Article Text

Statistics from Altmetric.com
Among constitutively present cells, bronchoactive leukotrienes are produced predominantly by mast cells1 and macrophages.2 A unique characteristic of asthmatic inflammation is the migration of leukocytes from the peripheral blood to the conducting airways of the lung. This is especially true of the eosinophil, which is not present in the airways of normal individuals, but may be found in massive numbers during periods of airway hyperresponsiveness in asthmatic individuals (fig1).3-5 Most experimental models indicate that eosinophils are an invariable component of asthmatic hyperresponsiveness, although some studies suggest that bronchoconstriction can occur in the relative absence of these cells.
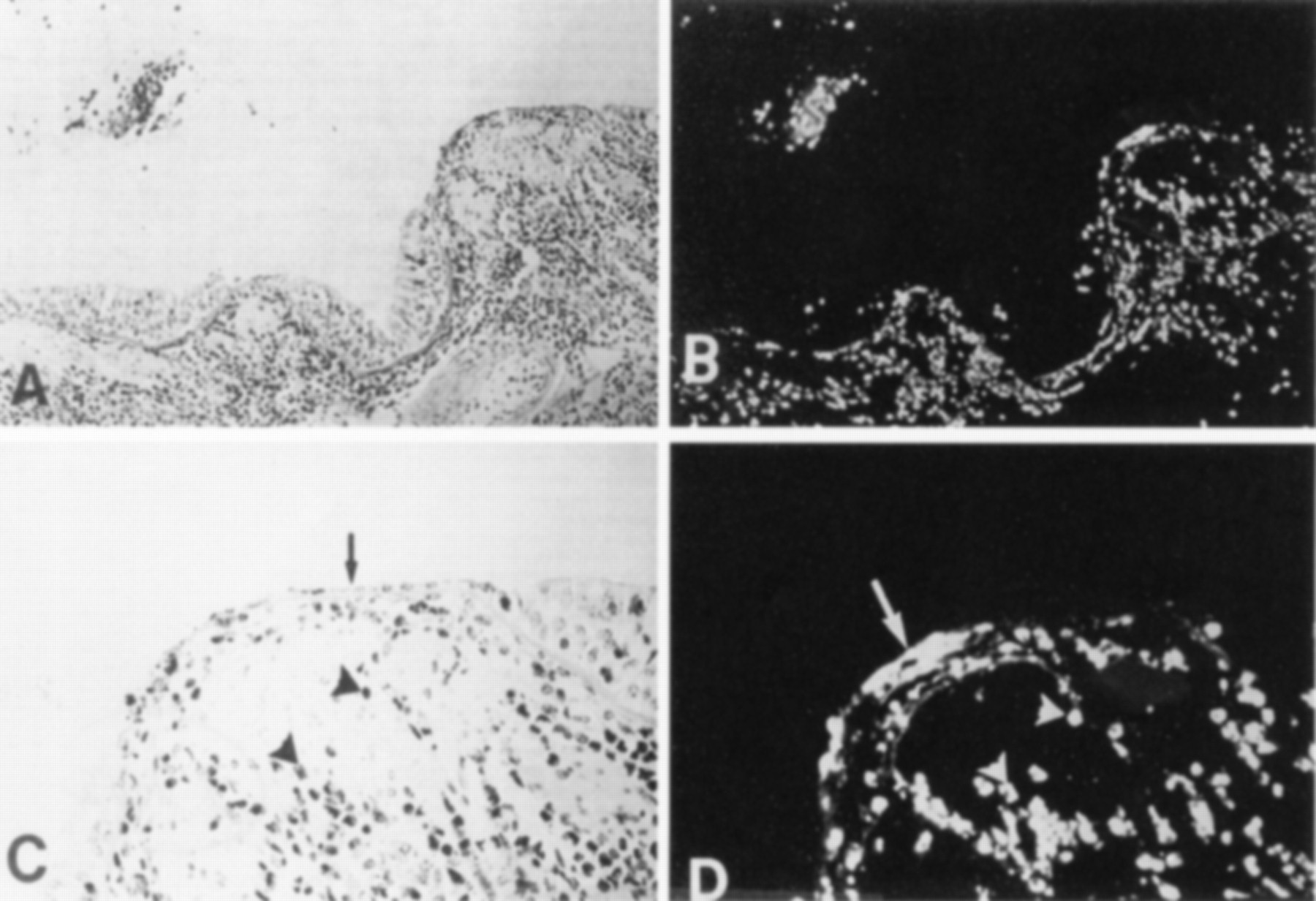
Effect of eosinophil infiltration into human airways. At later stages a cytotoxic effect probably accounts for the epithelial denudation found in human asthma. (A) Histological section of the airway of a patient with asthma (stain: haematoxylin-eosin; original magnification ×160). (B) Same section stained with fluorescent monoclonal antibody for the eosinophil major basic protein (MBP; magnification ×160). (C) Higher power view of the desquamated epithelium in (A) (basal magnification ×400). (D) Localisation of MBP in cells (arrowheads); these cells correspond directly to the cells marked in (C). MBP is also localised outside cells in association with epithelial desquamation (arrow; original transformation ×400). (C) and (D) illustrate the identical area; this section was first stained for MBP by immunofluorescence and subsequently stained with haematoxylin-eosin. Reprinted with permission from Gleich.5
For the purposes of this discussion, the eosinophil will be viewed as a leukotriene transport system capable of providing a substantial reservoir of bronchoactive leukotrienes to airways in relatively short time. They may be delivered to conducting airways in large numbers and have the capacity to produce cysteinyl leukotrienes (LTC4) which cannot be produced by neutrophils, but also migrate into conducting airways in some circumstances.6 ,7 This discussion deals first with the mechanism by which eosinophils, a minority constituent of the circulating blood, are honed selectively to the conducting airways of the lung in human asthma. The mechanism by which the recruitment process of cellular adhesion primes migrating eosinophils for augmented leukotriene synthesis is discussed with particular reference to interactions between surface integrins on eosinophils and their counterligands, the immunoglobulin supergenes on the endothelial surface,8 ,9 and fibronectin within the airway wall.10 The potential physiological significance of augmented secretion caused by adhesive interactions is suggested by experiments elucidated in this discussion. Finally, some new preliminary evidence for an autocrine mechanism of eosinophil recruitment that is mediated through the secretion of cysteinyl leukotrienes is considered.
Mechanisms of eosinophil recruitment
Eosinophils do not reside in human airways under normal circumstances. Normally, these cells are important in combating parasitic infections. While eosinophil morphology has certain similarities to other granulocytes such as neutrophils and basophils, this appearance is elusive. Unlike neutrophils, eosinophils have no phagocytic function. The granular proteins contained in eosinophils are unique to these granulocytes and differ substantially from those of other cells. Unlike neutrophils, eosinophils are a major defence against parasitic infections where they attach along the length of helminths and secrete their granular proteins, which are potent RNAses, to cause death of the infesting organism. Eosinophils are also capable of synthesising cysteinyl leukotrienes as well as LTB4. Neutrophils, by contrast, lack the synthase to convert LTA4into LTC4, and hence are incapable of producing bronchoactive leukotrienes even if recruited to conducting airways. Both neutrophils and eosinophils are capable of synthesising bronchoactive prostaglandins, but these do not appear to have pathogenetic significance in human asthma. Leukotriene B4 has substantial chemoattractant activity for neutrophils11 ,12 and for guinea pig eosinophils13 but has substantially less chemoattractant activity for human eosinophils.14
Eosinophils share surface ligands with other myelopoietic elements. The sequence and mechanism of activation thus determine which cells will be recruited to the airways. Recruitment occurs at the capillary level where flow is slow enough to overcome shear forces that exist at greater perfusing pressures. Cell rolling, presumably mediated by selectins on both the endothelial and eosinophil surface, is the first phase of recruitment and this slows laminar flow further.15 Interleukin-5 has been found in high concentrations in the conducting airways of the lung during periods of asthmatic activity15 ,16 and this cytokine selectively causes shedding of l-selectin from the eosinophil surface and the simultaneous upregulation of β2-integrins on the eosinophil surface. The nature of the process that causes relatively quiescent asthma to enter a phase of eosinophil recruitment is not defined. While it is presumed that the initial secretion of IL-5 is Th2 cell mediated, the trigger for this event is also unknown. Once activated, eosinophils further produce IL-5, and this suggests a possible autocrine amplification of the recruitment process.
The next phase of this recruitment process is tight adhesion of eosinophil integrins to surface ligands on the endothelium. β1-integrin (VLA-4) is constitutively expressed on the eosinophil surface; whether its conformation is changed to augment adhesion is unknown. β2-integrin expression is quantitatively upregulated by a mechanism directly related to binding at the IL-5 receptor (fig 2). Preliminary data (unpublished) suggest that the conformation of MAC-1, a β2-integrin, is also altered toward a higher affinity state by IL-5. As noted above, upregulation of these integrins by IL-5 does not occur for the neutrophil, which lacks the IL-5 receptor, and this may account for the selective migration of eosinophils, which share β2-integrins, MAC-1 and LFA-1 with neutrophils.

Adhesion of a granulocyte to the endothelial wall. Lower: “Rolling” function caused by selectin molecules is followed by tight adhesion to ICAM-1 by β2- and β1-integrin (not shown). Lower right: This leads to diapedesis through the endothelial cell wall into the airway parenchyma. Reprinted with permission from Wegner and Hamann.10
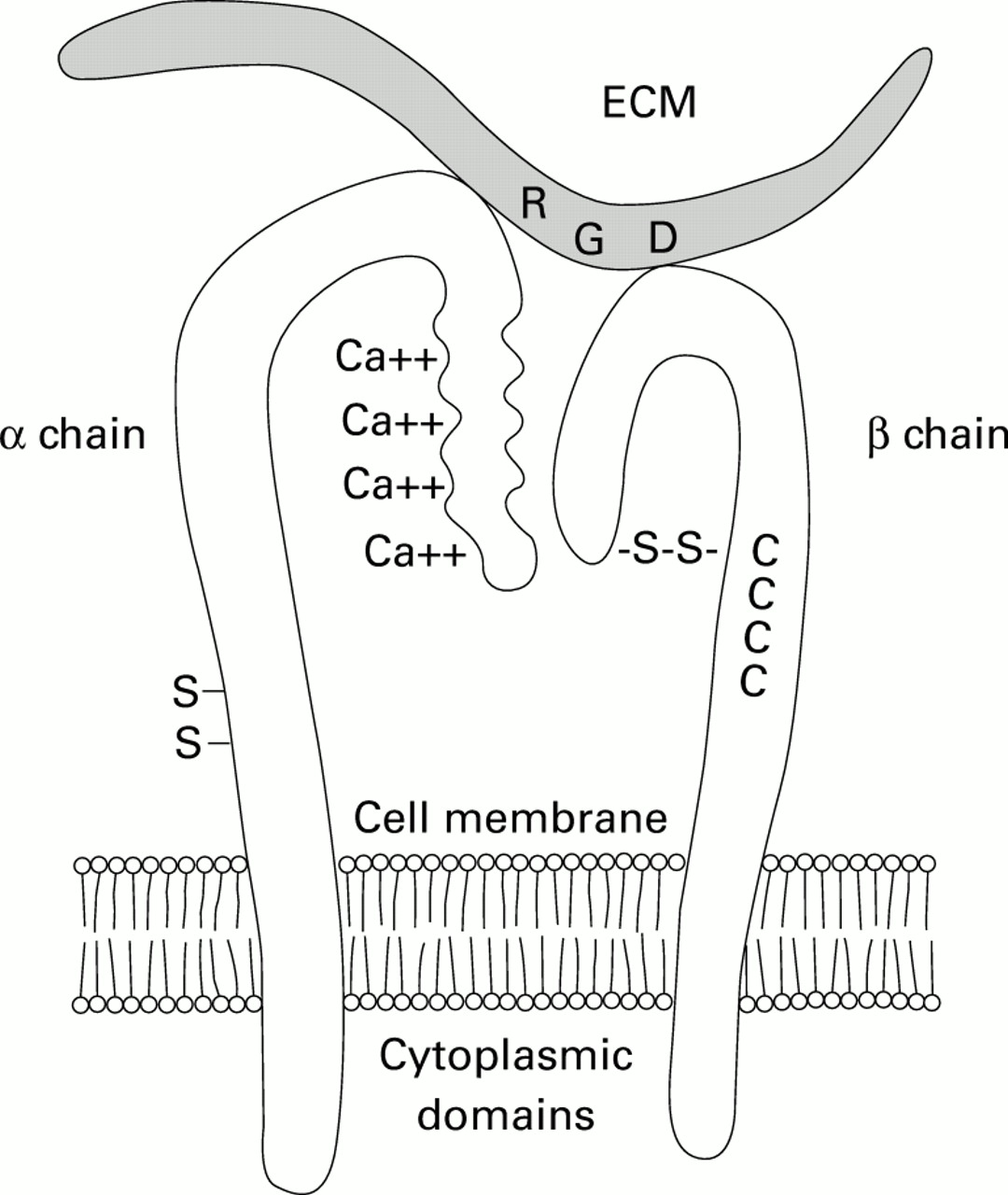
The counterligands for eosinophil surface integrins are the immunoglobulin (Ig) supergenes ICAM-1 (β2) and VCAM-1 (β1) which are endothelial surface molecules containing a 5-domain transmembrane component. The specificity for binding between integrins and endothelial surface ligands is determined by the pocket formed by the α and β chains of the surface integrin (fig 3) into which the ICAM-1 or VCAM-1 molecule inserts. As for eosinophil surface integrins, Ig supergenes are upregulated by specific cytokines including IL-1 and IL-4.17 ,18 The “trigger” for this process remains similarly unknown. It is not known whether β1- or β2-integrins further participate in the selectivity of eosinophil recruitment. β1-integrin is present only on the surface of eosinophils and lymphocytes, but not neutrophils. On the other hand, blockade of β2-integrin is sufficient to prevent eosinophil migration both in vitro and in vivo (see below).

Schematic characterisation of the ligation between integrins on the eosinophil surface (left side) and immunoglobulin supergenes (e.g. ICAM-1, VCAM-1) on the endothelial surface. Specificity of binding is determined by the conformation of the α and β chains into which the terminal portion of the 5-domain transmembranous portion of the immunoglobulin supergene fits.
Relationship between eosinophil recruitment and augmented synthesis of leukotrienes
Another unique property of eosinophils is their ability to ligate to matrix protein within the airway wall. VLA-4, which is not present on neutrophils, binds specifically to the RGD region of fibronectin (FN) (fig 4), and this binding also does not occur for neutrophils. Several investigations have examined the relationship between binding to FN in vitro and the stimulated synthesis of LTC4 using isolated human eosinophils incubated with FN coated microwell plates. The binding process is remarkably slow (60 min; fig 5) compared with binding to cultured human umbilical vein endothelial cells (about 5 min), suggesting that the molecule(s) may need to change conformation in the process. However, once binding has occurred there is a substantial augmentation of stimulated eosinophil secretion of LTC4. Figure 6 indicates that overall secretion increases by about 40%; however, given that only about 20% of the cells are actually bound in these studies, augmentation of LTC4secretion in bound cells may be estimated to be about fivefold. There are preliminary data to suggest that ligation of both β1- and β2-integrin to the endothelial surface also augments eosinophil secretion of LTC4.19 Thus, the process of cellular transmigration that accompanies chemoattraction of eosinophils into the airway matrix appears to be the process by which eosinophil synthesis of leukotriene synthesis is primed. The precise mechanism by which adhesion ligation causes this priming is unknown but is currently under active investigation.
Schematic characterisation of the ligation between VLA-4 with its characteristic α4β1 chains and the RGD region on fibronectin which causes intramural binding of eosinophils following diapedesis. Adapted from Wegner and Hamann.10

Time course of binding of eosinophils to fibronectin (FN) coated microwell plates. Significant binding requires incubation for 60 min, which is substantially longer than for binding to endothelial ICAM-1 by the same eosinophil surface ligand (see text). Reprinted with permission from Neeley et al.20

Augmented secretion caused by priming due to ligation of eosinophil VLA-4 and fibronectin. EPO = eosinophil peroxidase. Reprinted with permission from Neeley et al.20
Physiological consequences of augmented secretion
The notion that eosinophils could exist as innocent bystanders in the process of asthmatic hyperresponsiveness has been addressed in several experimental situations. The mere presence of these cells, even if the association were invariable, does not implicate their physiological significance in the bronchoconstrictor process. As noted above, it would appear that in some circumstances eosinophil infiltration is not essential to produce a substantial bronchoconstrictor response—for example, exercise induced bronchoconstriction. A question to be resolved is whether the transmigration of eosinophils into airways and the concurrent priming of leukotriene secretion is at least capable of causing constriction of airways. If quantities of leukotriene secreted are insufficient, or if blockade of eosinophil infiltration has no effect on airway responsiveness, the eosinophil might well be an innocent bystander.
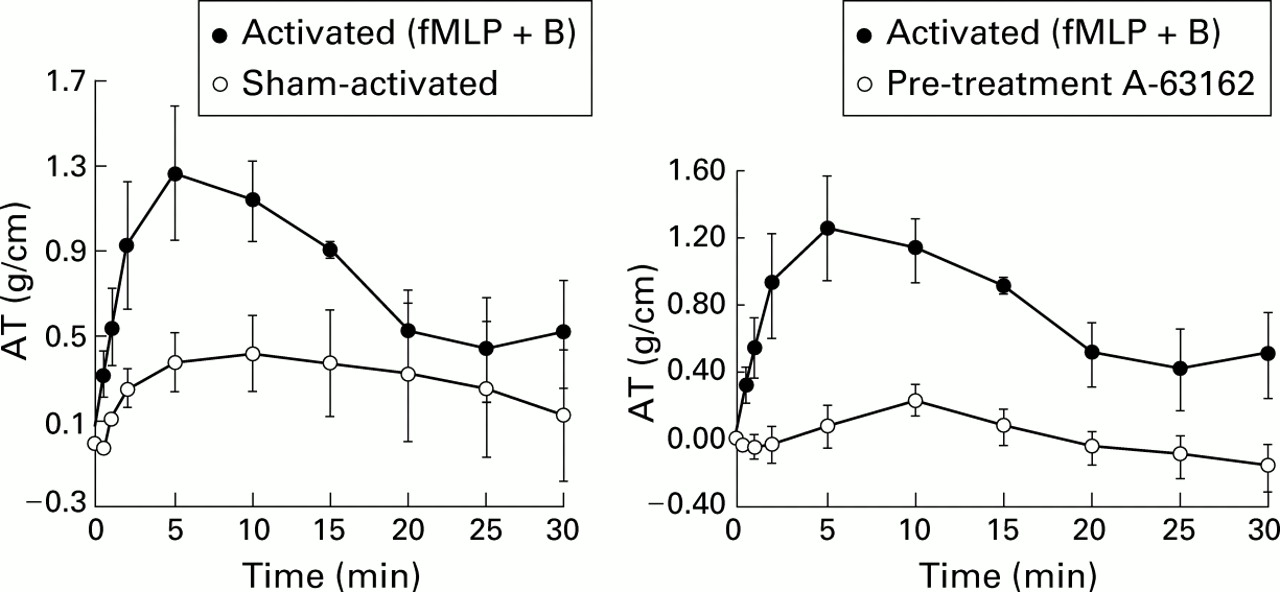
Prior investigation in monkeys sensitised withAscaris suum antigen have shown that eosinophils migrate into sensitised airways.21 Blockade of this migration with anti-ICAM-1 antibody both reduced the number of infiltrating eosinophils and substantially reduced the airway hyperresponsiveness on bronchial challenge. This would suggest that, under these experimental circumstances, eosinophils are essential to airway hyperreactivity (fig 7). In guinea pig tracheal preparations, eosinophils isolated from either normal donors or cultured from human umbilical vein cord blood elements cause substantial contraction (fig8) that is blocked completely with inhibitors of 5-lipoxygenase, the enzyme converting arachidonic acid into LTA4 which precedes production of LTC4. By contrast, comparable activation of isolated human neutrophils has no effect on guinea pig tracheal contraction (fig9).

Relationship between inhibition of eosinophil migration caused by ICAM-1 and airway hyperresponsiveness in monkeys sensitised with Ascaris suum antigen. Blockade of eosinophil migration is associated with return of airway hyperresponsiveness to near normal levels. Reprinted with permission from Leff et al.15
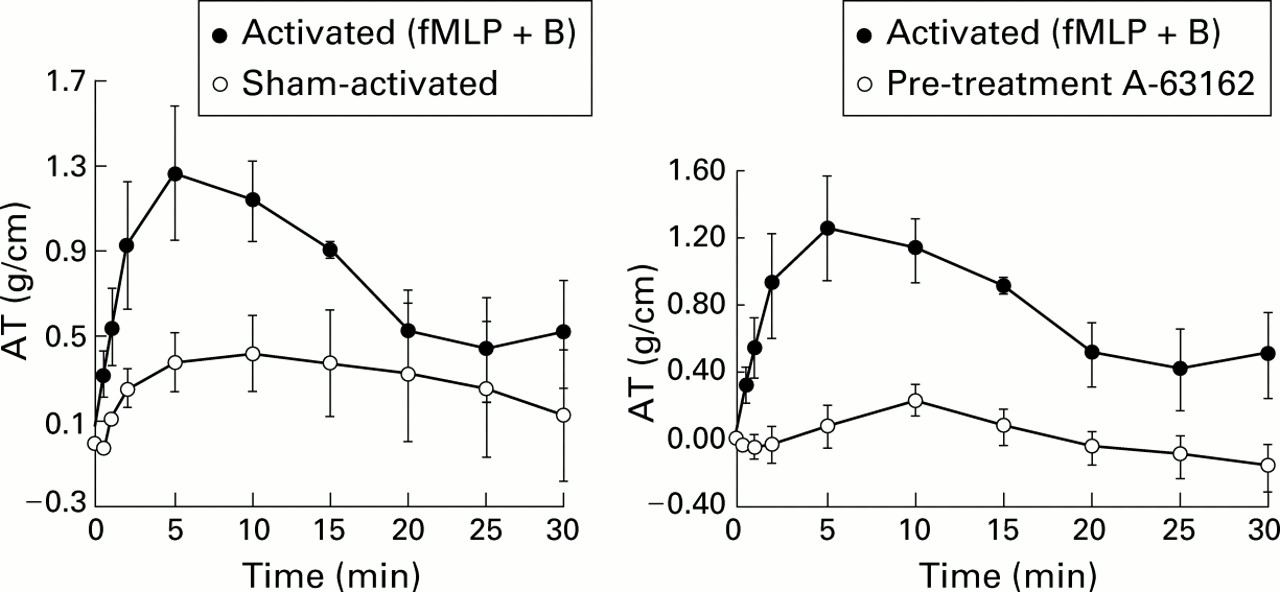
Effect of activated eosinophils cultured from human cord blood on isometric tone in guinea pig trachealis muscle in vivo. Human cells were activated first with fMLP + cytochalasin B and then applied directly to the trachea. Force is expressed as isometric tension per unit area of the tracheal surface. AT = active tension. Reprinted with permission from Hamann et al.22

Effect of human neutrophils activated as in fig 8 on isometric tone in guinea pig trachealis. There is no contractile effect caused by the activating substance fMLP given alone or by fMLP activated neutrophil (compared with fig 8 for eosinophils). Neutrophils lack the enzyme to produce cysteinyl leukotrienes. Positive controls are HL-60 cells, which produce bronchoconstriction prostaglandins, but not leukotrienes. AT = active tension. Reprinted with permission from Munoz et al.23
To test further whether activated human eosinophils could cause contraction of human airway smooth muscle, our laboratory developed a system for videomicrometry of small sections of human airways. Fifth to seventh generation airways can be incubated in 96 well microplates. The airways are photographed through an overhead microscope and the images are stored in real time on a computer. Changes in airway diameter are measured by determining lumenal pixel fitting in a manner described by Schmidt and Rabe.24 Exposure to progressively larger numbers of human eosinophils causes progressive contraction as measured by lumenal narrowing.25 ,26 This narrowing was blocked completely by pretreatment with the 5-lipoxygenase inhibitor A63162, a cogener of the drug zileuton which is now marketed in the United States for the treatment of asthma.
To determine whether the degree of augmentation of secretion of leukotriene caused by adhesion of isolated human eosinophils to FN also caused augmented contraction of human airways, the effect on airway narrowing in human airway sections using isolated human eosinophils was examined. Cells exposed to FN caused a twofold greater narrowing of lumenal area than cells exposed to bovine serum albumin (BSA) as a control (fig 10). Contraction was blocked completely for both BSA and FN treated cells activated with formyl-met-leu-phe (fMLP) + cytochalasin B by pretreatment with the 5-lipoxygenase inhibitorA63162.27 Contraction of airways in all studies was elicited with 105 cells. While these data do not predict the in vivo situation, they do indicate that a physiologically meaningful augmentation of human airway contraction is elicited by adhesive ligation. However, given the dramatic nature of the blockade of the contractile effects caused by eosinophils in this system, it is perhaps somewhat disappointing that anti-leukotriene therapies are less efficacious than would be predicted from these models. These data indicate that the process of asthmatic bronchoconstriction is substantially more complicated than that which can be predicted by a single in vitro model in an isolated cell system.

Effect of fibronectin (FN) ligation of human eosinophils on contraction of human bronchial explants in vitro. There is substantial augmentation of lumenal narrowing in fMLP activated human eosinophils first exposed to FN vs controls exposed to bovine serum albumin (BSA). Exposure of eosinophils to FN alone had no contractile effects. Hence, adhesion molecule ligation has a priming rather than a direct stimulatory effect on airway smooth muscle contraction. PAF = platelet activating factor. Reprinted with permission from Munoz et al.27
Leukotrienes in chemotaxis
Leukotrienes have variable chemotactic properties that are highly cell dependent. There is also a strong species dependence which make data from other species unreliable for prediction of the human condition. The guinea pig eosinophil is rich is LTB4receptors and hence is highly chemotactic in a variety of experimental systems.13 However, the human eosinophil (as noted above) is weakly attracted by LTB4, implying a weaker receptor population. By contrast, human neutrophils are strongly chemotactic and this was an initial concern in the development of anti-leukotriene treatments directed against 5-lipoxygenase. The specific concern was whether the concomitant blockade of LTB4—as well as cysteinyl leukotriene synthesis—that results as a consequence of the blockade of 5-lipoxygenase might also prevent neutrophil chemotaxis. The system has proved to be more robust than feared, and there is no increase in significant respiratory infection when a 5-lipoxygenase agent is used compared with a leukotriene receptor antagonist that has no effect on LTB4. Unfortunately, there is also no evidence that eosinophil chemotaxis is selectively diminished by LTB4 inhibition resulting from synthesis inhibitors. Hence, there is no obvious therapeutic benefit to drugs that block the entire leukotriene synthetic pathway over receptor antagonists that specifically block the cysteinyl leukotriene receptor.
A recently described phenomenon is the ability of the anti-leukotriene agents to block eosinophil migration, at least partially, even with short term use. In guinea pig tracheal explants, the chemotactic agent fMLP causes substantial and selective migration of eosinophils that reside naturally (and quiescently) in the lamina propria (fig 11). Administration of zileuton in concentrations of >10–8 M caused substantial inhibition of eosinophil migration, and full blockade was achieved with higher concentrations.13Because LTB4 receptors are significantly involved in the chemotactic process, it was postulated that the action of zileuton resulted from the unique ability of this anti-leukotriene to block the synthesis of LTB4 as well as the SP1 analogues. The selective LTB4 antagonist LTB4 dimethylamide caused even more potent blockade of eosinophil migration in this model (fig 12). However, administration of the highly selective LTD4 receptor antagonist zafirlukast caused equipotent blockade. Significant inhibition of eosinophil chemotaxis was observed at 10–12 M zafirlukast. Comparable trials have been conducted in humans. Calhoun et al 29 have reported in a preliminary study that large doses of zafirlukast given over 24 hours caused an approximate 50% decrease in the migration of eosinophils into the airways of challenged asthmatic subjects, and Diamant et al 30 have shown independently comparable findings.

Effect of pretreatment with the 5-lipoxygenase inhibitor zileuton on eosinophil migration from the lamina propria to the lumen in guinea pig tracheal explants. Chemotaxis was initiated by lumenal instillation of fMLP. Reprinted with permission from Munoz et al.28

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of pretreatment with inhibitors of LTB4 and LTD4 on eosinophil chemotaxis using the same preparation as for fig 11. LTB4 causes inhibition of chemotaxis at even lesser concentrations than zileuton, suggesting that there is a chemotactic effect of this cysteinyl leukotriene compound. The mechanisms by which LTD4 causes chemoattraction in this model are undefined but similar findings are suggested from human studies (see text). Reprinted with permission from Munoz et al.28
The mechanism by which LTD4 receptor blockade inhibits eosinophil migration remains elusive. There are no cysteinyl leukotriene receptors on the eosinophil surface in either rodents or humans. Hence, the effect is either non-specific—that is, non-receptor related—or the result of complex interactions within the airway involving other cells and/or tissues. Based on the guinea pig model, the airway itself appears to contain all the elements necessary to inhibit eosinophil migration through blockade of the cysteinyl leukotriene receptor. One recent investigation using the leukotriene receptor antagonist pranlukast has shown that the ability of this compound to block eosinophil migration into the airways of challenged guinea pigs is blocked with the monoclonal antibody TRFK-5 which is directed against IL-5.31 This implies, but does not by any means establish, that blockade of the LTD4 receptor on some element contained within the airway wall could have an inhibitory effect on IL-5 secretion, which is an important component of adhesion model upregulation in eosinophil chemotaxis (see above). However, definitive studies are still required to elucidate the mechanism whereby LTD4 receptor blockade affects eosinophil migration. It also remains to be determined whether the magnitude of inhibition of eosinophil migration in humans is clinically and pathophysiologically significant. If so, anti-leukotriene therapies could properly be viewed as being broadly anti-inflammatory in a manner (if not degree) comparable to that conferred by corticosteroid treatment. The implications of this for subsequent airway remodelling have been discussed elsewhere.
Conclusions
As asthma is viewed as an inflammatory process mediated at least in some part by leukotrienes, eosinophils are the major transport systems for these compounds to the airway smooth muscle where they cause contraction, and to the airway vasculature where they cause oedema. Leukotrienes are synthesised de novo in eosinophils directly from membrane phospholipids after activation by phospholipase A2 (PLA2). The process of selective chemoattraction is a fascinating one, as eosinophils are but a minor component of the circulating granulocytes. Even though eosinophils share common surface ligands with neutrophils, they are capable of selective migration into the airway wall. It is likely that cytokine specific processes regulate this selective migration—for example, IL-5.
It is also of considerable interest that the process of molecular adhesion and transmigration is intimately linked to the priming of eosinophil secretion of leukotrienes. The mechanism by which this occurs is unclear, but appears from some very preliminary studies to be related to the direct phosphorylation of PLA2-IV, which may occur as a consequence of adhesion. Another property of leukotrienes that remains unexplained is the apparent ability of these compounds to cause, by a mechanism yet to be defined, substantial chemotaxis of eosinophils in both animal models and in humans.
While eosinophils are the unique inflammatory cells of asthmatic inflammation, it is still unclear if they are essential for all manifestations of the asthma syndrome. It is further unclear whether leukotriene synthesis alone accounts for the bioactivity of these cells in causing airway narrowing in asthma. Blockade of leukotriene activity in human asthma does not cause improvement in airflow obstruction in a manner comparable to that obtained with corticosteroids or high efficacy β2 adrenoceptor drugs. The invariable presence of eosinophils in human asthma does not itself imply a role for these cells in the pathogenesis of the disease. However, the demonstration that adhesion primed eosinophils are capable of causing massive augmentation of leukotriene secretion and that this secretion is of a magnitude sufficient to cause contraction of human airway explants suggests that eosinophils are the source of leukotrienes in human asthma. Nonetheless, the roles of eosinophils and of leukotrienes in human asthma may vary among the different asthma phenotypes that are only now being defined.
References
Footnotes
Reprinted from the American Journal of Respiratory and Critical Care Medicine2000;161(Suppl):S125–32.