Article Text

Abstract
BACKGROUND Lymphangioleiomyomatosis (LAM) is an uncommon lung disease for which no effective method of treatment has been found. The predilection of LAM for premenopausal women has led to the assumption that hormonal factors play an important role in the pathogenesis of this disease. The aim of this study was to determine if women with LAM manifest alterations in the catechol-O-methyltransferase (COMT) pathway which is essential for preventing the generation of oestrogen derived reactive oxygen species (ROS).
METHODS Blood samples were collected from 15 women with LAM and compared with appropriate controls. The distribution of high and low activity alleles of COMT was determined with a PCR based RFLP assay. The enzymatic activity of COMT was measured in each sample and the potential presence of a circulating inhibitor of COMT was determined. Since an alteration in the COMT pathway could increase the oxidative stress, the plasma concentration of malondialdehyde (MDA), a secondary product generated from lipid peroxidation, has been used as an internal marker.
RESULTS The distribution of high and low activity alleles of COMT (namedCOMT HH,COMT LL, andCOMT HL) was similar in the two groups with proportions of 40%, 7%, and 53%, respectively, in the women with LAM and 38%, 6%, and 56% in the control subjects. The mean (SD) COMT activity was 24.2 (12.3) pmol/min/mg protein in women with LAM and 24.1 (6.3) pmol/min/mg protein in the control group. Incubation of plasma from women in the two groups with a preparation of commercial COMT showed that no detectable COMT inhibitor was present. The plasma concentration of MDA in the women with LAM was also not significantly different from control subjects.
CONCLUSIONS This study shows that there are no significant alterations in the COMT pathway of women with LAM. It is therefore unlikely that alterations in oestrogen mediated cell signalling pathways are mediated by oxidants derived from an excess of catecholoestrogens in LAM.
- lymphangioleiomyomatosis
- oestrogen metabolism
- catechol-O-methyltransferase
Statistics from Altmetric.com
Lymphangioleiomyomatosis (LAM) is an uncommon lung disease with an incidence of less than 1/100 000 per year which affects almost exclusively women in their childbearing years.1 Although much information has been accrued about the pathological and clinical manifestations of LAM, the molecular and cellular mechanisms responsible for the disease have remained a mystery and an effective treatment has yet to be defined. The predilection of LAM for premenopausal women has led to the assumption that hormonal factors play an important role in the pathogenesis of the disease.2 At least three different lung transplantation groups have reported that LAM can recur in the transplanted lung.3-5 These reports point to a circulating factor, possibly related to oestrogen metabolism, that could play an important part.
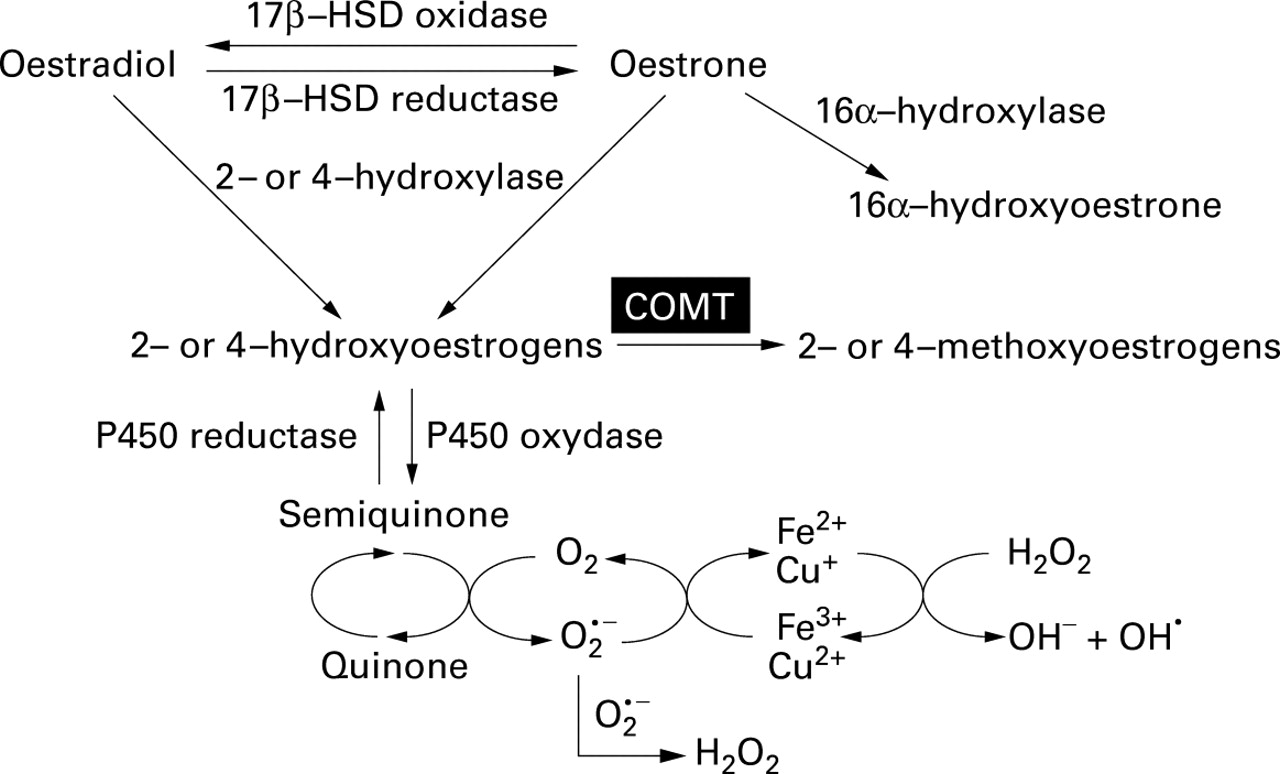
Oestradiol is converted to oestrone by the 17β-hydroxysteroid dehydrogenase (17β-HSD) oxidase and vice versa by the 17β-HSD reductase. Both oestradiol and oestrone can efficiently be hydroxylated by the 2- and 4hydroxylation pathways. These 2- or 4hydroxyoestrogens, also called catecholoestrogens, can be oxidised to form semiquinones and quinones. During this latter conversion oxygen is reduced, producing the superoxide anion which can generate hydroxyl radical in the presence of iron or copper (fig 1).6Hydroxyoestrogens derived from either oestradiol or oestrone show the same capacity to generate reactive oxygen species (ROS), as determined by induction of oxidative damage to DNA.7 At low concentration these ROS can behave as second messengers activating transcription factors such as NF-κB8 and, interestingly, mediate mesenchymal cell proliferation.9 In light of these studies, ROS continuously produced at low levels by hydroxyoestrogens may contribute to the induction of smooth muscle cell proliferation associated with LAM. Thus, alterations in oestrogen metabolism in LAM may conceivably involve an increase in catecholoestrogen production or a decrease in the expression of enzymes involved in their elimination.

Catecholoestrogen metabolism. 2- and 4-hydroxyoestrone are catecholoestrogens which, when oxidised, form semiquinones and quinones. This last redox cycle generates reactive oxygen species (ROS) in the presence of oxygen. Semiquinone/quinone synthesis is prevented by catechol-O-methyltransferase (COMT). 17β-HSD = 17β-hydroxysteroid dehydrogenase.
Accumulation of these hydroxyoestrogens is not important in healthy women. However, in diseases such as breast cancer, local alterations in oestrogen metabolism have been detected which favour the production of catecholoestrogens,10 and these oestradiol metabolites are suggested to be related to breast cancer progression.11
Prevention of ROS generation by catecholoestrogens is closely related to the activity of the catechol-O-methyltransferase (COMT). This enzyme catalyses the methylation of hydroxylated sites on the aromatic ring of catechol compounds which prevents their conversion to semiquinones and quinones, and therefore blocks the generation of ROS.6 While kidney and liver cells are major sources of COMT, red blood cells and mononuclear cells are also rich in COMT and can convert significant amounts of catecholoestrogens to their inoffensive monomethoxy derivatives. Red blood cell COMT has been shown to play an important part in the detoxification of oestrogen catechols, as low red blood cell levels of COMT are thought to contribute to oestrogen carcinogenicity in hamster kidneys.12Interestingly, kidney carcinogenesis is observed with increased frequency in patients with tuberous sclerosis, a disease in which the incidence of LAM is also increased.13
Another possible correlation of COMT with LAM is abnormal HMB-45 antigen positive smooth muscle cell proliferation along the lymphatics, the key histopathological feature observed in the lungs of patients with LAM.14 The HMB-45 reactive antigen has been well characterised and studies have shown that it has full homology with the protein encoded by cDNA of Pmel 17, a protein thought to play a critical role in the conversion of indole-5,6-quinone and indole-5,6-quinone-2-carboxylic acid to eumelanin (fig2).1 ,15 Eumelanin, a black-brown pigment, is one of two major classes of cutaneous melanin, the other being the yellow-red pheomelanin.15 ,16 Since the precursors of eumelanin are quinones with the potential to generate superoxide and hydroxyl radicals and since HMB-45 reactive antigen is homologous to the Pmel gene product, this raises the possibility that quinone derived oxidants may be directly involved in the pathophysiology of LAM.

Role of catechol-O-methyltransferase (COMT) in preventing the formation of oxidants derived from catechol dependent quinones. Methylation of hydroxycatechols prevents the formation of quinones and semiquinones which are significant sources of reactive oxygen species. The Pmel17/gp100 protein which is thought to play a role in the conversion of 5,6-dihydroxyindole and 5,6-dihydroxyindole-2-carboxylic acid to eumelanin is homologous to the antigen recognised by the HMB-45 antibody. Positive staining of lung cells with HMB-45 is a highly characteristic feature of LAM disease.
We hypothesised that LAM may be associated with a deficiency in COMT activity. If correct, this hypothesis would help explain why LAM is found exclusively in women, and why the lung tissue stains positively for HMB-45. Patients with LAM may have defective COMT activity in peripheral blood cells such as mononuclear phagocytes. Blood mononuclear phagocytes migrate to the lung and mature to become resident alveolar and interstitial macrophages. A defect in COMT activity would result in the accumulation of melanin and/or catecholoestrogens with the potential of generating superoxide and hydroxyl radicals. These oxidants could then induce tissue destruction, tumours, and smooth cell proliferation.
To verify this hypothesis, blood samples were collected from women with LAM and compared with matched controls. COMTgenotype was analysed to determine whether women with LAM disease would be prone to carrying the low activity alleles ofCOMT(COMT LL). The level of COMT activity in blood cells was also measured, and the presence of a potential COMT inhibitor was verified. The plasma concentration of malondialdehyde (MDA) was also measured since defective COMT activity or altered catechol compound metabolism would generate more oxidative damage.
Methods
STUDY POPULATION
Fifteen women of mean (SD) age 42.6 (9.2) years with clinical characteristics of LAM were recruited through Canada and the USA with the assistance of the LAM Foundation. Seventeen healthy age matched women of mean (SD) age 38.9 (9.8) years in Canada and the USA were recruited as a control group. Experimental protocols were approved by the institutional review board for human studies. Informed consent was obtained from all participants. Questionnaires were completed to determine whether participants had other diagnosed diseases, physiological conditions, or medication that could modify oestrogen metabolism. All subjects were non-smokers. The characteristics of the studied population are summarised in table 1.
Characteristics of LAM and control populations
COMT GENOTYPE
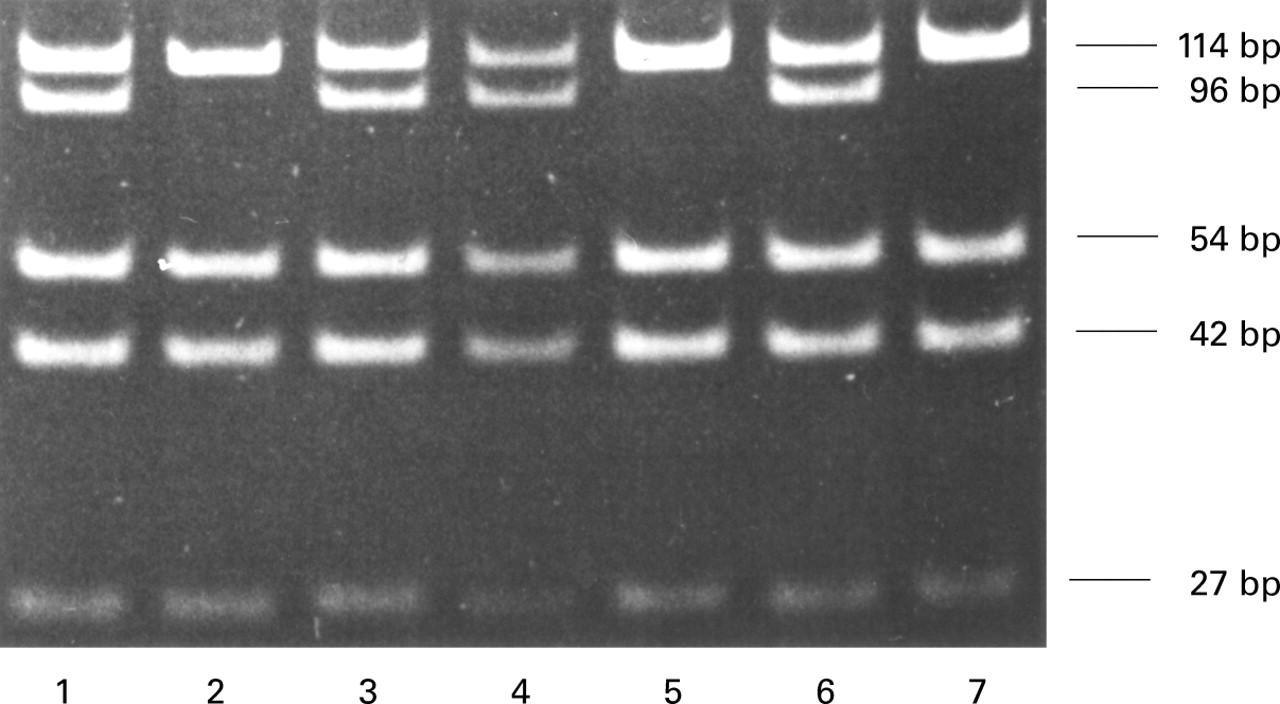
The presence of the G-to-A transition on exon 4 was used to analyse the polymorphic COMT genotype. Genomic DNA from LAM women and controls was subjected to a PCR based RFLP assay.17 A 237 bp fragment of theCOMT gene was first amplified by PCR using the forward primer TACTGTGGCTACTCAGCTGTGC and the reverse primer GTGAACGTGGTGTGAACACC. A 25 μl PCR reaction containing 50 ng genomic DNA, 250 μM each deoxynucleotide triphosphates, 300 nM each primer, 1 X reaction buffer (Perkin-Elmer, Branchburg, New Jersey, USA), and 0.6 unit Taq polymerase (Perkin-Elmer) was placed in a thermocycler (PTC-100, MJ Research Inc, Waltham, Massachusetts, USA). After denaturing for three minutes at 94°C the DNA was amplified for 35 cycles at 94°C for 30 seconds, at 60°C for 20 seconds, and at 72°C for 30 seconds, followed by a five minute extension at 72°C. An 8 μl sample of the PCR product was then digested for three hours at 37°C with 10 units of NlaIII (New England Biolabs, Beverly, Massachusetts, USA). The products of the restriction digest were combined with 1 μl of loading buffer (10% Ficoll and 0.25% xylene cyanol) and run on a 10% non-denaturing polyacrylamide gel in 1 X Tris-borate-EDTA buffer at 50 V for three hours. Restriction fragments of 27, 42, and 54 bp were present in every digested sample. The high and low activity alleles were detected by the presence of 114 bp and 96 bp fragments.
BLOOD SAMPLE PREPARATION
Blood samples were sent by courier and received within 24 hours. Precautions were taken to maintain the temperature at about 4°C using ice packs. Control blood samples were subjected to similar handling. Each sample was divided in two. Plasma was isolated from heparinised blood and used to assay for a potential COMT inhibitor. Determination of COMT activity in blood cells was performed in 200 μl whole blood collected in 2 ml heparinised tubes. A volume of 800 μl ice cold distilled water was added to lyse the blood cells and 100 μl of a suspension of sodium Chelex-100 was added to eliminate the calcium ion (Ca2+), a strong inhibitor of the COMT enzyme. The lysed samples and the resin were mixed with a tube rotator at 12 rpm for one hour at 4°C. Samples were then centrifuged at 7000g for 10 minutes and the supernatant was removed for the determination of COMT activity.
DETERMINATION OF COMT ACTIVITY
Determination of COMT activity was performed according to an established procedure.18 The 3,4-dihydroxybenzoic acid was used as a catechol substrate to measure the level of COMT dependent methylation activity. Briefly, 20 μl of the supernatant from each Chelex-100 treated sample of lysed whole blood was added to 180 μl Tris-Mg buffer (0.08 M Tris-HCl, pH 7.5, 1 mM MgCl2) and 100 μl of reaction buffer (0.08 M Tris-HCl, pH 7.5, 1 mM MgCl2, 2.8 μM S-adenosyl-l-(Me-14C)methionine, 20.2 μM non-radioactive S-adenosyl-l-methionine, 1 mM 3,4-dihydroxybenzoic acid, 4.2 mM dithiothreitol, and 0.64 units of adenosine deaminase.19 The reaction mixture was incubated for 90 minutes at 37°C in a shaker water bath and the reaction was stopped by the addition of 100 μl of 1.0 N HCl; 2 ml of toluene was then added to each tube. The tubes were vortexed for 10 seconds, centrifuged at 700g for 10 minutes, and the organic phase was added to counting vials containing 10 ml toluene fluor based liquid scintillation. To verify that the Me-14C compound extracted with toluene was the methylated dihydroxybenzoic acid, compounds were separated by HPLC on a Spherisorb ODS-2 column (5 μm, 25 cm × 0.46 cm) with a mobile phase containing 15% methanol and 85% 30 mM sodium citrate at pH 4.75 eluted at a flow rate of 1.0 ml/min.20 The compounds were detected by fluorescence (Ex = 310 nm, Em = 420 nm). Retention times of dihydroxybenzoic acid and its methylated derivatives were obtained by injecting appropriate standards.
COMT activity was expressed as pmol 4-hydroxy-3-methoxybenzoic acid formed per minute per mg protein quantified in the lysed whole blood with the Bio-Rad protein assay (Bio-Rad, Hercules, California, USA).
PRESENCE OF POTENTIAL COMT INHIBITOR
The presence of a circulating inhibitor of the COMT enzyme was verified as follows. Volumes of 10 μl or 50 μl plasma from LAM patients or controls were added to 12 units of a commercial preparation of COMT (Sigma, # C1897, porcine liver extract, 2500 U/mg). The enzymatic assay was performed as mentioned previously. The presence of a circulating inhibitor was determined by comparing the level of COMT activity measured in the presence and absence of plasma.
MDA
Using gas chromatography-mass spectrometry (GC-MS), the MDA can be detected in femtomole quantities in biological samples.21The analysis was performed with a Hewlett Packard 5890 Series II gas chromatograph interfaced to a 5989A mass spectrometer. The results were expressed as pmol MDA/ml plasma.
STATISTICAL ANALYSIS
The results are expressed as mean (SD). Statistical comparison between the two groups was analysed using the pairedttest.
Results
The COMT activity is dependent on the presence of high and low activity alleles of this enzyme. A PCR based RFLP assay was used to determine whether women with LAM were preferentially carrying the low activity alleles (fig 3). Heterozygous COMTwas detected by the presence of two bands of 114 bp and 96 bp on the polyacrylamide gel while a single band of 114 bp or 96 bp indicated the presence of homozygousCOMT HH orCOMT LL, respectively. Since LAM is a very rare disease, each group represents a small number of subjects. Nevertheless, the proportion ofCOMT alleles was similar for the two groups (table 2). The distribution ofCOMT HH,COMT LL, andCOMT HL genotype was 40%, 7%, and 53%, respectively, in the LAM group, which is in close agreement with the control group which had a distribution of 38%, 6%, and 56%. Interestingly, we found fewerCOMT LL cases than Thompsonet al 22 who found 19% ofCOMT LL, perhaps because of the small numbers of subjects in our groups. Nevertheless, the low incidence of COMT LL in this LAM group (1/15) suggests that the presence of low activity alleles ofCOMT is not required for the expression of this disease.

{kind=link}
{kind=link}
{kind=link}
PCR-based RFLP analysis for the COMT genotype. Representative polyacrylamide gel of the COMT exon 4 amplified by PCR and digested by the NlaIII restriction enzyme. Lanes 1, 3, 4, and 6 contain DNA from heterozygote individuals (COMTHL) and lanes 2, 5, and 7 contain DNA from homozygote individuals for the high activity allele (COMTHH).
Distribution of COMT genotype
If alterations in oestrogen dependent mesenchymal cell signalling in LAM result from defective COMT activity, then catecholoestrogen metabolism may be associated with the generation of ROS. To verify this hypothesis the enzymatic activity of COMT in whole blood of LAM women was measured. The blood samples were treated with Chelex-100 to eliminate the Ca2+ ion, a potent inhibitor of COMT, and the COMT activity was determined with an enzymatic assay using the 3,4-dihydroxybenzoic acid as a catechol substrate (table 3). No significant difference in COMT activity was detected between the two groups (24.2 (12.3) pmol/min/mg protein in the LAM group and 24.1 (6.3) pmol/min/mg protein in the control group). The capacity of blood cells from women with LAM to prevent the generation of ROS by methylating the catecholoestrogens was therefore not altered.
Cumulative results
Another possibility is that the activity of COMT may be reduced by the presence of a circulating inhibitor in the plasma. This hypothesis was verified by incubating 10 μl or 50 μl plasma from the two groups of women with a preparation of commercial COMT (table 3). The plasma used was not treated by Chelex-100 or any other procedure. Since the Ca2+ was still present, the activity obtained with the commercial COMT was reduced by 6–33 times according to the volume and sample of plasma tested. There was no statistical difference between the final COMT activity obtained by the addition of plasma from LAM or control women, which indicates that women with LAM do not carry a specific inhibitor for COMT enzyme in their plasma.
According to the COMT pathway hypothesis, altered catechol compound metabolism would result in overproduction of ROS which could increase the level of oxidative damage found in blood. The plasma concentration of MDA, a secondary product generated from lipid peroxidation, has been used as an internal marker of oxidative stress (table 3). Using a GC-MS analysis, the average MDA level in the women with LAM was 178.5 (80.1) pmol/ml plasma, which was not significantly different from the control group in whom a mean level of 155.4 (75.0) pmol/ml plasma was detected. Therefore, according to this assay, LAM is not associated with an increased level of oxidative stress, as detectable in blood.
Discussion
LAM is an uncommon lung disease for which the aetiology remains a mystery and an effective treatment has yet to be defined. The potential relationship with hormonal factors has frequently been suggested since LAM affects almost exclusively premenopausal women.2 It has been hypothesised that alterations in oestrogen metabolism may be involved in the pathophysiology of LAM. This study is the first to verify this hypothesis, and specifically addresses whether the catechol-O-methyltransferase pathway is defective in LAM. This pathway is particularly important since a failure in COMT activity would result in a continuously higher production of hydroxyoestrogen derived ROS, molecules known to induce smooth muscle cell proliferation.
Our results have shown that the genotype ofCOMT, as well as the enzymatic activity of COMT, did not differ significantly between the women with LAM and the control subjects, and that no specific inhibitor of COMT was detectable in blood. To eliminate definitively the possible involvement of ROS overproduction by catecholoestrogen in LAM disease, it remains to be established whether the blood and lung levels of 2- and 4-hydroxyoestradiol and 2- and 4-hydroxyoestrone are increased. Higher activity of the 2-hydroxylase and 4-hydroxylase pathways leading to these oestrogen metabolites have already been detected in breast carcinoma.23 It has been suggested that these alterations play an important part in the development of breast cancer,24 and that a higher level of these hydroxyoestrogens (catecholoestrogens) induces the development of resistance against the anticancer agent methotrexate.25
Our results also indicated that the level of MDA was not modified, which suggests that there was no increase in systemic oxidative stress. However, we cannot exclude the possibility that ROS are present at a higher level in lung tissue with the potential to act as second messengers.8 It would therefore be of interest to determine whether some signalling pathways sensitive to ROS are activated in women with LAM. On the other hand, the superoxide anion generated by catecholoestrogens reacts very rapidly with nitric oxide (rate constant 7 × 109/M/s) to form the highly reactive peroxynitrite anion,26 a product not detected by the MDA assay. Since the concentration of nitric oxide can be important in the lung,27 alteration in the production of either nitric oxide or superoxide anion could have deleterious effects and might deserve to be investigated.
Thus, this study showed that there were no significant differences in the COMT pathway in women with LAM. The relationship between LAM and altered oestrogen metabolism should now therefore be focused on other oestrogen metabolic pathways responsible for the production or elimination of oestrogens. Since smooth muscle cell proliferation can be stimulated by oestradiol,28 an alternative hypothesis may be related to a lower activity of 17β-HSD oxidase or a higher activity of 17β-HSD reductase in women affected by LAM, both leading to overproduction of oestradiol.
Acknowledgments
This research project has been supported by the LAM Foundation. The authors would like to thank Ginette Bilodeau and Marc Martel for expert technical assistance.