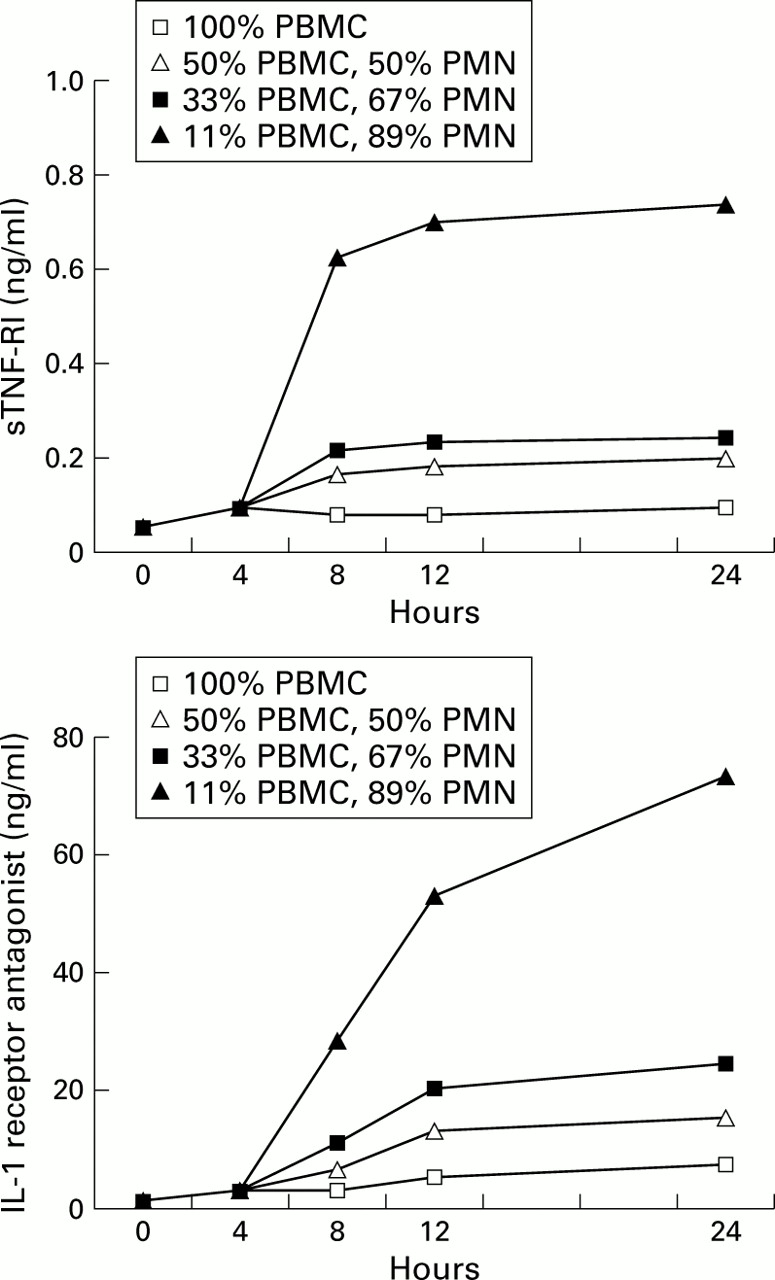
To define more clearly the relationship between PMN and anti-inflammatory cytokines we performed additional in vitro experiments. Incubation of isolated PBMCs from five healthy volunteers resulted in a relatively low release of sTNF-RI and IL-1ra which increased slightly during the 24 hour incubation period. Co-incubation with autologous PMN, however, led to a dose dependent increase in the median (SE) levels of IL-1ra after 24 hours from 7.64 (2.19) ng/ml (100% PBMC) to 15.38 (2.91) ng/ml (50% PBMC, 50% PM; p<0.05), 24.70 (4.95) ng/ml (33% PBMC, 67% PMN; p<0.05), and 73.41 (17.11) ng/ml (11% PBMC, 89% PMN; p<0.05). Similar increases were observed in sTNF-RI levels after 24 hours from 98.85 (7.19) pg/ml (100% PBMC) to 200.66 (17.61) pg/ml (50% PBMC, 50% PMN; p<0.05), 248.45 (27.69) pg/ml (33% PBMC, 67% PMN; p<0.05), and 740.60 (57.12) pg/ml (11% PBMC, 89% PMN; p<0.05, fig2).

{kind=link}
{kind=link}
Dose dependent increase of anti-inflammatory cytokine release by PBMC from healthy volunteers (median values from five experiments) after co-incubation with different concentrations of autologous PMN.
Discussion
The main finding of our study is that leucocyte counts in patients with CAP are associated with a preferential increase in levels of the anti-inflammatory cytokines IL-1ra and sTNF-RI. The correlation between the levels of these cytokines and the number of leucocytes may result from differences in the severity of disease. However, we found no association between the leucocyte count and the underlying condition, risk category, microbiological findings, or the clinical pulmonary infection score. Alternatively, PMN may be directly responsible for the strong anti-inflammatory host response.
Previous studies12 ,13 have shown that highly purified PMN are able to release IL-1ra but not IL-1β or TNFα. These cells produced more IL-1ra in response to LPS than PBMC. Moreover, recent data from our group14 have shown that co-incubation of PBMC with autologous PMN leads to significantly increased levels of IL-1ra after 12 and 24 hours, while levels of TNFα and IL-1β decline. In the present study we have extended these findings by demonstrating a quantitative relationship between clinically relevant proportions of PMN in peripheral blood cell populations and the release of IL-1ra and sTNF-RI. For instance, a neutrophil proportion of 89%, which is representative of most patients with acute pneumonia, led to a 10.6-fold increase in IL-1ra concentrations after 24 hours which indicates a predominant role for the neutrophil in regulating the release of this cytokine. This over-proportional increase may be due to autocrine and paracrine effects of neutrophil stimulation. Taken together, these in vivo and in vitro findings support our hypothesis that an increase in anti-inflammatory cytokines in pneumonia may be causally related to PMN concentrations.
A strong anti-inflammatory response in patients with severe CAP,15 adult respiratory distress syndrome (ARDS),16 and sepsis17 ,18 has been described previously. This may be explained by the finding that IL-1ra protein synthesis and release by leucocytes is stimulated by LPS, GM-CSF, TNFα, IL-1β, and IL-10 in vitro.19 However, in acute infections the cellular source of IL-1ra—which can be released by different cell types including monocytes, macrophages, PMN, and fibroblasts—has not been determined.
Van der Poll et al 20 observed a pronounced rise in IL-1ra concentrations (apparent after one hour, peaking after three hours) in four healthy volunteers after intravenous injection of rTNF. The rise in IL-1ra after administration of endotoxin in eight healthy chimpanzees started two hours later. Neutralisation of the early endotoxin induced TNF activity by anti-TNF caused a marked reduction in IL-1ra concentrations. These results are in line with our data, showing an early but short term rise in TNFα followed by a sustained increase in IL-1ra. This may indicate that TNF is an early trigger of the IL-1ra response in CAP.
TNFα showed a positive correlation with the leucocyte count but not with the severity of pneumonia as assessed by the CPIS. In contrast, IL-1β was not associated with leucocyte counts but was correlated significantly with the CPIS. It is therefore possible that IL-1β is a better marker of disease severity. However, Purenet al 21 found, in a more heterogenous patient population, that IL-1β was associated with the severity of infection but TNFα was a marker of the severity of pneumonia.
Low concentrations of anti-inflammatory cytokines in patients with acute lung injury are associated with a high mortality rate.16 Therapeutic administration of recombinant IL-1ra, which has shown favourable effects in experimental animal models,22 ,23 may be beneficial for patients with severe pneumonia and low leucocyte counts. Studies in humans have clearly established the importance of endogenous IL-1ra and sTNF-RI in counterbalancing the pro-inflammatory cascade and limiting subsequent organ damage, but the results of therapeutic trials of exogenous IL-1ra or TNF-receptor proteins were disappointing24 and have even been associated with increased mortality.25 An alternative approach may be to increase the amount of circulating neutrophils which, after transmigration to the inflammatory site, release the anti-inflammatory cytokines at the proper location. This may be achieved by the administration of granulocyte-colony stimulating factor (G-CSF) which has been shown to increase the levels of leucocytes and anti-inflammatory cytokines in both experimental models and in humans.26 ,27 Administration of G-CSF to non-neutropenic patients with CAP had no effect on survival but there were some benefits with regard to the rate of complications such as ARDS, disseminated intravascular coagulation, and multilobar spread in the treated group.28
In summary, our data indicate that leucocytosis may be beneficial for the infected host, not only by increasing the antimicrobial capacity but also by raising the levels of anti-inflammatory cytokines such as IL-1ra. Further studies are needed to evaluate the prognostic relevance of these findings and the efficacy of specific therapeutic interventions.