Article Text

Abstract
BACKGROUND Aspergillus fumigatus is a pathogenic fungus which causes a range of diseases, particularly in the human lung. The pathological mechanism is unknown but may involve a complex mixture of biomolecules which can diffuse from the spore surface. This material is known asA fumigatus diffusate (AfD) and has previously been shown to have a range of immunosuppressive functions. It is hypothesised that AfD may influence the binding of spores to extracellular matrix (ECM) proteins and lung epithelial cells, thereby affecting the ability of the fungus to cause infection.
METHODS The binding of spores to ECM proteins and to epithelial cells was carried out using a direct binding assay in microtitre plates and spores were counted by phase contrast microscopy. Rat bronchoalveolar lavage (BAL) fluid was enriched for surfactant protein D (SP-D) using maltose agarose affinity chromatography. The effects of AfD and the SP-D enriched BAL fluid were assessed by pre-incubation with ECM proteins or epithelial cells in the direct binding assay.
RESULTS AfD enhanced the binding of spores to laminin by 137% and to A549 epithelial cells by 250%. SP-D enriched BAL fluid inhibited spore binding to ECM proteins and epithelial cells. Pre-incubation of ECM proteins and epithelial cells with SP-D enriched BAL fluid prevented the enhancement of spore binding by AfD, and pre-incubation of ECM proteins and epithelial cells with AfD prevented the inhibition of spore binding by SP-D enriched BAL fluid. This pretreatment did not prevent the enhancement of spore binding, giving an increase of 95% for collagen I, 80% for fibronectin, 75% for laminin, and 150% for A549 cells.
CONCLUSIONS The hypothesis that AfD would affect spore binding to ECM proteins and epithelial cells was confirmed. Rat BAL fluid, with SP-D as the possible bioactive agent, prevented this enhancement. The in vivo significance is unclear but the enhanced binding of spores may increase the chance of fungal infection in the lung which could be prevented by the protective effects of lung surfactant components (possibly SP-D). The results suggest that there may be competition between AfD and a BAL fluid component (possibly SP-D) for the same or similar binding sites on ECM proteins and epithelial cells. Whether this competition occurs in vivo requires further investigation.
- Aspergillus fumigatus
- surfactant protein D
- bronchoalveolar lavage fluid
- A549 epithelial cells
- extracellular matrix proteins
Statistics from Altmetric.com
- Aspergillus fumigatus
- surfactant protein D
- bronchoalveolar lavage fluid
- A549 epithelial cells
- extracellular matrix proteins
The fungus Aspergillus fumigatus is commonly found in the environment, its usual habitat being dead or decaying organic matter. The fungus causes many human lung diseases1 ,2 and a major factor in determining its pathogenicity is the size of the spores which are about 3 μm in diameter. Mullins et al 3 have shown that many more spores of A fumigatusare present in the lungs at necroscopic examination than would be anticipated from their presence in the air. The survival rate of spores of A fumigatus in rabbit lung is also greater than that of control spores of a similar size.4The spores therefore appear to have some survival advantage in the human lung over other spores. The exact mechanism of pathogenicity is unclear but the adhesion of spores to host tissues and, in particular, to proteins of the extracellular matrix is believed to be an essential event leading to persistence of the fungus in the lung.5-7 Fungal spores also adhere to lung epithelial cells whereby they can be internalised and evade phagocytosis,8 leading to destruction of the host tissue.7 Various defence mechanisms exist to protect against spore persistence such as phagocytosis by the alveolar macrophage and, in particular, the involvement of bioactive components found in lung surfactant. This is a complex mixture of lipids and proteins which is essential for normal lung function by reducing surface tension.9 ,10 Surfactant proteins A (SP-A) and D (SP-D) have been shown to enhance the uptake of bacteria11and A fumigatus spores12 by phagocytic cells. SP-D seems to be particularly important as levels have been reported to increase in patients with lung injury or pathogen infection13 while those of SP-A seem to decrease.14 Several other factors may contribute to the pathogenicity of A fumigatus. These include gliotoxin (an immunosuppressive hyphal toxin),15 proteases which reduce cell spreading16 and promote detachment of cultured cells,17 ,18 and an as yet uncharacterised component from the spore surface known asAspergillus fumigatus diffusate (AfD).19 AfD has several activities including inhibition of the spreading and chemotaxis of macrophages,16inhibition of the macrophage respiratory burst,20 and inhibition of macrophage cytokine gene expression21 and phagocytosis.22 The components of AfD responsible for these activities are as yet unknown, but studies are ongoing in this laboratory to characterise AfD more fully. Since AfD is quickly released from the spore surface in aqueous solution19 and, when inhaled, the spore comes into contact with lung surfactant and its components, it is possible that some of the constituents of AfD may influence the ability of spores to bind to epithelial cells and to proteins of the extracellular matrix. It is also possible that protective components within the surfactant such as SP-D may interfere with spore binding. This study was therefore undertaken to examine the effects of AfD and SP-D enriched BAL fluid on spore binding to ECM proteins and A549 epithelial cells.
Methods
MATERIALS
In-house bred Wistar rats were used throughout. All chemicals were of the highest analytical grade available. Collagen I (calf skin), laminin (from Engelbreth-Holm-Swarm mouse sarcoma), fibronectin (human plasma), and maltose agarose were from Sigma (Poole, Dorset). Fibrinogen (human plasma) was from Calbiochem.
BRONCHOALVEOLAR LAVAGE
Rats were killed by intraperitoneal injection of sodium pentobarbitone (Euthatal; Rhone Merieux Ltd). The lungs were dissected free of the thoracic cavity and were lavaged with 4 × 8 ml sequential volumes of saline at 37°C. The bronchoalveolar lavage (BAL) fluid was concentrated either by freeze drying or by using a centrifugal microconcentrator with a cut off molecular weight of 5 kDa (Microsep, Flowgen). This was aliquoted and stored at –30°C until required for production of SP-D enriched BAL fluid.
FUNGAL SPORES AND A FUMIGATUSDIFFUSATE
A single strain of A fumigatus was isolated from the sputum of a patient with allergic bronchopulmonary aspergillosis. Spores were obtained from cultures grown for 10 days at 30°C on malt agar. Spore suspensions were prepared in 5 mM sodium phosphate buffer pH 7.3 by gentle homogenisation. Spore counts were performed using an improved Neubauer chamber. To obtain diffusate the spores were incubated at a concentration of 108/ml in Hanks' balanced salt solution (HBSS) at 37°C for one hour in an orbital incubator. Spores were removed by passing the suspension through Whatman number 1 filter paper followed by a 0.22 μm sterile filter and the soluble diffusate was stored at –20°C. To prepare spores for the binding assays a spore suspension was prepared which was incubated as above. The suspension was centrifuged at 25 000g for 30 minutes at room temperature, the supernatant was removed, and the spore pellet was resuspended in the same buffer. This was repeated a total of six times and the washed spores were used immediately in the binding assay. Alternatively, they could be stored at –30°C for at least three months without affecting binding ability. To ensure that all the AfD components were removed by repeated washing, samples of the supernatants were routinely subjected to SDS-PAGE to monitor loss of protein. Protein was not usually detectable after three washes.
SPORE BINDING ASSAY
Laminin, fibrinogen, fibronectin, or collagen I in 200 μl phosphate buffered saline (PBS) were coated onto 96-well plates (Helena Biosciences) at concentrations up to 500 μg/ml at 4°C for 16 hours. The plates were washed three times with PBS and were blocked for one hour at 37°C with 1% w/v bovine serum albumin (BSA) in PBS. The plates were washed several times with PBS and 200 μl of spore suspension (106/ml) was added. This was incubated for one hour at 37°C and washed several times with PBS. Glutaraldehyde (200 μl 10% v/v solution) was added for 15 minutes at room temperature and the spores were counted (five fields of view for each triplicate well) using a DM 1L Leica inverted phase contrast microscope at ×400 magnification.
GROWTH OF A549 EPITHELIAL CELLS AND SPORE BINDING ASSAY
Cells were grown in complete Dulbecco's modified Eagle medium (CDMEM) at 37°C in 5% v/v CO2 for 2–3 days. They were harvested by trypsinisation and centrifugation at 3000g and resuspended in CDMEM. Cells were seeded into 96-well culture plates (Corning) at 105cells/well and were grown to confluence overnight. The binding assay is an adaptation of one previously reported.6 Briefly, after washing the plates with PBS, 200 μl of spore suspension (106/ml in CDMEM) was added to each well and incubated as above for 30 minutes with gentle shaking. The plates were then washed with PBS containing 0.05% v/v Tween-20 then fixed with 2.5% v/v glutaraldehyde for 15 minutes and counted as above.
PARTIAL PURIFICATION OF SP-D
Partial purification of SP-D was carried out as previously reported by affinity chromatography using maltose agarose.23 The SP-D was eluted using NaCl which was then dialysed exhaustively at 4°C against PBS. This SP-D enriched fraction (SP-D enriched BAL fluid) was then concentrated using a centrifugal micro-concentrator with a cut off molecular weight of 5 kDa (Microsep, Flowgen), aliquoted, and stored at –30°C. SDS-PAGE of this material showed an intense protein band at around 43 kDa, the known molecular weight of SP-D, together with several other proteins, one of which was around 30 kDa which is characteristic of SP-A.
ENHANCEMENT OF SPORE BINDING BY AFD
This was performed as for the standard spore binding assay above except that AfD (up to 100% v/v) was added to the wells after the blocking step followed by incubation at room temperature for three hours. The plates were washed several times with PBS before incubation with spores. A control used to test the specificity of AfD was a diffusate made from spores of Aspergillus nidulans prepared as given above for AfD.
EFFECT OF SP-D ENRICHED BAL FLUID AND AFD ON SPORE BINDING
This was performed as for the standard spore binding assay above except that up to 10 μg of lavage protein or SP-D enriched fraction in PBS was added to each well after the blocking stage. Plates were then incubated for three hours at 37°C and washed in PBS before incubation with spores. In some experiments (1) up to 5 mM CaCl2 was added before the SP-D enriched BAL fluid, (2) AfD (up to 100% v/v) was added after SP-D enriched BAL fluid, or (3) AfD (up to 100% v/v) was added before SP-D enriched BAL fluid and CaCl2 followed by incubation for three hours at 37°C. When the binding assay involved A549 cells, CaCl2, AfD and SP-D enriched BAL fluid were in CDMEM with all incubations at 37°C in 5% v/v CO2. To test the specificity of the SP-D enriched BAL fluid, controls consisting of the same concentration of BSA in place of this fraction were also included.
SDS-PAGE AND PROTEIN ASSAY
Protein was assayed using the BioRad reagent and SDS-PAGE was performed by the method of Laemlli.24 Proteins were stained using GelcodeTM silver stain (Pierce).
EXPERIMENTAL DESIGN AND STATISTICAL ANALYSIS
All experiments were performed in triplicate at least three times (n ⩾9) to allow for any variability in the quality of the AfD in terms of spore binding enhancement since, in a previous publication,19 we found that the ability of AfD to inhibit the lung macrophage oxidative burst was highly variable. The data were subjected to analysis of variance with the Tukey test for significance of differences between treatments with p<0.05 as the level of significance. In the experimental results shown in fig 4 two way analysis of variance was done using the concentrations of lavage protein and calcium as the two factors, with p<0.05 as the level of significance.

Effect of SP-D enriched BAL fluid on spore adherence to ECM proteins.
Results
SPORE BINDING TO EXTRACELLULAR MATRIX PROTEINS AND A549 EPITHELIAL CELLS
Spores bound to both collagen I and laminin in a concentration dependent manner (fig 1). A similar result was obtained for fibronectin and the plasma glycoprotein fibrinogen although there was some variation in the number of spores bound at each protein concentration when compared with collagen I and laminin (data not shown). The effect of spore concentration on the adherence of spores to a fixed concentration of A549 cells is shown in fig 2 where again a dose-dependent relationship is seen. Using BSA as a control, it was previously shown that this binding was specific.25 It had been reported that incubation of A549 epithelial cells with AfD had an effect on the viability of the cells which was decreased to about 37% (95% CI 32 to 44) after treatment with 100% AfD for 48 hours.25 In all the experiments incubation was for three hours and the cell viability was more than 90% as shown by trypan blue exclusion.
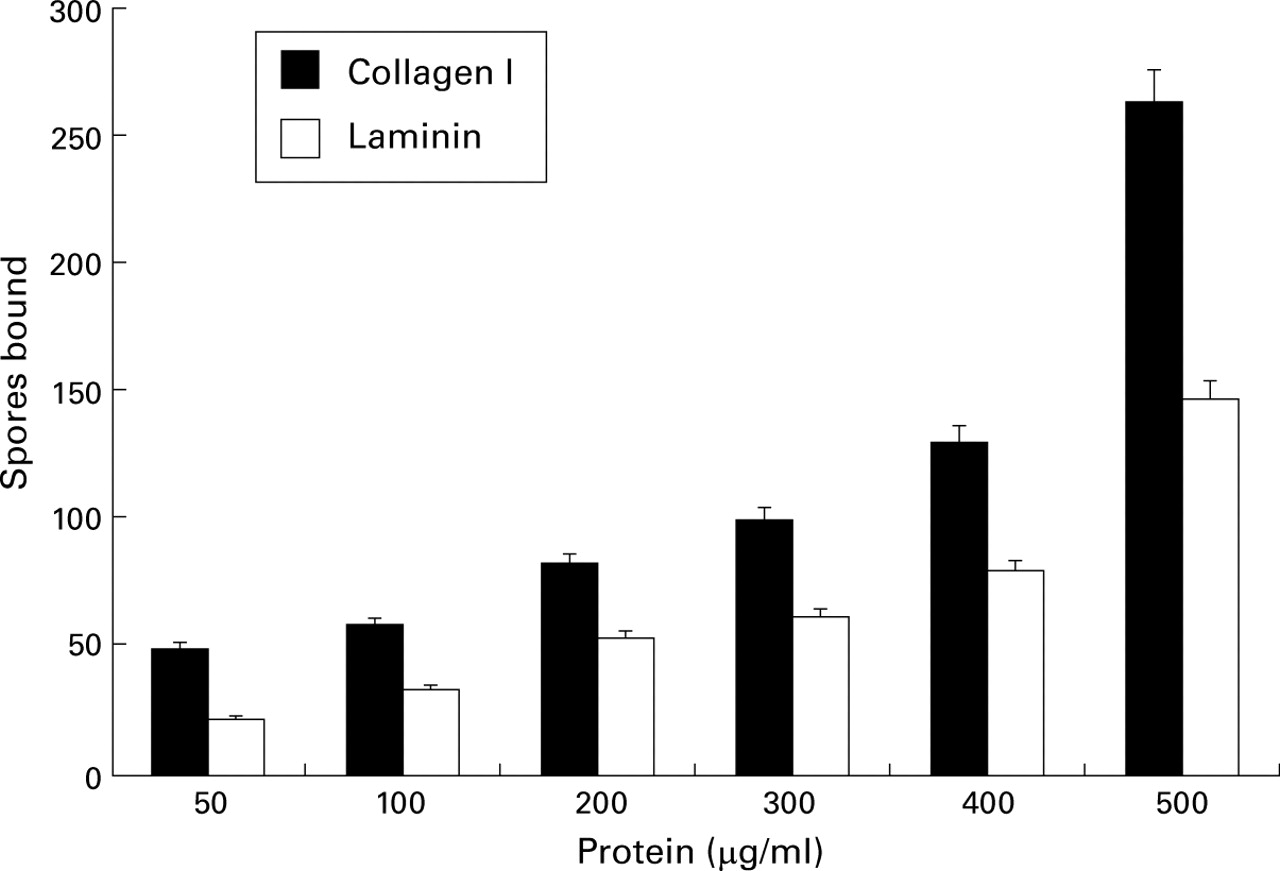
Effect of protein concentration on spore adherence to ECM proteins.

Effect of spore concentration on adherence to A549 epithelial cells.
EFFECT OF AFD ON SPORE BINDING TO EXTRACELLULAR MATRIX PROTEINS AND A549 EPITHELIAL CELLS
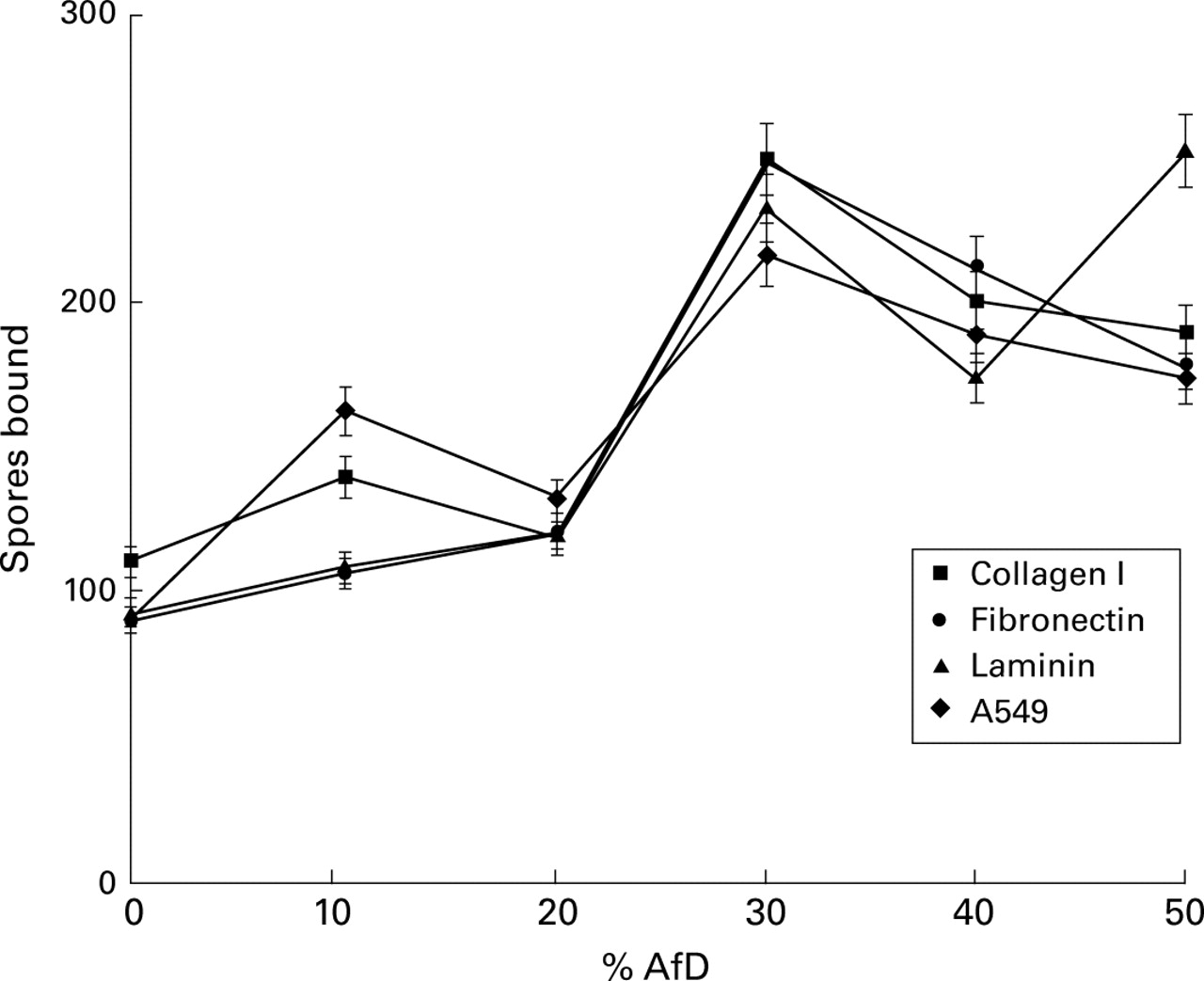
AfD significantly enhanced spore binding to A549 cells at both concentrations used (p<0.002; fig 3). After pre-incubation of the cells with 100% AfD the number of bound spores was enhanced by about 250% (95% CI 261 to 292). A control using a diffusate fromA nidulans did not show significant enhancement of spore binding to A549 cells (data not shown). A significant enhancement was also seen in spore binding to laminin of 137% (95% CI 172 to 221), p<0.005, but not to fibrinogen. A similar enhancement effect of AfD (data not shown) was seen for collagen I (p<0.005) and for fibronectin (p<0.002).

Effect of AfD on spore adherence to ECM proteins and A549 epithelial cells.
EFFECT OF SP-D ENRICHED BAL FLUID ON SPORE BINDING TO EXTRACELLULAR MATRIX PROTEINS AND A549 EPITHELIAL CELLS
Pre-incubation of collagen I, laminin, and fibronectin with SP-D enriched BAL fluid inhibited spore binding compared with the controls (fig 4) and this was enhanced for collagen I and laminin in the presence of calcium but not for fibronectin. Pre-incubation of collagen I with 5 μg SP-D enriched BAL fluid protein in the presence or absence of calcium showed no significant difference from the control. At 10 μg/ml protein there was a significant (p<0.002) difference in spore binding compared with the control and again there was enhanced inhibition in the presence of calcium. Similar results were obtained for laminin except that the inhibition of spore adherence in the presence of calcium was more pronounced. Similar data showing inhibition of spore adherence were obtained for A549 epithelial cells (data not shown) although calcium did not enhance the inhibition. When the SP-D enriched BAL fluid protein was replaced with BSA no significant effect was seen (data not shown).
INFLUENCE OF SP-D ENRICHED BAL FLUID ON THE ENHANCEMENT OF SPORE BINDING TO EXTRACELLULAR MATRIX PROTEINS AND A549 EPITHELIAL CELLS BY AFD
Spore binding following pre-incubation of collagen I with 5 μg and 10 μg SP-D enriched BAL fluid was not significantly different from the control (p>0.05). Only when 10 μg SP-D enriched BAL fluid protein was used in the presence of calcium was there significant inhibition of spore binding (p<0.005; fig 4). When the ECM proteins and A549 cells were pre-incubated with 10 μg SP-D enriched BAL fluid protein in the presence of calcium, AfD failed to enhance spore binding significantly (fig 5). Pre-incubation with SP-D enriched BAL fluid protein appears to prevent the enhancement of spore binding by AfD as shown in fig 3. No significant effect was seen when SP-D enriched BAL fluid protein was replaced with BSA (data not shown).

Effect of AfD on the enhancement of spore binding to ECM proteins and A549 epithelial cells pre-treated with SP-D enriched BAL fluid.
INFLUENCE OF AFD ON SP-D ENRICHED BAL FLUID INHIBITION OF SPORE BINDING TO BASEMENT MEMBRANE PROTEINS AND A549 EPITHELIAL CELLS
When ECM proteins and A549 cells were pre-incubated with AfD, SP-D enriched BAL fluid protein did not prevent enhancement of spore binding (fig 6). Pre-incubation with AfD also seemed to prevent the inhibition of spore binding by SP-D enriched BAL fluid protein since 50% AfD gave a clear enhancement of binding compared with 0% AfD: 90% (95% CI 171 to 208) for collagen I, 80% (95% CI 165 to 192) for fibronectin I, 75% (95% CI 160 to 191) for laminin, and 150% (95% CI 221 to 266) for A549 cells. No significant effect was seen when SP-D enriched BAL fluid protein was replaced with BSA (data not shown).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of SP-D enriched BAL fluid on the inhibition of spore binding to ECM proteins and A549 epithelial cells pre-treated with AfD.
Discussion
It is clear from the data presented here and from the literature5-7 that spores from A fumigatus bind to ECM proteins and to A549 epithelial cells. The effects of the spore surface diffusate AfD have also been widely reported.16 ,19-22 ,25 What has not been investigated previously are the possible effects of components of AfD, which are rapidly released into aqueous solution from the spore surface, on spore adhesion and their persistence in the lung. Clearly, the lung surface is an aqueous environment and it was thought that AfD may influence spore binding and interfere with spore clearance mechanisms. Our results show that AfD enhances spore binding to ECM proteins, fibrinogen, and to A549 epithelial cells. The mechanism of enhancement remains to be elucidated and we are currently attempting to purify the component(s) of AfD involved. This enhancement may provide a mechanism by which the residence time of the spore in the lung is increased and, together with the known immunosuppresive effects of AfD,16 ,19-22 ,25 the chances of germination and infection are thus increased. The in vivo significance of this enhancement needs to be determined as the amount of AfD used in these experiments is far greater than would be encountered in the lung. However, it is only necessary for a single spore to evade clearance to potentially cause infection. This interaction clearly requires further study.
Lung surfactant proteins SP-A and SP-D have an important role to play in lung defence mechanisms and bind directly to A fumigatus spores.12 ,26 Our results show that, when rat lung lavage fluid enriched with SP-D was pre-incubated with ECM proteins, fibrinogen and A549 cells, spore binding was decreased significantly. It is not clear which component of SP-D enriched BAL fluid is responsible for the decrease in spore binding but it has been suggested that it may be SP-D since the inhibition is enhanced by the addition of calcium and it has already been reported that binding of both SP-A and SP-D to conidia was enhanced by calcium.12This fits in well with its known role in enhancement of spore clearance and its general protective role in the lung.
Pre-incubation of ECM proteins and A549 cells with SP-D enriched BAL fluid prevented the enhancement of spore binding by AfD. Furthermore, pre-incubation of ECM proteins and A549 cells with AfD prevented the inhibition of spore binding by SP-D enriched BAL fluid but did not prevent the enhancement of spore binding. This strongly suggests that one or more components in lavage fluid, possibly SP-D, is competing with a component(s) of AfD for the same binding sites on A549 epithelial cells and the ECM proteins. It remains to be seen whether SP-D can, indeed, bind to ECM proteins; we are not aware of any such study in the literature.
The significance of the potential competition between components of AfD and lung surfactant in vivo remains unclear but, in the situation where spores are inhaled, release of AfD from the spore surface could block potential binding sites for SP-D on epithelial cells or ECM proteins thus preventing SP-D from exerting a protective role. Variation in the lung levels of SP-D or other lung surfactant components may also be a factor in determining the clearance of spores. Lower levels of SP-D than normal may enhance the residence time of spores in the lung. Although the authors are not aware of reports of decreased levels of SP-D, it has been reported that SP-A levels can decrease14and SP-D levels increase13 in patients with lung disease.
The data presented here suggest a complicated interaction and potential competition between components of AfD and lung surfactant, the outcome of which may or may not result in A fumigatus infection. Research will continue on two fronts—to fractionate the AfD to identify the enhancer of spore binding and to study the interaction of purified SP-D with ECM proteins and A549 cells.
References
Footnotes
This work was funded by the School of Life Sciences, Napier University with a postgraduate research studentship to ZY.