Article Text

Abstract
BACKGROUND There is increasing evidence that the cytokine network is central to the immunopathology of inflammatory airway diseases. The interleukin 1 (IL-1) receptor antagonist (IL-1RN) is a naturally occurring anti-inflammatory agent that binds to the IL-1 receptor but does not possess agonist activity. Each of the genes of the IL-1 locus on chromosome 2q14 is polymorphic. The IL1RNgene contains an 86 bp tandem repeat and allele 2 of this polymorphism has been associated with various inflammatory diseases. The IL-1β (IL1B) gene contains a promoter polymorphism (C-511T) that has been associated with inflammatory diseases and is in linkage disequilibrium with the IL1RNpolymorphism.
METHODS We investigated whether polymorphisms in theIL1B and IL1RNgenes were associated with rate of decline of lung function. Genotypes were determined in 284 smokers with a rapid decline in lung function and 306 smokers with no decline in lung function.
RESULTS None of the genotypes was associated with the rate of decline of lung function. However, the distribution of IL1B/IL1RNhaplotypes was different between smokers with a rapid decline in lung function and those with no decline in lung function (p=0.0005).
CONCLUSION These results suggest that IL1B/IL1RN haplotypes play a role in the rate of decline in lung function in smokers.
- interleukin 1
- lung function
- smoking
- polymorphism (genetics)
Statistics from Altmetric.com
Interleukin 1 (IL-1) and similar cytokines play a central role in chronic inflammatory diseases.1 The IL-1 family consists of two pro-inflammatory cytokines, IL-1α and IL-1β, and a naturally occurring anti-inflammatory agent, the IL-1 receptor antagonist (IL-1RN). The two forms of IL-1 are the products of different genes, but they are structurally related and bind to the same receptor. They are synthesised by a variety of cell types, mainly monocytes and macrophages. IL-1RN is a 16–18 kD protein that binds to the IL-1 receptor with the same affinity as IL-1 but does not possess agonist activity and therefore acts as a competitive inhibitor of IL-1.2 IL-1 and its antagonist may influence the rate of decline in lung function because of the effects of IL-1 on neutrophil function and chemotaxis. IL-1 stimulates the synthesis of IL-8 which is a potent chemotactic agent for neutrophils and it also induces release of neutrophil elastase.3 IL-1 promotes the adhesion of neutrophils and other cells by enhancing the expression of adhesion molecules such as ICAM-1, VCAM-1, andl-selectin.4 ,5
Evidence for a contribution of the IL-1 gene complex to the pathogenesis of inflammatory airway disease was provided by measurements of IL-1 in asthmatic subjects. IL-1β expression in the alveolar macrophages of asthmatic subjects is upregulated6and there is increased protein expression of both IL-1β and IL-1RN in biopsy specimens of the bronchial epithelium of asthmatics.7 In animal models, intratracheal administration of IL-1β to Brown-Norway rats leads to an increased number of neutrophils in bronchoalveolar lavage fluid and increased airway hyperresponsiveness to bradykinin.8 Pretreatment with IL-1RN reduces bronchial hyperreactivity in guinea pigs.9
The genes of the IL-1 complex map to the long arm of chromosome 2,10 and each of the genes is polymorphic. The IL-1β gene (IL1B) has a single nucleotide polymorphism in the promoter region (C–511T)11 and theIL1RN gene has a penta-allelic polymorphic site in intron 2 containing 2–6 tandem repeats of an 86 bp sequence.12 There is evidence that allele 2 of theIL1RN gene (IL1RN*2) is associated with increased susceptibility or more severe outcome in chronic inflammatory diseases such as ulcerative colitis, systemic lupus erythematosus, and alopecia areata.13-17 The IL1B C–511T has been associated with inflammatory bowel disease18 as well as plasma levels of IL-1β and IL-1RN.19
Chronic obstructive pulmonary disease (COPD) is a chronic inflammatory disease that is associated with progressive airway remodelling and obstruction. The role of IL1B andIL1RN polymorphisms in COPD has only been investigated in a small number of subjects.20 IL1RN*2 has been associated with non-atopic asthma in a Japanese population,21 but not in black or white South African individuals.22 We hypothesised that carriers of IL1RN*2 andIL1B –511T who smoked would have more severe airflow obstruction and a more rapid decline in lung function than smokers who did not have these alleles.
Methods
SUBJECTS
Subjects were selected from the participants in the NHLBI Lung Health Study. The design of this multicentre randomised clinical trial is described more extensively elsewhere.23 Study participants were healthy current smokers aged 35–60 who had mild to moderate airflow obstruction (forced expiratory volume in 1 second (FEV1) 55–90% of predicted). Exclusion criteria included serious illnesses such as cancer, heart attack, stroke, or other important conditions that required medical treatment. The primary outcome was rate of decline of FEV1 over a follow up period of 5 years. Of the 3216 persistent smokers in this cohort, 284 were chosen for having the most rapid decline in lung function (decline in FEV1 >3.0% predicted per year), and 306 were selected because of absence of decline over the same period (increase in FEV1 >0.4% predicted per year). All participants were white.
GENOTYPING
DNA was extracted from blood samples using a standard phenol/chloroform protocol.24 The promoter region ofIL1B contains a single nucleotide repeat at position –511 (C→T). The following primers flanking this region were used to amplify the polymorphic region by PCR: 5′ TGG CAT TGA TCT GGT TCA TC (sense) and 5′ GTT TAG GAA TCT TCC CAC TT (antisense).11 The 306 bp polymerase chain reaction (PCR) product contains a cut site for DdeI resulting in two fragments of 146 and 160 bp, respectively. The C allele contains an additional cut site resulting in three fragments of 160, 114 and 32 bp, respectively. Polymerase chain reaction conditions were as follows: 40 cycles at 94°C for 30 seconds, 57°C for 30 seconds, and 72°C for 20 seconds.
Intron 2 of the IL-1RA gene contains a variable number of tandem repeats of an 86 bp length of DNA. The following primers flanking this region were used to amplify the polymorphic region by PCR: 5′ CTC AGC AAC ACT CCT AT (sense) and 3′ TCC TGG TCT GCA GGT AA (antisense).12 Polymerase chain reaction conditions were as follows: 35 cycles at 94°C for 30 seconds, 60°C for 30 seconds, and 72°C for 20 seconds.
The size of the amplified products was determined by electrophoresis on a 2% agarose gel stained with ethidium bromide for visualisation with ultraviolet light. Negative controls without DNA template were included with each reaction. All analyses were performed blind with respect to subject characteristics. Genotypes were confirmed by a second person not directly involved in the study.
STATISTICAL ANALYSIS
The results are presented as genotype and allele frequencies for both subjects with a rapid decline in lung function and those with no decline in lung function among the participants in the Lung Health Study. χ2 tests for statistical significance were performed on allele frequencies. Logistic regression was used to adjust the results for potentially confounding variables such as age, sex, cigarette smoking, baseline lung function, and response to methacholine. Unpaired t tests were used to compare baseline values. Haplotype frequencies and linkage disequilibrium were estimated using the expectation-maximisation (EM) algorithm implemented in the Arlequin software package.25
Results
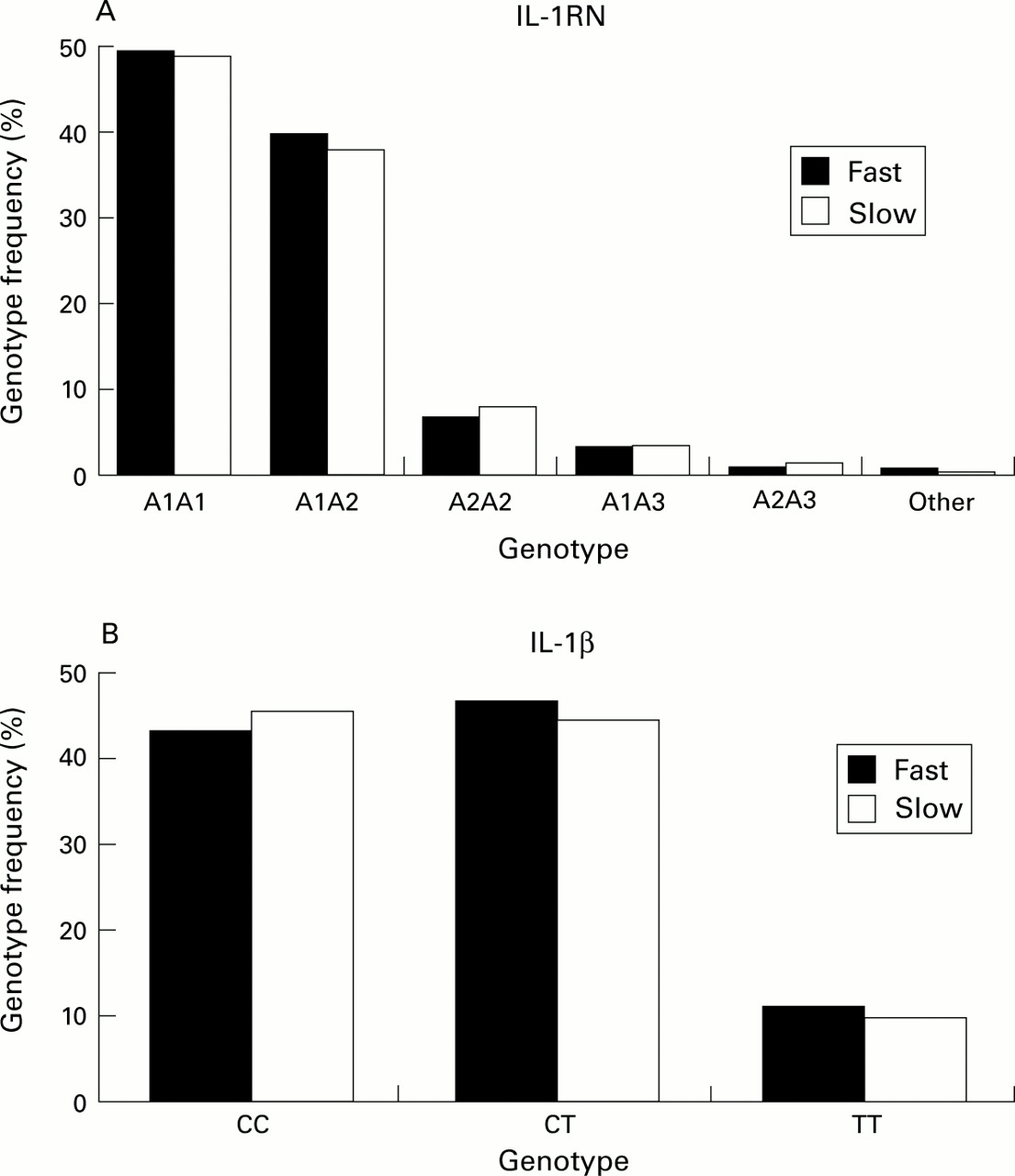
The characteristics of the 590 smokers are given in table 1. Figure 1A shows the distribution of IL1RNgenotypes within the groups of smokers who had a rapid decline and those with no decline in lung function. The observed genotype frequencies were similar to those previously reported.12Figure 1B shows the distribution of IL1Bgenotypes in the smokers who had a rapid decline and those who had no decline in lung function. For both polymorphisms the overall observed distribution of homozygotes and heterozygotes conformed to expectations based on Hardy-Weinberg analysis.
Characteristics of 590 participants in the Lung Health Study group divided into those with no decline and those with a rapid decline in lung function

Frequency of IL1RN and IL1B genotypes in smokers with (n=284, black bars) and without (n=306, white bars) a decline in lung function over 5 years. The differences in genotype frequencies were not statistically significant.
Carriers of the IL1B –511T or theIL1RN*2 allele were not more frequent in the group who had a rapid decline in lung function, even after adjusting for the significantly different baseline values (age, sex, pack-years, baseline FEV1 % predicted, response to methacholine). However, there was a significant difference inIL1B-IL1RN haplotype distribution between those with a rapid decline in lung function and those in whom there was no decline (table 2). Of the individual haplotypes,IL1RN A1/IL1B –511T was increased in those whose lung function declined rapidly (odds ratio (OR) 1.5, 95% CI 1.1 to 2.1, p=0.01) and IL1RN A2/IL1B –511T was significantly decreased in this group (OR 0.7, 95% CI 0.5 to 0.9, p=0.02). As in previous studies,26 linkage disequilibrium was found between IL1RN VNTR and theIL1B polymorphism (table3).
Frequency of IL1RN/IL1B haplotypes in the study groups estimated using the expectation-maximisation (EM) algorithm25
Linkage disequilibrium between the IL1RN and the IL1B polymorphisms
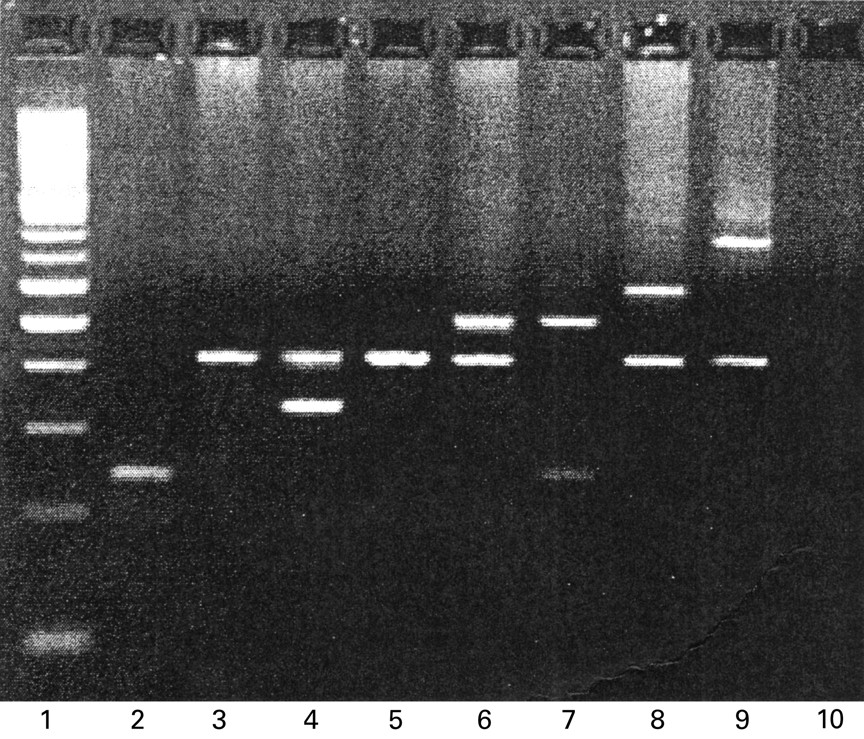
Five alleles of the IL1RN gene have been previously described.12 We found a sixth allele that consisted of eight repeats of the 86 bp sequence (fig2).

{kind=link}
{kind=link}
Two per cent agarose gel displaying IL1RN genotypes. Lane 1: 100 bp DNA ladder; lane 2: allele 2 homozygote (240 bp, 2 repeats); lanes 3 and 5: allele 1 homozygote (412 bp, 4 repeats); lane 4: allele 4 heterozygote (326 bp, 3 repeats); lanes 6 and 7: allele 3 heterozygote (498 bp, 5 repeats); lane 8: allele 5 heterozygote (584 bp, 6 repeats); lane 9: allele 6 heterozygote (756 bp, 8 repeats); lane 10: negative control.
Discussion
We have investigated polymorphisms in the IL-1 gene cluster as risk factors for accelerated decline in lung function. Genotype frequencies were compared in smokers with a rapid decline in lung function and those with no decline in lung function. There was a strong association betweenIL1RN/IL1Bhaplotypes and decline in lung function. Allele 1 of theIL1RN gene in combination with theIL1B T allele was a risk factor for an accelerated rate of decline in lung function. In contrast, allele 2 of the IL1RN gene in combination with theIL1B T allele was protective against a rapid decline in lung function.
IL1B and IL1RNare logical candidate genes in inflammatory airway diseases because of the critical role of IL-1 in inflammation. The actions of IL-1 include activation of T and B cells, and chemotaxis of neutrophils and macrophages. In addition, IL-1 stimulates the production of other cytokines such as tumour necrosis factor (TNF) and granulocyte-macrophage colony stimulating factor (GM-CSF).1 Airway inflammation is a common feature in the development of both COPD and asthma. Previous studies have shown an association between IL1RN*2 and several inflammatory and autoimmune diseases such as ulcerative colitis, systemic lupus erythematosus, diabetic nephropathy, and alopecia areata.13-17 Another polymorphism in theIL1RN gene, which is in complete linkage disequilibrium with IL1RN*2, has been associated with fibrosing alveolitis.27 TheIL1B C–511T polymorphism has been associated with several diseases including inflammatory bowel disease,18 alcoholic cirrhosis,28 and Parkinson's disease.29
The functional effect of these polymorphisms remains unclear. Data on the effect of IL1RN*2 on plasma levels of IL-1RN are contradictory.30 ,31 Hurmeet al investigated the effect ofIL1RN*2 in combination with theIL1B C–511T polymorphism and found thatIL1RN*2 was associated with higher plasma levels of IL-1RN than IL1RN*1, but only in individuals who also had the T allele ofIL1B. 19 These data would appear to conflict with the reported association of theIL1RN*2 allele with inflammatory diseases since increased production of IL-1RN would be expected to reduce the binding of IL-1 to its receptor and therefore have an anti-inflammatory effect. However, mononuclear cells from carriers of theIL1RN*2 allele were shown to produce a higher level of IL-1β in vitro.32 It is possible thatIL1RN*2 predominantly increases the level of IL-1β and therefore leads to an imbalance in the IL-1β/IL-1RN ratio resulting in an increased susceptibility or more severe outcome of inflammatory diseases. Alternatively, it could be hypothesised that theIL1RN*2 allele is a marker for a linked disease associated locus and may not be a direct disease causing allele.
We found no association of either the IL-1RN or IL-1β genotypes with the rate of decline in lung function. However, the association of haplotypes strongly suggests that the combination of risk alleles contributes to the susceptibility to the deleterious effects of cigarette smoke on lung function. This is consistent with the in vitro data of Hurme et al.19 IL1RN*2 in the presence of theIL1B T allele was associated with higher levels of IL-1RN in vitro and with a normal rate of decline in lung function in this study. Conversely, IL1RN*1in combination with the IL1B T allele was associated with lower levels of IL-1RN in vitro and with an accelerated rate of decline in lung function. Individuals with lower levels of IL-1RN may therefore be less protected against the pro-inflammatory effects of IL-1 and thus may be more susceptible to cigarette smoke induced lung injury.
In summary, although the individual polymorphisms in the IL-1 genes were not associated with rate of decline in lung function in smokers, we were able to demonstrate a significant influence ofIL1RN/IL1B haplotypes in these individuals. As in previous functional studies, this suggests that it is the combination of alleles which is important in the regulation ofIL1RN gene expression.
Acknowledgments
This study was supported by a grant from the Canadian Institutes of Health Research. The Lung Health Study was supported by contract N01-HR-46002 from the Division of Lung Diseases of the National Heart, Lung, and Blood Institute. Dr L Joos was supported by the Swiss National Science Foundation, Novartis Research Foundation, Uarda-Frutiger Foundation and the Swiss Society of Pneumology. Dr Sandford was supported by a Parker B Francis Fellowship.