Article Text

Abstract
BACKGROUND Interleukin 5 (IL-5) has an important role in mobilisation of eosinophils from the bone marrow and in their subsequent terminal differentiation. A study was undertaken to determine whether inhaled and intravenous IL-5 could induce pulmonary eosinophilia and bronchial hyperresponsiveness (BHR) independently of these effects.
METHODS Nine mild asthmatics received inhaled (15 μg) or intravenous (2 μg) IL-5 or placebo in random order in a double blind, crossover study. Blood samples were taken before and at 0.5, 1, 2, 3, 4, 5, 24, and 72 hours following IL-5 or placebo, and bronchial responsiveness (PC20 methacholine) and eosinophil counts in induced sputum were determined.
RESULTS Serum IL-5 levels were markedly increased 30 minutes after intravenous IL-5 (p=0.002), and sputum IL-5 levels increased 4 and 24 hours after inhaled IL-5 (p<0.05). Serum eotaxin was raised 24 hours after intravenous IL-5 but not after inhaled IL-5 or placebo. Blood eosinophils were markedly reduced 0.5–2 hours after intravenous IL-5 (p<0.05), followed by an increase at 3, 4, 5, and 72 hours (p<0.05). Sputum eosinophils rose significantly in all three groups at 24 hours but there were no differences between the groups. Bronchial responsiveness was not affected by IL-5.
CONCLUSION The effects of IL-5 appear to be mainly in the circulation, inducing peripheral mobilisation of eosinophils to the circulation without any effect on eosinophil mobilisation in the lungs or on bronchial responsiveness.
- asthma
- eosinophil
- interleukin 5
- bronchial responsiveness
Statistics from Altmetric.com
Chronic asthma is characterised by an inflammation of the bronchi with the presence of eosinophils and CD4+ T helper cells.1 ,2 T helper type 2 (Th2) cells are the predominant source of the cytokine interleukin (IL)-5 and are increased in the asthmatic airways.2 IL-5 plays an important role in the mobilisation of eosinophils from the bone marrow and in the terminal differentiation and maintenance of mature eosinophils.3IL-5 levels have been measured in the circulation of asthmatics patients4 and rises during asthma exacerbations.1 Additionally, increased IL-5 mRNA expression is seen in bronchial mucosa and bronchoalveolar lavage (BAL) CD4+ T cells in asthma,2 ,5 and IL-5 protein levels increase following allergen challenge.6 A positive correlation between asthma severity and IL-5 mRNA levels in bronchial biopsy specimens has also been demonstrated.7
In guinea pig and rodent models airway administration of IL-5 induces an increase in airway eosinophils without demonstrable change in bronchial responsiveness.8 ,9 Airway and blood eosinophilia has also been shown in IL-5 transgenic mice, but without bronchial hyperresponsiveness.10-12 IL-5 administration to the airways of asthmatic subjects, however, has been associated with peripheral blood eosinophilia, airway eosinophilia, and bronchial hyperresponsiveness.13 ,14
These conflicting observations suggest a complex cooperative role for IL-5 in the accumulation of circulating and airway eosinophils and their possible subsequent contribution to bronchial hyperresponsiveness. Since IL-5 also appears to have important effects on the bone marrow,9 ,15 ,16 we questioned whether the route of IL-5 production was crucial to its effects on the airways. We hypothesised that IL-5 confined entirely to the lungs and airways would have little systemic effect on eosinophil mobilisation, while IL-5 administered to the circulation might possess this primary effect. Further, we sought to establish whether pulmonary or systemic IL-5 could induce specific pulmonary effects consistent with asthma. We therefore investigated the effects of IL-5 administered intravenously or by inhalation to patients with mild asthma on eosinophil counts in blood and in sputum, and on airway hyperresponsiveness.
Methods
SUBJECTS
Nine non-smoking atopic patients with mild persistent asthma (five women, age range 25–41 years) participated in the study (table 1). All subjects had a history of episodic chest tightness, and wheezing and symptoms were controlled by on demand usage of short acting inhaled β2 agonists alone. All subjects had been free of symptoms of respiratory tract infection for at least 2 weeks before study commencement. Atopy was determined by positive skin prick responses (>3 mm) to extracts of six common aeroallergens (Vivodiagnost, ALK, Benelux). Baseline forced expiratory volume in 1 second (FEV1) was >70% of predicted17 and bronchial hyperresponsiveness (PC20 <8 mg/ml methacholine) was determined by a standardised technique.18 The protocol was approved by the ethics committee of the Royal Brompton Hospital and all participants gave written informed consent.
Characteristics of study subjects
PROTOCOL
The study was of a placebo controlled, double blind, crossover design and consisted of three periods (fig 1) separated by washout periods of at least 2 weeks. At a screening visit baseline PC20 and sputum induction were performed. In each period the subjects visited the laboratory on three separate days. On day 1 of each period all subjects received an intravenous and an inhaled administration of either IL-5 or placebo, or both administration routes contained placebo. Blood eosinophils were determined before and 0.5, 1, 2, 3, 4, 5, 24, and 72 hours after administration. PC20 and sputum induction were performed at 4, 24 and 72 hours following each administration.
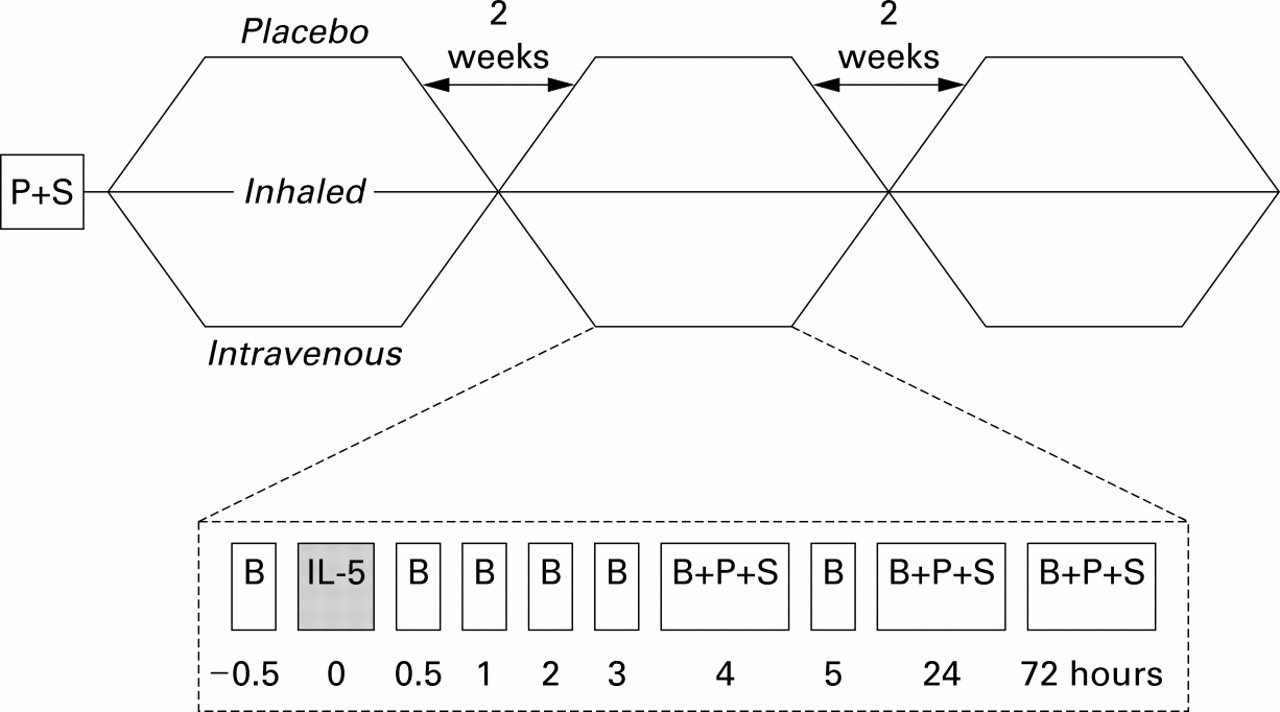
Study design. Subjects were treated in random order with inhaled IL-5 (15 μg), intravenous IL-5 (2 μg), or placebo in a crossover design at 2 week intervals. Specimen sampling was performed at time points noted. B = blood sample; P = PC20methacholine; S = sputum induction.
IL-5 ADMINISTRATION
Recombinant human IL-5 (Genzyme, Boston, MA) was reconstituted in 0.9% NaCl (2 ml) and administered at a dose of 2 μg intravenously via a slow infusion in a forearm vein over a period of 5 minutes. This intravenous dose was calculated according to the levels of IL-5 measured in circulating blood of patients with asthma during an exacerbation.1 IL-5 (15 μg) was inhaled over 5 minutes via a nebuliser (Mecic-aid, Pagham, Sussex, UK) from a mouthpiece using a one way exhaust valve (LC Plus, Pari, Surrey, UK). To ensure complete inhalation of IL-5 the chamber was then refilled with 2 ml 0.9% NaCl and the nebulisation was repeated.
LUNG FUNCTION AND METHACHOLINE CHALLENGE
Baseline FEV1 was recorded from the best of three attempts using a dry wedge spirometer (Vitalograph, Buckingham, UK).17 Spirometric tests were performed prior to and hourly after the administration of IL-5 or placebo until the start of methacholine challenge at 4 hours.
Methacholine challenges were performed according to a standardised methodology.18 After a nebulised saline challenge, doubling doses of methacholine (0.06–32 mg/ml) were administered via a dosimeter (Mefar, Bovezzo, Italy) with an output of 100 μl. A total of five inhalations of each concentration was administered (inhalation time 1 second, breath holding time 6 seconds). FEV1 was measured 2 and 3 minutes after each dose. The test was discontinued if FEV1 decreased by 20% or more than with the control inhalation (0.9% NaCl). The PC20 was calculated by log-linear interpolation of the last two data points.
PERIPHERAL BLOOD EOSINOPHIL COUNTS
Peripheral blood eosinophils were identified by the combination of peroxidase staining and side scatter and counted using the automated Advia 120 Hematology System (Bayer, Newbury, Berks, UK).
SPUTUM INDUCTION
All subjects received 200 μg salbutamol following the methacholine challenge. After recovery, sputum was induced according to a previously described method.19 Hypertonic saline aerosols (NaCl 3.5%) were nebulised at room temperature via an ultrasonic nebuliser (DeVilbiss Ultraneb 2000) at maximum output. Subjects inhaled the aerosols during three 5 minute intervals. Between each interval, or as soon as the subjects started coughing, they were instructed to wash out their mouth and blow their nose in order to minimise salivary contamination. The induced sputum was collected into a 50 ml tube, stored at 4°C, and processed within 2 hours.
SPUTUM PROCESSING
The whole sputum sample was diluted with 2 ml Hank's balanced salt solution (HBSS) containing 0.25% dithiotrietol (DTT; Sigma Chemicals, Poole, UK). The sample was gently mixed at room temperature, the volume determined, and the sample further diluted with HBSS to 10 ml. The sample was centrifuged (350g, 10 minutes), supernatant was removed, and the cell pellet was resuspended in 1 ml HBSS. Total cell counts were determined on a haemacytometer slide. Cytospin slides were prepared (600 rpm, 6 minutes; Shandon, Runcorn, UK) and stained with May-Grunwald-Giemsa. Differential cell counts were performed by a blinded observer with 300 non-squamous cells counted on each of two slides. To correct for the variable salivary contamination, differential cell counts were expressed as a percentage of 300 nucleated cells, excluding squamous epithelial cells. An adequate sample was defined if there was <80% squamous cell contamination. Supernatants were stored at –80°C.
IL-5 AND EOTAXIN ASSAYS
Serum IL-5 and eotaxin and sputum IL-5 levels were measured by ELISA according to the manufacturer's instructions (Pharmingen, Cambridge, UK). Briefly, purified rat anti-human monoclonal antibodies were incubated at 2 μg/ml in coating solution (0.1 M NaHCO3 (BDH), pH 8.2 at 4°C) overnight. Plates were washed, then blocked with 10% FCS for 2 hours at room temperature. Sample supernatants were added to each plate in duplicate. Cytokine standards (R&D Systems, Abingdon, Oxon, UK) were diluted in 10% (v/v) FCS/PBS/Tween and added in duplicate. The plates were then incubated at 4°C overnight, biotinylated rat anti-human antibody (R&D) was added, and they were incubated at room temperature for 45 minutes, then washed before the addition of avidin-peroxidase. The plates were incubated at room temperature for 30 minutes before washing with PBS/Tween and (ABTS, Sigma) substrate. They were then developed for approximately 10 minutes before measurement on a plate reader (Anthos Labtec, Austria) at 405 nm. The detection limit of this assay was 35 pg/ml.
DATA ANALYSIS
PC20 was log transformed before statistical analysis and reported as geometric mean (SD) values. IL-5 levels and blood and sputum eosinophils were not normally distributed and were therefore log transformed before analysis and reported as median (range) for eosinophils and geometric mean (SD) for IL-5 levels. Analysis of variance (ANOVA) for repeated measures was used for between group analyses. Within group changes in PC20, sputum and blood eosinophil levels, and serum and sputum supernatant IL-5 were analysed using the Student's pairedt test. Mean differences between treatments with 95% confidence intervals for these effect estimates are presented for serum IL-5 and blood eosinophil counts. All statistical analyses were performed using the SPSS program; p values <0.05 were considered statistically significant.
Results
SERUM AND SPUTUM IL-5 LEVELS
Serum IL-5 levels were rapidly cleared from the circulation and returned to baseline levels between 1 and 72 hours. Serum IL-5 concentrations in atopic asthmatic patients increased significantly 0.5 hours after intravenous IL-5 administration (387.4 (162.1) pg/ml) compared with baseline (14.0 (12.8) pg/ml, p=0.002) with rapid clearance, returning to baseline levels between 1 and 72 hours (fig 2). The serum IL-5 levels at 1, 2, 3, 4, and 5 hours after intravenous IL-5 were 16.8 (5.9), 14.2 (8.4), 5.7 (3.5), 8.1 (3.9), and 3.7 (2.0) pg/ml respectively, indicating that by 1 hour the levels had returned towards baseline values. In order to determine whether there was any difference in the kinetics of clearance of IL-5 between asthmatic patients and non-asthmatic controls, we also studied three non-asthmatic non-atopic volunteers. The mean (SE) serum level in non-asthmatic subjects was 3.6 (2.5) pg/ml at baseline, which increased to 54.2 (24.6) pg/ml 0.5 hours after intravenous injection of 2 μg IL-5 (mean difference –60.53, 95% CI –107.7 to –13.3, p<0.05; fig 2). These data indicate that there is no difference in the clearance in IL-5 between normal and asthmatic volunteers, although the asthmatic subjects had higher levels of baseline serum IL-5 than non-asthmatics. There was a trend towards an increase in serum IL-5 levels 0.5 hours after inhaled IL-5 administration (from 18.18 (12.4) pg/ml to 51.14 (100.56) pg/ml) in the asthmatic subjects, but placebo had no effect on serum IL-5 levels.

(A) Mean IL-5 serum levels. Serum IL-5 levels rose markedly following intravenous IL-5 but were not significantly increased by inhaled IL-5 or placebo in patients with mild asthma. In addition, in normal volunteers the administration of intravenous IL-5 led to a peak of serum IL-5 levels at 0.5 hours similar to that in patients with asthma. (B) Mean IL-5 levels in sputum supernatants. Sputum IL-5 levels rose 4 hours after inhaled IL-5 and remained raised at 24 hours but were not affected by intravenous IL-5 or placebo.
There was a significant increase in the IL-5 concentration measured in sputum supernatant from baseline at 4 and 24 hours following inhaled IL-5 (baseline 49.0 (16.4), 4 hours 2067 (1091), 24 hours 152.3 (42.8) pg/ml; p=0.04 and p=0.02, respectively), but no changes in sputum supernatant IL-5 following placebo or intravenous IL-5.
SERUM EOTAXIN
Serum eotaxin levels were 207.5 (80.2) ng/ml at baseline, 248.1 (93.6) at 0.5 hours, 232.5 (101) at 4 hours, and 301.4 (113.6) at 24 hours following intravenous IL-5. There was a trend to increased serum eotaxin levels following intravenous IL-5 with a 43.0 (23.3)% increase at 24 hours compared with baseline (fig 3). Comparative eotaxin levels were 12.0 (23.8) ng/ml following inhaled IL-5 and 1.5 (6.8) ng/ml following placebo. No correlation was observed between peak IL-5 levels (0.5 hours) and eotaxin levels at 0.5 or 24 hours following IL-5, nor was there correlation between eotaxin levels and peripheral blood lymphocyte or eosinophil numbers.

Serum eotaxin. There was a non-significant trend to increased serum eotaxin levels 24 hours after intravenous IL-5 but not after placebo.
EFFECT OF IL-5 ON PERIPHERAL BLOOD CELLS
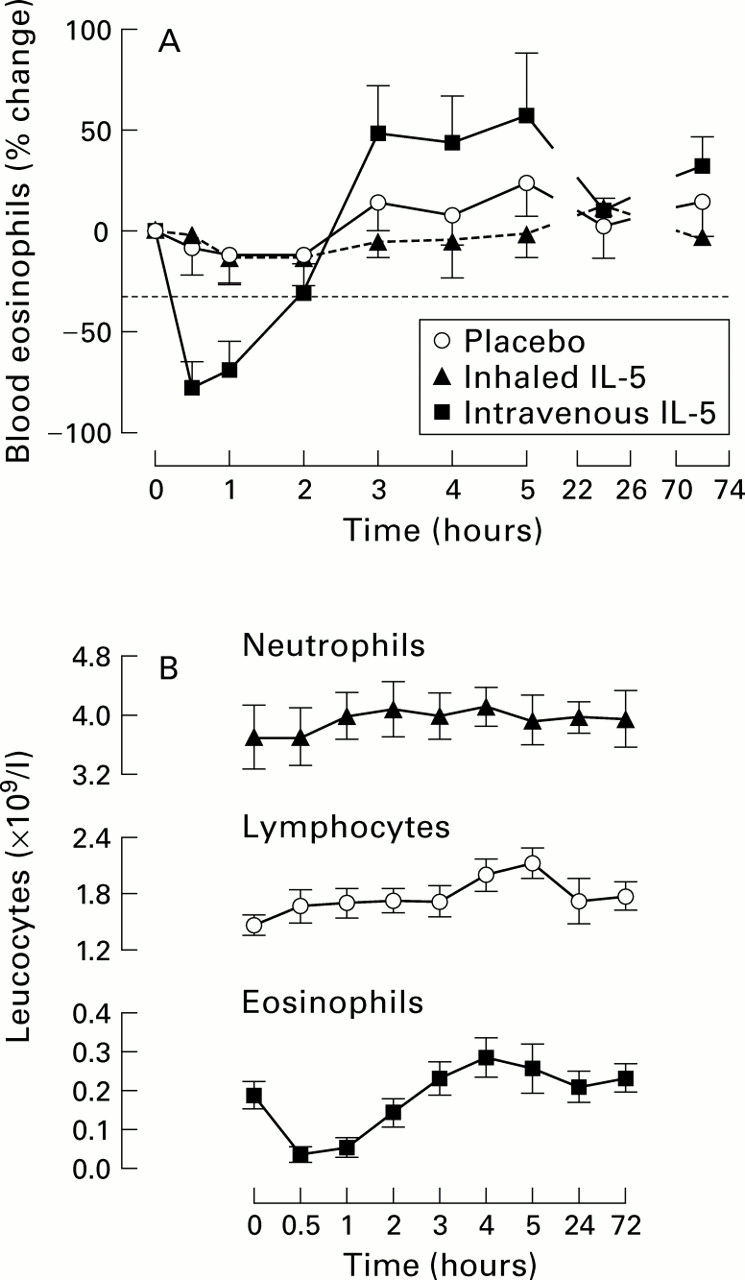
Blood eosinophil counts were reduced 30 minutes (mean difference 0.15, 95% CI 0.05 to 0.26, p<0.01) and 2 hours (0.13, 95% CI 0.02 to 0.24, p<0.05) following the administration of intravenous IL-5 compared with baseline (fig 4). The decrease in eosinophils 0.5 hours after intravenous administration was significantly different from changes following inhaled IL-5 and placebo (p<0.03). The increase in blood eosinophil counts observed at 3, 4, 5, and 72 hours was not statistically significant.

(A) Time course of blood eosinophil counts after placebo, inhaled and intravenous IL-5. Intravenous IL-5 induced a rapid fall in circulating eosinophil numbers (p<0.05) which was not seen with inhaled IL-5 or placebo. There was a prolonged but non-significant increase in eosinophils at 3, 4, 5, and 72 hours after intravenous IL-5. (B) Time course of changes in blood neutrophil, lymphocyte, and eosinophil counts after intravenous IL-5. Data shown are mean (SE).
In addition, peripheral blood lymphocytes were significantly increased from baseline at 4 hours (p<0.01, mean difference –0.54, 95% CI –0.97 to –0.11) and 5 hours (p<0.001, –0.59, 95% CI –1.02 to –0.16) following intravenous but not inhaled IL-5 or placebo.
EFFECT OF IL-5 ON SPUTUM EOSINOPHILS
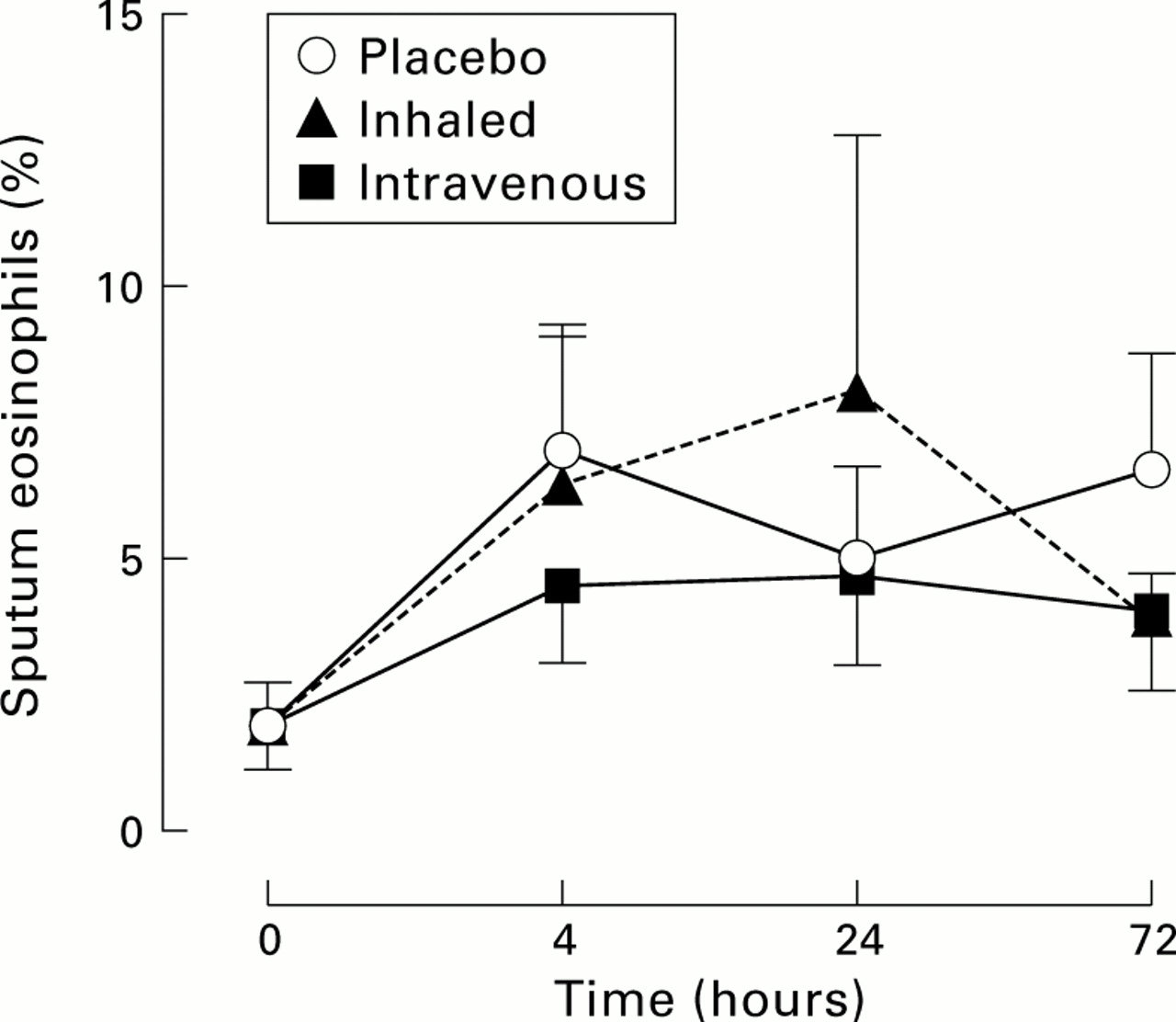
The median sputum eosinophil counts were 1.0% (range 0–6.3%) at baseline, increasing to 5.0% at 24 hours after inhaled IL-5 (p=0.02) and persisting at 72 hours (4.6%, p=0.02). However, similar increases were observed after intravenous IL-5 and after placebo treatment (median at 24 hours: 3.6% and 3.7%, respectively, p<0.01). There were no significant differences in sputum eosinophils between the treatment groups (fig5).

Mean sputum eosinophil counts. Sputum eosinophils were raised 4 hours from baseline in each of the treatment groups but there were no differences between groups at any of the time points measured.
EFFECT OF IL-5 ON LUNG FUNCTION AND BRONCHIAL RESPONSIVENESS
Baseline FEV1 was 95.1 (3.8)% predicted and PC20 was 1.15 (2.21) mg/ml. No significant changes were observed in FEV1 or PC20 during the three study periods (fig 6). There were no significant correlations between sputum IL-5 levels and change in PC20, nor between eosinophils in blood or sputum and levels of IL-5 measured in sputum or serum.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mean log PC20 to methacholine. Bronchial responsiveness did not change significantly from baseline after inhaled IL-5, intravenous IL-5, or placebo.
Discussion
We report the first observations on the comparative effects of inhaled and intravenous IL-5 in mild asthma. Significant increases were seen in IL-5 levels in the serum but not in the sputum when administered intravenously, and the other way round when given by inhalation. Thus, inhaled IL-5 appeared to be confined to the lung and intravenous IL-5 to the blood compartment. Intravenous IL-5 induced a rapid decline and subsequent non-significant rise in blood eosinophils which was not associated with airway eosinophilia or bronchial hyperresponsiveness. Sputum eosinophils were indeed slightly increased after intravenous and inhaled IL-5, but were also raised following placebo, which indicates that these increases were non-specific. Inhaled IL-5 had no systemic effect as reflected by blood eosinophil counts or serum eotaxin levels and did not affect bronchial responsiveness. A trend to increased serum eotaxin levels was observed 24 hours following intravenous IL-5. The cellular source of this increase in eotaxin levels could have been the vascular endothelial cells or circulating eosinophil and T cells. Systemic IL-5 is a potent stimulus to eosinophil mobilisation to the circulation, albeit not in itself sufficient specifically to induce lung eosinophilia (as measured in induced sputum) or airway hyperresponsiveness.
We observed significant, sequential, and converse effects on peripheral blood eosinophils following intravenous IL-5. Between 0.5 and 2 hours eosinophils were markedly reduced, and this effect has not been previously reported in animals or humans. At later time points (4–72 hours) there was a significant increase in blood eosinophil counts. We speculate that these effects result from rapid vascular margination followed by either demargination or replenishment of the circulating eosinophil pool from bone marrow and other extramedullary leucocyte reservoirs. A rapidly induced but short lived peripheral eosinophilia has been observed previously following intravenous administration of IL-5 in animals,20 ,21 while a prolonged peripheral blood eosinophilia occurred after inhaled IL-5 in asthmatic subjects.13
The finding that IL-5 did not specifically affect sputum eosinophils levels contradicts a previous study of IL-5 inhalation in patients with mild asthma.14 We observed minor but significant increases in sputum eosinophil numbers in all groups. These changes are unlikely to be explained by methacholine challenge22 or repeated sputum induction.23 Our findings are, however, consistent with previous studies in rodents. Wang et al 24 showed that circulating IL-5 is essential for the development of airway eosinophilia, whereas local lung IL-5 production did not lead to eosinophil recruitment to the lungs. Similarly, Mould et al 11observed a doubling of eosinophil counts in BAL fluid following the administration of intrapulmonary and intravenous IL-5 compared with intrapulmonary IL-5 administration alone in mice. Lee and coworkers12 created an IL-5 transgenic mouse expressing IL-5 in airway epithelium which developed baseline bronchial hyperresponsiveness and BAL eosinophilia; however, in this model IL-5 was expressed at unusually high levels in the serum with concomitant prominent peripheral eosinophilia.
We saw no effect of IL-5 on bronchial responsiveness. Consistent findings are provided in human studies in which monoclonal anti-IL-5 blocking antibodies substantially reduced allergen induced blood and sputum eosinophilia but had no effect on the late phase response and bronchial hyperresponsiveness to allergen challenge.25These findings have been consistently confirmed in animal models.26 ,27 The contribution of IL-5 to bronchial hyperresponsiveness is contentious and in asthma models it is difficult to dissociate from effects induced by mediators other than IL-5.28-31 Consistently, however, IL-5 transgenic mice have prominent airway eosinophilia but have acetylcholine responses similar to wild type mice.32 These studies collectively suggest a close association between IL-5 and both circulating and bronchial eosinophil numbers, but a dissociation between pulmonary IL-5 and bronchial responsiveness in both animals and humans.25 ,31 ,33
Our observations are substantially different from those of Shiet al 14 in terms of sputum eosinophilia and bronchial responsiveness induced by IL-5, despite using similar amounts of inhaled IL-5 obtained from a similar source and comparable cohorts of asthmatic patients in terms of severity and baseline sputum eosinophilia. These studies were performed in ethnically different populations, raising the possibility that there may be racial susceptibility to the effects of IL-5. Another possibility is that the high level of endemic parasitic colonisation in China34-36 may induce an expanded population of activated tissue eosinophils35-37 that are primed to respond more to IL-5. Sputum ECP levels in the mild asthmatics studied by Shiet al were, however, 2-log fold higher than levels previously described in severe asthma.38
Collins et al 9 reported that, in contrast to intravenous administration, local IL-5 did not induce tissue accumulation of eosinophils. This indicates that local IL-5 has little chemoattractant activity, whereas intravenous IL-5 has a rapid enhancing effect on eosinophil accumulation by stimulating the release of a rapidly mobilisable pool of bone marrow eosinophils. Furthermore, although IL-5 is necessary, other signals from activated T lymphocytes may also be required to induce accumulation of eosinophils in the airways.20 ,39 ,40 Eotaxin, an eosinophil selective chemokine, augments the accumulation of eosinophils by IL-5 in a synergistic fashion. This implies that IL-5 and eotaxin act cooperatively to promote the recruitment of eosinophils into the tissue.9 ,20 Our data indicate that, when administered on its own to the airways by aerosol, IL-5 is not capable of inducing eosinophilic inflammation and bronchial hyperresponsiveness in mild asthmatic subjects. However, changes in circulating eosinophil numbers were observed with intravenous administration of IL-5, indicating potential systemic effects on the bone marrow. Thus, IL-5 may have systemic rather than pulmonary effects in asthma.
Acknowledgments
The authors thank GlaxoWellcome for their support of this study.