Article Text

Statistics from Altmetric.com
Cytokines are usually extracellular signalling proteins, usually less than 80 kD in size, and many are glycosylated. They are produced by many different cell types that are involved in cell-to-cell interactions acting through specific receptors on the surface of target cells. Cytokines usually have an effect on closely adjacent cells and therefore function in a predominantly paracrine fashion, although they may also act at a distance (endocrine) and may have effects on the cell of origin (autocrine). Cytokines may be regarded as a mechanism for cell-cell communication, and within this group may be included growth factors and cytokines with primarily chemoattractant properties (chemokines). They act on target cells to cause a wide array of cellular functions including activation, proliferation, chemotaxis, immunomodulation, release of other cytokines or mediators, growth and cell differentiation, and apoptosis. Cytokines were originally characterised (and named) according to some aspect of their functional activity that was initially discovered, but the cloning of the genes for these cytokines has now provided a better insight into their classification and grouping. It is apparent that there is a wide pleiotropy and element of redundancy in the cytokine family in that each cytokine has many overlapping functions, with each function potentially mediated by more than one cytokine.
The effect of an individual cytokine in the context of disease may not be easy to predict because it may be influenced by other cytokines released simultaneously from the same cell or from target cells following activation by the cytokine. The effects of cytokines are mediated by binding to cell surface high affinity receptors usually present in low numbers. The number of these receptors can be upregulated with cell activation, and there the effect of a cytokine may depend on the modulation of its receptors. Cytokines themselves may induce the expression of receptors which may change the responsiveness of both source and target cells. Two examples are the actions of interferon γ (IFN-γ) in decreasing the effect of tumour necrosis factor α (TNF-α) receptors on macrophages1 and that of interleukin 1β (IL-1β) in increasing the expression of the same receptors.2 Some cytokines may stimulate their own production in an autocrine manner, whereas others stimulate the synthesis of different cytokines that have a feedback stimulatory effect on the first cytokine resulting in an increase in its effects.
Inflammation in asthma
Cytokines play an integral role in the coordination and persistence of the inflammatory process in the chronic inflammation of the airways in asthma since they are capable of inducing many of the pro-inflammatory effects characteristic of this disease (table 1). Many cytokines are expressed and their function in this process can be surmised from a knowledge of their properties or from information obtained from animal studies. The chronic airway inflammation of asthma is unique in that the airway wall is infiltrated by T lymphocytes, eosinophils, macrophages/monocytes and mast cells, and sometimes by neutrophils too.3-6 In addition, an acute-on-chronic inflammation may be observed with acute exacerbations, with an increase in eosinophils and neutrophils in the airway submucosa and release of mediators such as histamine and cysteinyl leukotrienes (cys-LTs) from eosinophils and mast cells to induce bronchoconstriction, airway oedema, and mucus secretion. Changes in the resident cells are also observed in the asthmatic airway—for example, an increase in the thickness of the airway smooth muscle with hypertrophy and hyperplasia,7 more myofibroblasts with an increase in collagen deposition in the lamina reticularis,8 more vessels9 and an increase in the goblet cell numbers in the airway epithelium.10 Clearly, these chronic and acute inflammatory changes observed in the asthmatic airway could result from excessive release of many types of cytokines which has been observed in experimental induction of asthma by allergen exposure and virus infections, or during symptomatic asthma.11-14 Not only are cytokines involved in maintaining the chronic inflammatory process, they are also responsible for the initiation or the early stages of this process.
Summary of effects of cytokines
It is not simple to classify the numerous cytokines that are potentially involved in asthma because of their pleiotropic nature and overlapping properties. However, with regard to the specific abnormalities of asthma and to our current understanding of the pathogenesis of asthma, they may be grouped as follows:
- (1)
- Lymphokines: IL-2, IL-3, IL-4, IL-5, IL-13, IL-15, IL-16, IL-17.
- (2)
- Pro-inflammatory cytokines: IL-1, TNF, IL-6, IL-11, GM-CSF, SCF.
- (3)
- Anti-inflammatory cytokines: IL-10, IL-1ra, IFN-γ, IL-12, IL-18.
- (4)
- Chemotactic cytokines (chemokines): RANTES, MCP-1, MCP-2, MCP-3, MCP-4, MCP-5, MIP-1α, eotaxin, IL-8.
- (5)
- Growth factors: PDGF, TGF-β, FGF, EGF, IGF.
Role and source of cytokines in asthma
The CD4+ T lymphocytes of the asthmatic airways express a panel of cytokines including IL-3, IL-4, IL-5, IL-10, IL-13 and granulocyte-macrophage colony stimulating factor (GM-CSF), indicating that these T lymphocytes are of the T helper type 2 (Th2).15-17 The primary signals that activate Th2 cells are unknown but may be related to the presentation of a restricted panel of antigen in the presence of appropriate cytokines. Dendritic cells are ideally suited to being the primary contact between the immune system and external allergens. Co-stimulatory molecules on the surface of antigen presenting cells, in particular B7.2/CD28 interaction, may lead to proliferation of Th2 cells (fig1).18 With the expression of IL-4, synthesis of IgE by B lymphocytes on immunoglobulin isotype switching occurs.19Other cytokines, including TNF-α and IL-6 may also be important. The IgE produced in asthmatic airways binds to FcεRI receptors (“high affinity” IgE receptors) on mast cells, priming them for activation by antigen. The development of mast cells from bone marrow cells represents a process of maturation and expansion involving growth factors and cytokines such as stem cell factor (SCF) and IL-3 derived from structural cells. Mast cells recovered by bronchoalveolar lavage from asthmatic subjects show an enhanced release of mediators such as histamine.20 Mast cells also elaborate IL-4 and IL-5.21
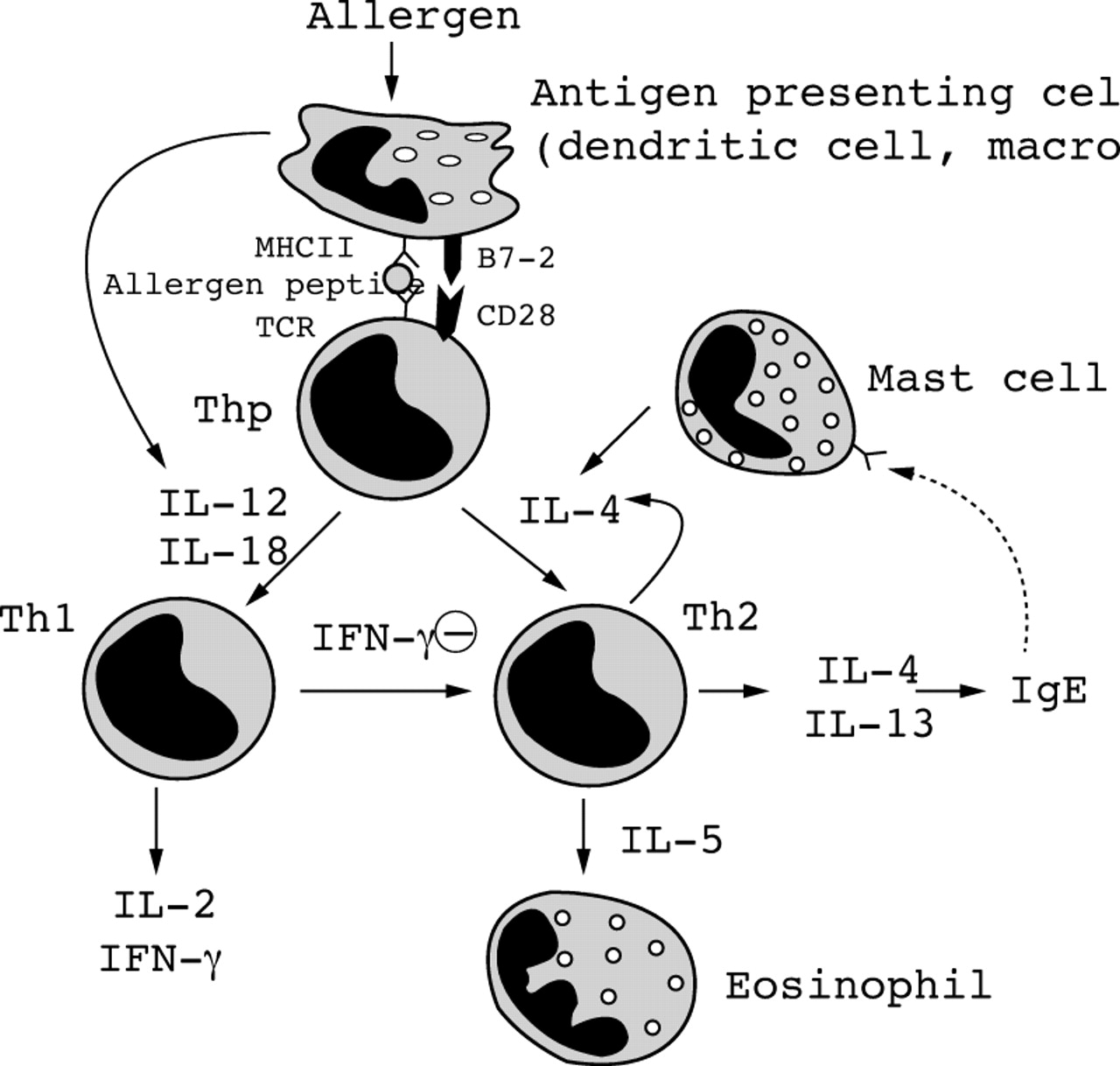
Scheme of cytokines involved in antigen presentation, activation of T helper progenitor (Thp) cells preferentially into Th2 rather than Th1 cells, and the effects of these cytokines. For abbreviations, see text.
IL-4 also increases the expression of an inducible form of the low affinity receptor for IgE (FcεRII or CD23) on B lymphocytes and macrophages.22 This may also account for the increased expression of CD23 on alveolar macrophages from asthmatic patients,23 which in turn could account for the increased release of cytokines from these macrophages.24 ,25 In addition, IL-4 is very important in driving the differentiation of CD4+ Th precursors into Th2-like cells.
The differentiation, migration and pathobiological effects of eosinophils may occur through the effects of GM-CSF, IL-3, IL-5 and certain chemokines such as eotaxin.26-31 IL-5 and eotaxin also induce the mobilisation of eosinophils and eosinophil precursors into the circulation.32 ,33 Once recruited from the circulation, mature eosinophils in the presence of these cytokines change phenotype into hypodense eosinophils which show increased survival in bronchial tissue.34 ,35 These eosinophils are primed for ligand mediated generation of increased amounts of cys-LTs and for cytotoxicity to other cells such as the airway epithelium.36-38 Eosinophils themselves may also generate other cytokines such as IL-3, IL-5, and GM-CSF.39 ,40
Cytokines may also play an important role in antigen presentation (fig1) and may enhance or suppress the ability of macrophages to act as antigen presenting cells. Normally, airway macrophages are poor at antigen presentation and suppress T cell proliferative responses (possibly via release of cytokines such as IL-1 receptor antagonist), but in asthma there is evidence for reduced suppression after exposure to allergen.41 ,42 Both GM-CSF and IFN-γ increase the ability of macrophages to present allergen and express HLA-DR.43 IL-1 is important in activating T lymphocytes and is an important co-stimulator of the expansion of Th2 cells after antigen presentation.44 Airway macrophages may be an important source of “first wave” cytokines such as IL-1, TNF-α and IL-6, which may be released on exposure to inhaled allergens via FcεRII receptors. These cytokines may then act on epithelial cells to release a second wave of cytokines, including GM-CSF, IL-8, and RANTES, which then amplify the inflammatory response and lead to influx of secondary cells such as eosinophils, which themselves may release multiple cytokines (fig 2).

Cytokines released from airway epithelial cells following various stimuli and other cytokines released from other cells such as macrophages. Cytokines from airway epithelial cells have effects on other cell types such as eosinophils, lymphocytes, airway smooth muscle cells, and fibroblasts. For abbreviations, see text.
Cytokines may also exert an important regulatory effect on the expression of adhesion molecules, both on endothelial cells of the bronchial circulation and on airway epithelial cells. Thus, IL-4 increases the expression of the adhesion molecule, VCAM-1, on endothelial and airway epithelial cells and this may be important in eosinophil and lymphocyte trafficking.45 IL-1 and TNF-α increase the expression of ICAM-1 in both vascular endothelium and airway epithelium.46 Cytokines also play an important role in recruiting inflammatory cells to the airways (table2).
Chemoattractant effects of cytokines
Proliferation of myofibroblasts and the hyperplasia of airway smooth muscle may occur through the action of several growth factors such as platelet-derived growth factor (PDGF) and transforming growth factor β (TGF-β). They may be released from inflammatory cells in the airways such as macrophages and eosinophils, but also by structural cells such as airway epithelium, endothelial cells, and fibroblasts. These growth factors may stimulate fibrogenesis by recruiting and activating fibroblasts or transforming myofibroblasts. Epithelial cells may release growth factors since collagen deposition occurs underneath the basement membrane of the airway epithelium.47 Growth factors may also stimulate the proliferation and growth of airway smooth muscle cells. PDGF and epidermal growth factor (EGF) are potent stimulants of human airway smooth muscle proliferation48 ,49 and these effects are mediated via activation of tyrosine kinase and protein kinase C. Growth factors may also be important in the proliferation of mucosal blood vessels and in the goblet cell hyperplasia that are characteristic of the chronically inflamed asthmatic airway. Cytokines such as TNF-α and fibroblast growth factors (FGF) may also play an important role in angiogenesis which is reported in chronic asthma.
Cytokine receptors
The receptors for many cytokines have now been cloned and, based on common homology regions, these have been grouped into superfamilies.50
CYTOKINE RECEPTOR SUPERFAMILY
This large receptor superfamily includes IL-2R β and γ chains, IL-4R, IL-3R α and β chains, IL-5 α and β chains, IL-6R, gp130, IL-12R and GM-CSFR. The extracellular regions of the cytokine receptor family contain combinations of cytokine receptor domains, fibronectin type III domains and usually C2 immunoglobulin constant region-like domains. Some members are comprised of a single polypeptide chain which binds its ligand with high affinity. For other receptors there may be more than one binding affinity for the ligand, typically high and low binding affinity sites. Additional subunits have been identified which are required for high affinity receptor expression. Some of these subunits are shared by more than one cytokine receptor, giving rise to heterodimeric structures, such as receptors sharing the GM-CSF receptor β-chain (IL-3, IL-5 and GM-CSF), receptors sharing the IL-6 receptor β-chain, gp130 (IL-6, leukaemia inhibitory factor, oncostatin M), and receptors sharing the IL-2 receptor γ-chain (IL-2, IL-4, IL-7 and IL-15).
Cytokine receptors may be secreted as soluble forms produced by alternative splicing of their mRNA transcripts to produce proteins lacking the transmembrane region and the cytoplasmic proximal charged residues which anchor the protein into the membrane. They may act as agonists or antagonists or as transport proteins to carry cytokines to other sites.
IMMUNOGLOBULIN SUPERFAMILY
Cytokine receptors with immunoglobulin superfamily domains in their extracellular sequences include IL-1R, IL-6R, PDGFR, and M-CSFR. The immunoglobulin domains are characterised by a structural unit of about 100 amino acids, with a distinct folding pattern known as the immunoglobulin fold.
PROTEIN KINASE RECEPTOR SUPERFAMILY
These receptors have glycosylated extracellular ligand binding domains, a single trans-membrane domain, and an intracellular tyrosine kinase catalytic domain. The superfamily includes receptors for growth factors such as PDGF, EGF, and FGF.
INTERFERON RECEPTOR SUPERFAMILY
This group includes IFN-α/β receptor, IFN-γ receptor, and IL-10 receptor. They are single spanning transmembrane glycoproteins, characterised by either one (IFN-γ and IL-10 receptors) or two (IFN-α/β receptors) homologous extracellular regions. Signal transduction involves phosphorylation and activation of JAK and TYK2 protein tyrosine kinases.
NERVE GROWTH FACTOR SUPERFAMILY
This includes cytokine receptors for NGFR, TNFR-I (p55), and TNFR-II (p75). These are characterised by three or four cysteine-rich repeats of about 40 amino acids in the extracellular part of the molecule. The mode of signal transduction has not been elucidated.
SEVEN TRANSMEMBRANE G PROTEIN COUPLED RECEPTOR SUPERFAMILY
These receptors include the chemokine receptors which have a characteristic structure of relatively short acidic extracellular N terminal sequence followed by seven transmembrane spanning domains with three extracellular and three intracellular loops. The receptors are coupled to heterotrimeric GTP binding proteins which induce phosphatidylinositol phosphate hydrolysis and activate kinases, phosphatases, and ion channels.
Cytokines in this review
Many cytokines are involved in the development of the atopic state and of the chronic inflammatory processes of asthma (fig 3), ultimately contributing to the release of mediators such as histamine and cys-LTs, airway remodelling, bronchoconstriction, and bronchial hyperresponsiveness. The potential role of each cytokine in these processes can be evaluated by studying their properties, their presence and localisation in the airway wall and airway secretions of patients with asthma, and the effect of specific inhibitors such as receptor antagonists or specific antibodies. Anti-inflammatory drugs for asthma may be developed by targeting inhibition of cytokine production and effects (such as cytokine antibodies, cytokine receptor antagonists, or blockers of specific signal transduction effects) or by using or modifying anti-inflammatory cytokines. We will consider each individual putative cytokine involved in asthma with regard to their synthesis and release, receptors, effects, and individual role in asthma and use the grouping proposed above. This is clearly necessary in order to appreciate the potential contribution of each cytokine and in view of the multiple functions that each cytokine has and that make one cytokine different from another. The potential role of each cytokine can be judged from its expression in asthmatic airways, from studies in transgenic or knock-out mice, or from studies involving the use of inhibitors of synthesis or antibodies or blockers at the receptor level. This approach we have taken does not underestimate the fact that cytokines work as a network.

{kind=link}
{kind=link}
{kind=link}
Interactions between resident and inflammatory cells and cytokines in the airways. For abbreviations, see text.
T cell derived cytokines (lymphokines)
INTERLEUKIN 2
Synthesis and release
Activated T cells, particularly Th0 and Th1 T cells, are a major source of IL-2,51 while B lymphocytes can be induced under certain conditions to secrete IL-2 in vitro. IL-2 is secreted by antigen activated T cells following activation, accompanied later by an upregulation of high affinity IL-2 receptors on the same cells. Binding of IL-2 to IL-2R induces proliferation of T cells, secretion of cytokines, and enhanced expression of receptors for other growth factors such as insulin. The IL-2 receptor complex is then removed from the T cell surface by internalisation. IL-2 can also be produced by eosinophils52 and by airway epithelial cells.53
Receptors
The IL-2 receptor complex is composed of three chains—α, β, and γ—and belongs to the family of haematopoietic cytokine receptors.54 ,55 The α and β chains bind to IL-2 with low affinity while the γ chain does not bind IL-2 alone. The high affinity complex is a heterotrimer of α/β/γ, while α/γ and β/γ heterodimers have an intermediate affinity. The β chain, which is expressed constitutively in T lymphocytes, is essential for signal transduction and the intracellular domain has critical sequences necessary for growth promoting signals.56 The γ chain also appears to be important for signal transduction57while the α chain alone is unable to transduce any signal.
Effects
IL-2 stimulates the growth and differentiation of T cells, B cells, natural killer (NK) cells, lymphokine activated cells, and monocytes/macrophages. IL-2 functions as an autocrine growth factor for T cells and also exerts paracrine effects on other T cells.58 IL-2 is also involved in TcR stimulated T cell apoptosis.59 It promotes the differentiation and immunoglobulin secretion of B cells. IL-2 acts on monocytes to increase IL-1 secretion, cytotoxicity, and phagocytosis.58Experiments with IL-2 gene knock out mice show that these animals develop a normal thymus and normal T cell subpopulations in peripheral tissues, indicating that IL-2 activity is redundant and not confined to IL-2 alone.60 Together with IL-4, IL-2 can reduce glucocorticoid receptor binding affinity of blood mononuclear cells.61 IL-2 stimulates NK cells to secrete IFN-γ, to proliferate and to increase cytolysis. IL-2 enhances the production of granulocyte-macrophage colony stimulating factor (GM-CSF) in peripheral blood mononuclear cells from asthmatics and IL-5 production from T cells from patients with the hypereosinophilic syndrome.62 ,63 IL-2 is a potent chemoattractant for eosinophils in vitro.64
Infusion of IL-2 as part of chemotherapy treatment results in eosinophilia with an associated increase in eosinophil colony stimulating activity.65 ,66 This activity was abolished by neutralising antibody to IL-3, IL-5 or GM-CSF, indicating that IL-2 is acting indirectly by promoting the synthesis of these cytokines. Repeated administration of IL-2 induces bronchial hyperresponsiveness in Lewis rats.67 In ovalbumin sensitised Brown-Norway rats IL-2 caused a threefold increase in the late phase response compared with the response in rats receiving only saline prior to allergen exposure.68 IL-2 caused an inflammatory response around the airways with a significant increase in eosinophils, lymphocytes, and mast cells.
Role in asthma
Levels of IL-2 are increased in bronchoalveolar lavage fluid of patients with symptomatic asthma.13 ,69 Increased BAL cells expressing IL-2 mRNA are also present,11 and a non-significant increase in IL-2 mRNA positive cells was observed in asthmatics following allergen challenge.70 Particularly high levels of IL-2 and IL-4 mRNA positive bronchoalveolar lavage cells are observed in steroid resistant asthmatics compared with steroid sensitive asthmatics71; this increase is not abolished by pretreatment with oral prednisolone in the steroid resistant patients and there were no differences in the expression of IL-5 and IFN-γ mRNA between the two groups. IL-2R bearing T lymphocytes are increased in the circulating blood of patients with acute severe asthma and in bronchoalveolar lavage cells recovered from asthmatics after allergen exposure.70 ,72
Cyclosporin A, which inhibits IL-2 gene transcription in activated T lymphocytes through interference with the transcription factors AP-1 and NF-AT, inhibits allergic airway eosinophilia but not bronchial hyperresponsiveness in animal models.73 However, in severe asthmatics cyclosporin A causes a reduction in the amount of oral steroid therapy needed to control asthmatic symptoms,74although not confirmed in another study.75 These effects of cyclosporin A may derive from an inhibition of IL-2 expression as well as an inhibition of other cytokines such as GM-CSF and IL-5.
INTERLEUKIN 3
Synthesis and release
Activated helper T cells are the predominant sources of IL-3, together with mast cells.76 ,77
Receptors
The IL-3 receptor is formed by the association of a low affinity IL-3 binding α subunit (IL-3Rα) with a second β subunit which is common to the IL-5 and GM-CSF receptors but does not itself bind to these cytokines.78 IL-3 binding to its receptor results in rapid tyrosine and serine/threonine phosphorylation of a number of cellular proteins including the IL-3Rβ subunit itself.79 ,80 A monoclonal antibody to the IL-3Rα chain abolishes its function.81 Human IL-3R is expressed on myeloid, lymphoid, and vascular endothelial cells. It is selectively induced in human endothelial cells by TNF-α and potentiates IL-8 secretion and neutrophil transmigration.82
Effects
IL-3 is a pluripotential haematopoietic growth factor that, together with other cytokines such as GM-CSF, stimulates the formation of erythroid, megakaryocyte, neutrophil, eosinophil, basophil, mast cell, and monocytic lineages.83 GM-CSF also increases the responsiveness of neutrophils to IL-3.84 Mice that overexpress IL-3 only show modest eosinophilia but die early due to massive tissue infiltration and destruction by myeloid cells such as neutrophils and macrophages.85
Role in asthma
An increase in the number of cells expressing IL-3 mRNA has been reported in mucosal biopsy specimens and in bronchoalveolar lavage cells of patients with asthma.11 ,86 However, after allergen challenge the numbers of IL-3 mRNA positive lymphocytes are not increased, in contrast to those expressing IL-5.70
INTERLEUKIN 4
Synthesis and release
IL-4 is derived from Th2 derived T lymphocytes and certain populations of thymocytes as well as eosinophils and cells of the basophil and mast cell lineage. Crosslinking of the CD40 ligand on human CD4+ T cells from normal non-allergic subjects generates a co-stimulatory signal that increases IL-4 synthesis.87Synthesis can also be induced by stimulation of the antigen receptor on T lymphocytes and by IgE Fc receptor crosslinking in mast cells and basophils. Interestingly, corticosteroidsenhance the capacity to induce IL-4 synthesis from CD4+ T cells.88 High affinity IL-4 receptors are abundant in activated B and T cells and are also present on haematopoietic progenitor cells, mast cells, macrophages, endothelial cells, epithelial cells, fibroblasts, and muscle cells.89-91
Receptors
The IL-4 receptor is a complex consisting of two chains, a high affinity IL-4 binding chain (p140, α chain) which binds IL-4 and transduces its growth promoting and transcription activating functions92 ,93 and the IL-2R γ chain (the common γ chain, γc) which amplifies signalling of the IL-4R.94 ,95 The α chain belongs to the cytokine receptor superfamily. A recombinant extracellular domain of the human IL-4R is a potent IL-4 antagonist.96 The IL-2R γ chain augments IL-4 binding affinity.94 ,95 A low affinity IL-4 receptor has also been identified.97 High affinity IL-4 receptors are abundant in activated B and T cells. They are also present on haematopoietic progenitor cells, mast cells, macrophages, endothelial cells, epithelial cells, fibroblasts, and muscle cells.89-91 Expression of the α subunit of the IL-4R has been localised to the airway epithelium, T cells, and mast cells in the airway mucosa, with greater expression in bronchial biopsy specimens from asthmatic subjects.98
IL-4 induces phosphorylation of the IL-4 induced phosphotyrosine substrate, which is associated with the p85 subunit of phosphatidylinositol-3 kinase and with Stat-6 and Janus protein kinase (JAK) after cytokine stimulation.99-102 The transcription factor Stat-6 is essential for mediating the effects of IL-4.103 ,104 IL-4 also stimulates phosphoinositol phosphate-2 hydrolysis, yielding IP3 and subsequent calcium flux followed by increased intracellular cAMP.105Interestingly, an association with atopy has been found with a R567 allele of the IL-4R α subunit106 which enhances signalling and decreases the binding of the phosphotyrosine phosphatase SHP-1 implicated in termination of signalling by means of cytokine receptors.102 ,107
Effects
IL-4 plays an important role in B lymphocyte activation by increasing expression of class II MHC molecules as well as enhancing expression of CD23, the low affinity (FcεRII) receptor, CD40 and the α chain of the IL-2 receptor. It promotes immunoglobulin synthesis by B lymphocytes and plays a central role in immunoglobulin class switching of activated B lymphocytes to the synthesis of IgG4 and IgE. This switching is accompanied by germline ε chain synthesis. IL-4 promotes the development of Th2-like CD4 T cells and inhibits the development of Th1-like T cells.108 ,109 It also enhances the cytolytic activity of CD8 cytotoxic T cells. Virus-specific CD8+ T cells can be induced by IL-4 to produce IL-5.110
IL-4 also exerts effects on monocytes and macrophages. It enhances the surface expression of MHC class II molecules and the antigen presenting capacity of macrophages, but inhibits the macrophage colony formation and cytokine release of TNF, IL-1, IL-12, IFN-γ, IL-8, and macrophage inflammatory protein 1α (MIP-1α). Together with other cytokines such as G-CSF and IL-6, IL-4 can promote the growth of mast cell and myeloid and erythroid progenitors. IL-4 also upregulates endothelial VCAM-1 expression on the endothelium. Interaction of VCAM-1 with the very late activation antigen 4 (VLA-4) promotes eosinophil recruitment.45 IL-4 also induces fibroblast chemotaxis and activation111 ,112 and, in concert with IL-3, IL-4 promotes the growth of human basophils and eosinophils.113
IL-4 has inhibitory effects such as suppression of metalloproteinase biosynthesis in human alveolar macrophages,114 inhibition of the expression of inducible nitric oxide synthase in human epithelial cells,115 and reduction of RANTES and IL-8 expression in human airway smooth muscle cells.116 ,117
Role in asthma
IL-4 is expressed by CD4+ and CD8+ T cells, eosinophils, and mast cells in both atopic and non-atopic asthma.118 ,119Increased numbers of lymphocytes expressing IL-4 mRNA together with IL-5 mRNA in bronchoalveolar lavage cells are observed following allergen challenge.86 The potential importance of IL-4 in inducing allergic airway inflammation has been addressed in IL-4 knock out mice. Sensitisation and exposure to ovalbumin did not induce lung eosinophilia as it did in the wild type litter mates.120No ovalbumin specific IgE was observed on active sensitisation and repeated exposures to ovalbumin did not induce bronchial hyperresponsiveness.121 IL-4 appears crucial to Th2 cell development. In IL-4 knock out mice T cells recovered from the airways do not synthesise a Th2 cytokine pattern, correlating with the absence of inflammatory airway changes.122 When wild type mice are treated with an anti-IL-4 antibody during the exposure to aerosolised ovalbumin but not during the sensitisation process, the influx of eosinophils to the airways is not inhibited.122 ,123 IL-4 receptor blockade prevented the development of antigen induced airway hyperresponsiveness, goblet cell metaplasia, and pulmonary eosinophilia in a mouse model.124 Inhalation of IL-4 by asthmatics causes an increase in eosinophil numbers in induced sputum, together with a transient increase in bronchial responsiveness.125IL-4 overexpression in mouse airways induces mucin MUC5AC gene expression and mucin hypersecretion, indicating a potential role for IL-4 in mucus hypersecretion.126
INTERLEUKIN 5
Synthesis and release
IL-5 was first isolated from supernatants of activated murine spleen cells which were shown to induce eosinophil colony formation. The isolated soluble activity was shown to stimulate eosinophil production from murine bone marrow selectively and was termed eosinophil differentiation factor. IL-5 was isolated from this soluble activity.127 It is produced by T lymphocytes and an increased expression of IL-5 mRNA has been demonstrated in CD4+ T cells in asthmatic airways using in situ hybridisation.128Bronchoalveolar lavage CD4+ and CD8+ T cells can also secrete IL-5.129 Human eosinophils can express IL-5 mRNA and release IL-5 protein in vitro130 and endobronchial challenge results in IL-5 mRNA expression in eosinophils in BAL fluid131 with an increase in IL-5 concentrations of up to 300-fold.132 ,133 Raised IL-5 concentrations have been reported in BAL fluid from symptomatic but not asymptomatic asthmatic subjects.134 Increased circulating levels of immunoreactive IL-5 have been measured in the serum of patients with exacerbations of asthma and these levels fall with corticosteroid treatment.135 IL-5 levels are raised in induced sputum following allergen challenge of asthmatic patients.136IL-5 protein has also been localised by immunochemistry in mast cells in bronchial biopsy specimens of patients with asthma together with IL-4, IL-6, and TNF-α.21 The transcriptional control of the human IL-5 gene involves several transcription factors including NF-AT.137
Receptors
The human IL-5R has been identified in vitro on eosinophils but not on neutrophils or monocytes.138 It consists of a heterodimer with two polypeptide chains, a low affinity binding α chain and a non-binding β chain shared with the IL-3R and GM-CSFR.139 Both chains belong to the cytokine receptor superfamily.140 The α subunit alone is sufficient for ligand binding and is specific for IL-5, but association with the β chain leads to a 2–3-fold increase in binding affinity and allows signalling to occur. Some IL-5R mutants have antagonistic effects and may act as receptor antagonists.141 Transcriptional regulation of the specific chain yields either membrane bound or soluble forms of the receptor (IL-5Rm and IL-5Rs).142 The membranous form interacts with the β subunit, leading to a substantial increase in affinity for IL-5.143 The soluble form is secreted in body fluids and interacts with IL-5 and antagonises the action of IL-5 on target cells.142 ,144 The expression of IL-5R is restricted to eosinophils and their immediate precursors. The number of cells in bronchial biopsy specimens from asthmatic subjects expressing both forms of the receptor is increased, with the expression of IL-5R mRNA being predominantly in eosinophils.145 An increase in membrane bound IL-5R mRNA on bone marrow progenitor cells (CD34+) occurs following allergen challenge of atopic asthmatic subjects.146
There are two major signalling pathways of IL-5 in eosinophils. IL-5 activates the tyrosine kinases Lyn, Syk and JAK2 and propagates signals through the Ras-MAPK and JAK-STAT pathways. For eosinophil survival Lyn, Syk and JAK2 tyrosine kinases and SHP-2 tyrosine phosphatase are important, while for eosinophil degranulation and adhesion molecule expression Raf-1 kinase is critical.147
Effects
IL-5 can influence the production, maturation, and activation of eosinophils. It acts predominantly at the later stages of eosinophil maturation and activation148 ,149 and can also prolong the survival of eosinophils.150 IL-5 appears to be the main cytokine involved in the development of eosinophilia in vivo. Administration of exogenous IL-5 causes eosinophilia in many in vivo models.151 IL-5 transgenic mice in which transcription of IL-5 is coupled to the dominant control region of the gene encoding for the constitutive marker CD2 show lifelong eosinophilia in organs with predicted T cell expression such as bone marrow, spleen and peritoneum, with fewer cells in the airway mucosa.85 IL-5 transgenic mice behave normally, indicating that eosinophils need other factors for degranulation and subsequent tissue damage. Thus, intratracheal administration of another eosinophil chemotactic agent, eotaxin, leads to further eosinophil accumulation in the lungs with bronchial hyperresponsiveness, an effect not observed in wild type mice.152 IL-5 may cause eosinophils to be released from the bone marrow while local release of another chemoattractant may be necessary to cause tissue localisation of eosinophils.32On the other hand, IL-5 instilled into the airways of patients with asthma induce significant airway eosinophilia153 and inhaled IL-5 caused eosinophilia in induced sputum and bronchial hyperresponsiveness but had no effect on airway calibre154. The eosinophilotactic responses of BAL fluid of asthmatics during the pollen season is accounted for by IL-5 and RANTES.155
Role in asthma
IL-5 may play an important part in eosinophil maturation, chemoattraction, and activation in asthma, and may underlie bronchial hyperresponsiveness. It may also interact with other eosinophil chemoattractants and activators such as chemokines to activate and induce chemoattraction of eosinophils.32 ,156 The expression of IL-5 in tissues and cells from patients with asthma is discussed above. Studies with IL-5 monoclonal antibodies clearly support a role for IL-5 in asthma. Pretreatment with anti-IL-5 monoclonal antibodies can suppress allergen induced airway eosinophilia.157-160 There is some debate about whether the IL-5 induced eosinophilia is the direct cause of bronchial hyperresponsiveness induced by allergen exposure. In some studies there is an effect of anti-IL-5 antibodies on bronchial hyperresponsiveness,157 ,159 while such an effect is not reported in another study despite inhibition of eosinophilia.123 In IL-5 knock out mice both allergen induced eosinophilia and airway hyperresponsiveness are abolished.161 The site of IL-5 expression may be critical to eosinophil recruitment and the development of airway hyperresponsiveness. Transgenic mice overexpressing IL-5 in lung epithelial cells showed raised levels of IL-5 in BAL fluid and serum, lung histopathological changes reminiscent of asthma, and display baseline airway hyperresponsiveness.162 On the other hand, studies in mice indicate that circulating but not local lung IL-5 is required for the development of antigen induced airways eosinophilia.163 Indeed, sensitisation and allergen challenge of mice leads to an increase in IL-5 producing T cells in the bone marrow.164 In addition to its effect in mobilising eosinophils from the bone marrow, there is evidence for its effect as a regulator of eosinophil homing and migration into tissues in response to local chemokine release.165
Studies of anti-IL-5 antibodies in human asthma are currently under way. In patients with worsening asthma, systemic corticosteroids reduces the expression of IL-5 mRNA in the airways mucosa associated with an improvement in asthma.166 Cyclosporin A and tacrolimus (FK506), immunosuppressant agents sometimes used in the treatment of severe asthma, inhibit the expression of IL-5 mRNA in activated human T lymphocytes in response to phytohaemagglutinin or phorbol esters.167
INTERLEUKIN 9
Synthesis and release
IL-9, originally identified as a T cell growth factor,168 is a T cell derived cytokine with pleiotropic effects on many cell types.169 It is produced in vitro and in vivo by CD4+ T cells, preferentially by the Th2 subset.170-173
Effects
IL-9 can stimulate the proliferation of activated T cells,168 ,174 ,175 enhance the production of immunoglobulins including IgE in B cells,176 and promote the proliferation and differentiation of mast cells177 ,178 and of haematopoietic progenitors.179 ,180 It strongly synergises with stem cell factor for the growth and differentiation of mast cells.181 IL-9 may upregulate the expression of mast cell proteases including the monocyte chemoattractant proteins mMCP-1, mMCP-2, mMCP-4178 and granzyme B.182 It may be involved in lymphomagenesis.183
Role in asthma
Transgenic mice created by expression of IL-9 regulated by a rat Clara cell 10 protein promoter showed lung selective expression of IL-9 with massive infiltration with eosinophils and lymphocytes, and increased numbers of mast cells within the airway epithelium.184 Epithelial cell hypertrophy associated with accumulation of mucus-like material within non-ciliated cells and increased subepithelial deposition of collagen was also observed. The mice also demonstrated marked bronchial hyperresponsiveness with normal baseline airway calibre.184 In another IL-9 transgenic mouse, eosinophilic airway inflammation, increased serum IgE levels, and bronchial hyperresponsiveness were observed.185
IL-9 has been suggested as a candidate gene predisposing to asthma on the basis of linkage disequilibrium between serum total IgE levels and a marker within the IL-9 gene which is situated on the 5q31-q33 chromosome.186 In inbred strains of mice, IL-9 has been identified as a factor regulating bronchial hyperresponsiveness.187 The human IL-9 receptor has been proposed as another potential asthma gene candidate188 and there are non-functional transcripts of IL-9 receptors.189
INTERLEUKIN 13
Synthesis and release
IL-13 is synthesised by activated CD4+ and CD8+ T cells and is a product of Th1-, Th2-, and Th0-like CD4 T cell clones.190Both CD4+ and CD8+ T cell clones synthesise IL-13 in response to antigen specific or polyclonal stimuli.191
Receptors
There is a close similarity between IL-4 and IL-13 receptors. An IL-4 receptor antagonist derived from a mutant protein192is a potent receptor antagonist of the biological activity of IL-4 and also of IL-13. It particularly inhibits the effect of IL-13 in inducing IgE synthesis in peripheral blood mononuclear cells. There is evidence from cDNA cloning of the IL-13 receptor to suggest that the IL-4R α chain is a component of IL-13R.193 However, despite this, these receptors appear to be distinct.191
Effects
IL-13 is a potent modulator of human monocyte and B cell function.190 It has profound effects on human monocyte morphology, surface antigen expression, antibody dependent cellular toxicity, and cytokine synthesis.190 ,194 IL-13, like IL-4, upregulates the expression of β1-integrin and VCAM-1 and the production of IL-6 and MCP-1 from human lung fibroblasts.195 On the other hand, in human monocytes stimulated by lipopolysaccharide, the production of proinflammatory cytokines, chemokines and colony-stimulating factors (IL-1β, IL-6, IL-8, IL-10, IL-12, IFN-γ, and GM-CSF) is inhibited by IL-13, while IL-1ra secretion is increased.196-198 MIP-1α, IL-1 and TNF-α release is inhibited from human alveolar macrophages.198 ,199 IL-13 inhibits the release of RANTES and IL-8 from airway smooth muscle cells in vitro.116 ,117These actions of IL-13 are similar to those of IL-4 and IL-10. The suppressive effects of IL-13 and of IL-4 are not related to endogenous production of IL-10. Similar to IL-4, IL-13 decreases the transcription of IFN-γ and of IL-12. It is possible that IL-13 acts like IL-4 and suppresses the development of Th1 cells by downregulating IL-12 production by monocytes, thereby favouring the development of Th2 cells.108 ,109 ,200 IL-13, unlike IL-4, fails to activate human T cells which appears to be due to a lack of IL-13 receptors on these cells. IL-13 diminishes monocyte glucocorticoid receptor binding affinity.201 It activates eosinophils by inducing the expression of CD69 cell surface protein and prolonging eosinophil survival.202
IL-13 induces the expression of CD23 on purified human B cells and acts as a switch factor directing IgE synthesis, similar to IL-4.203 ,204 A potent receptor antagonist of the biological activity of IL-4, a mutant protein of IL-4, antagonises IL-13 actions such as blocking the proliferation of B cells and IgE synthesis.205 This mutant protein of IL-4 may therefore have therapeutic potential for the treatment of allergies.
Role in asthma
An increased expression of IL-13 mRNA has been reported in the airway mucosa of patients with atopic and non-atopic asthma.206 ,207 In addition, levels of IL-13 together with IL-4 increased following segmental allergen challenge of patients with asthma.208 There is a significant correlation between the eosinophil counts and the levels of IL-13. A cloned piece of soluble IL-13a2-IgGfc fusion protein that specifically binds to and neutralises IL-13 without affecting IL-4 suppresses the increase in mucus secretion, eosinophilia and bronchial hyperresponsiveness following allergen exposure in sensitised mice.209 ,210 IL-13 administered to mice increases airway eosinophilia and bronchial hyperresponsiveness. IL-13 is therefore independently involved in the mouse sensitised model.
INTERLEUKIN 15
Synthesis and release
IL-15 is produced by both CD4+ and CD8+ T cells after activation.211 IL-15 mRNA is expressed in lung fibroblasts and epithelial cell lines as well as monocytes and human blood-derived dendritic cells.212
Effects
IL-15 shares some of the properties of IL-2, such as the stimulation of proliferation of T cells and lymphokine activated natural killer cells. However, there are many other distinct effects of IL-15. IL-15 can induce IL-8 and MCP-1 production in human monocytes.213 It also induces the release of soluble IL-2Rα from human blood mononuclear cells.214 It promotes angiogenesis in vivo215 and can also activate neutrophils and delay their apoptosis.216 IL-15 promotes the synthesis of IL-5 from house dust mite specific human T cell clones,217 an effect inhibited by the tyrosine kinase inhibitor, herbimycin A. This indicates that IL-15 produced at the site of allergic inflammation may play a part in recruitment and activation of eosinophils by inducing IL-5 production from T cells. IL-15 is also a chemoattractant for human blood T lymphocytes, an effect inhibited by an anti-IL-2R β chain antibody.218
Role in asthma
There are no data specific to asthma.
INTERLEUKIN 16
Synthesis and release
IL-16, previously known as lymphocyte chemoattractant factor, was first identified as a product of peripheral blood mononuclear cells following mitogen and histamine stimulation in vitro.219 ,220 Subsequently, it was shown to be produced by CD8+ T cells following stimulation with histamine and serotonin in vitro.221 ,222 IL-16 can also be produced by epithelial cells,223 eosinophils,224 and mast cells.225
Effects
IL-16 has specific activities on CD4+ T cells.226 It selectively induces migration of CD4+ including CD4+ T cells and CD4-bearing eosinophils.227 IL-16 acts as a growth factor for CD4+ T cells and induces IL-2R and MHC class II molecules on these cells.228
Role in asthma
Increased concentrations of IL-16 have been found in BAL fluid obtained from asthmatic subjects following allergen and histamine challenge.229 ,230 In stable atopic asthmatic subjects there is predominant expression of IL-16 mRNA and immunoreactivity in airway epithelium.231 In the ovalbumin sensitised and exposed mouse model IL-16 immunoreactivity was detected in the airway epithelium and an anti-IL-16 antibody prevented OVA specific IgE responses, bronchial hyperresponsiveness, but not airway eosinophilia.232
INTERLEUKIN 17
IL-17 is a CD4+ T cell derived cytokine which stimulates NF-κB and IL-6 production in fibroblasts and co-stimulates T cell proliferation.233 It stimulates epithelial, endothelial, and fibroblastic cells to secrete cytokines such as IL-6, IL-8 and GM-CSF, and PGE2.234 ,235 In the presence of IL-17, fibroblasts can sustain the proliferation of CD34+ haematopoietic progenitors and their preferential maturation into neutrophils. IL-17 increases the release of NO in cartilage from patients with osteoarthritis via NF-κB activation.236
INTERLEUKIN 18
IL-18 or IFN-γ-inducing factor (IGIF) is a cytokine which is a potent inducer of IFN-γ production and plays an important part in Th1 responses.237 Human IL-18 has been cloned from normal human liver cDNA libraries using murine IL-18 cDNA clones. IL-18 is synthesised as a precursor molecule without a signal peptide, but requires the IL-1-converting enzyme (ICE, caspase-1) for cleavage into a mature peptide. The human IL-18 receptor has been recently purified and characterised. Human IL-1 receptor protein is a functional IL-18 receptor component.238 IL-18 receptors are expressed selectively on murine Th1 cells but not on Th2 cells.239
Recombinant human IL-18 induces IFN-γ production by mitogen stimulated peripheral blood mononuclear cells and enhances natural killer (NK) cell cytotoxicity, increases GM-CSF production, and decreases IL-10 production. IL-18 induces IL-8, MIP-1α, and MCP-1 in human peripheral blood mononuclear cells in the absence of any co-stimuli. It directly stimulates gene expression and synthesis of TNF-α from CD3+/CD4+ T cells and NK cells, with the subsequent production of IL-1β and IL-8 from CD14+ monocytes.240IL-18 induces phosphorylation of p56 (1ck) and mitogen activated protein kinase, and these may be involved in TCR/CD3 mediated responses.241 IL-18 also activates NF-κB in murine Th1 cells for enhancement of IL-2 gene expression by Th1 cells.242 IL-18, together with IL-12, induces anti-CD40 activated B cells to produce IFN-γ, which inhibits IL-4 dependent IgE production.243 IL-18 and IL-12 have synergistic effects on Th1 development which may be due to reciprocal upregulation of their receptors.239
Pro-inflammatory cytokines
INTERLEUKIN 1
Synthesis and release
There are two forms of IL-1 (α and β) derived from two different genes. Although the amino acid sequence homology between human IL-1α and IL-1β is only 20%, the molecules bind to the same receptor and have nearly identical properties. IL-1β (17.5 kDa) is synthesised as a larger precursor molecule with a molecular weight of 31 kDa and is released into the extracellular space and the circulation. The most active form of IL-1β is its cleaved mature form resulting from the action of a specific cysteine protease, IL-1 converting enzyme.244 ,245
IL-1 is produced by a variety of cells including monocytes/macrophages, fibroblasts, B cells, both Th1 and Th2-like T cell lines, NK cells, neutrophils, endothelial cells, epithelial cells, airway smooth muscle cells, and vascular smooth muscle cells. The major source of IL-1 in most tissues is the stimulated monocyte/macrophage. Monocytes produce 10 times as much IL-1β as IL-1α,246 ,247 and IL-1α is mainly cell associated while IL-1β is mostly released. Eosinophils can produce IL-1α248 while human epithelial cells can augment IL-1β expression when exposed to the air pollutant nitrogen dioxide.249 A wide variety of stimuli including IL-1 itself,250 TNF-α,251GM-CSF,252 endotoxin, and phagocytosis can increase the expression of IL-1 in monocytes/macrophages. IL-1 production by endothelial and vascular smooth muscle cells can be induced also by IL-1, TNF, or endotoxin. On the other hand, PGE2 and corticosteroids can attenuate the capacity of endotoxin and other stimuli to release IL-1 through an inhibition of transcription and through a decrease in IL-1 mRNA stability.253-255 An inhibitor of the IL-1 converting enzyme inhibits inflammatory responses to IL-1β.256
Receptors
Two IL-1 receptors have been described. The type I receptor (CDw121a) and type II receptor (CDw121b) are transmembrane glycoproteins that bind IL-1α, IL-1β, and IL-1ra. IL-1R1 is expressed on many cells including T cells, B cells, monocytes, NK cells, basophils, neutrophils, eosinophils, dendritic cells, fibroblasts, endothelial cells, and vascular endothelial cells while IL-R2 is also expressed on T cells, B cells and monocytes. An IL-1R accessory protein (IL-1R-AcP) has been described257 which, when associated with IL-1R1, increases its affinity for IL-1β. Only IL-1R1 transduces a signal in response to IL-1,258 while IL-1R2 on binding to IL-1 does not. Thus, IL-1R2 may act as a decoy receptor, preventing IL-1 from binding to IL-1R1.259 IL-1 signal transduction pathways are associated with TNF receptor associated (TRAF) adaptor proteins, particularly TRAF-6.260 TRAF-6 associates with IL-1 receptor associated kinase (IRAK) which is recruited to and activated by the IL-1 receptor complex.261
A soluble receptor found in normal human serum and secreted by the human B cell line RAJI which binds preferentially to IL-1β has been described.262 IL-1 downregulates the numbers of IL-1 receptors263 ,264 while PGE2 increases the expression of IL-1 receptors.265 ,266 PDGF can increase IL-1 receptor expression and IL-1 receptor mRNA in fibroblasts,267 ,268 while IL-4 increases receptor expression on T cells.269 TGF-β may decrease the expression of IL-1 receptors,270 and may also uncouple the response of the cells to IL-1 without affecting IL-1 receptor expression or binding of IL-1.271
Some of the effects of IL-1 can be mimicked by agents that increase cAMP and activate protein kinase A,272 ,273 while others can be mimicked by agents that activate protein kinase C (PKC).274-276 Many cells produce cAMP in response to IL-1. Activation of protein kinase A by an IL-1 induced increase in cAMP may lead to increased transcription of a certain number of cellular genes. These may activate activating transcription factors (ATF) that bind to a cis-acting cAMP responsive element277 and NF-κB through the phosphorylation of an inhibitor protein, IκB. AP-1 activity may also be induced by IL-1278 through activation of PKC. Phosphorylation of several cellular proteins through the action of PKC independent serine/threonine kinase may also occur on activation of IL-1 receptor.279
Effects
IL-1 induces fever similar to other endogenous pyrogens such as TNF and IL-6. It partly causes leucocytosis by release of neutrophils from the bone marrow and induces the production of other cytokines including IL-6.
IL-1 is a growth factor for mature and immature thymocytes and a co-factor in the induction of proliferation and IL-2 secretion by peripheral blood CD4 and CD8 T cells following engagement of their antigen receptors. IL-1β is an important growth factor for Th2 cells in response to antigen primed antigen presenting cells but not for Th1 cells.280 Synergistic effects between IL-1 and IL-6 have been reported for the activation of T cells.281-283 It also functions as a growth factor for B cells.284-286IL-1 induces the induction of many other cytokines such as IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-8, RANTES, GM-CSF, IFN-γ, PDGF, and TNF from a variety of cells. It induces fibroblasts to proliferate,287 an effect that may be due to release of PDGF,288 increases prostaglandin synthesis and collagenase secretion,263 ,289 and increases the synthesis of fibronectin and types I, III, and IV collagen.290 Together with TNF-α and IFN-γ, IL-1β can induce or upregulate the expression of ICAM-1 and VCAM-1 on endothelial cells and also on respiratory epithelial cells which may lead to increased adhesion of eosinophils to the vascular endothelium and respiratory epithelium.291 ,292 IL-1 induced adhesion of eosinophils to endothelial cell monolayers is inhibited by anti-ICAM and anti-VCAM antibodies.293
Role in asthma
Levels of IL-1β in BAL fluid of patients with asthma were found to be increased compared with those of non-asthmatic volunteers, together with an increase in IL-1β specific mRNA transcripts in BAL fluid macrophages.294 In addition, patients with symptomatic asthma show increased levels of IL-1β in BAL fluid compared with patients with asymptomatic asthma.13Increased expression of IL-1β in asthmatic airway epithelium has been reported, together with an increased number of macrophages expressing IL-1β.295 Selective inhibition of IL-1β expression in the epithelium of the airway wall, without a reduction in IL-1ra expression after corticosteroid therapy, has been described in patients with asthma.296
IL-1β induces airway neutrophilia and increases airways responsiveness selectively to bradykinin in the rat.297IL-1β can induce eosinophil accumulation in rat skin, an effect blocked by an anti-IL-8 antibody.298 Of interest, IL-1β has profound effects on the coupling of the β2 adrenergic receptor to adenylyl cyclase, an effect mediated through the upregulation of inhibitory G proteins299 and the induction of cyclo-oxygenase 2 enzyme.300
TUMOUR NECROSIS FACTOR α(TNF-α)
Synthesis and release
Two major forms of TNF exist—TNF-α and TNF-β—which have only 35% amino acid homology but bind to similar receptors. TNF-α (previously known as cachectin) is expressed as a type II membrane protein attached by a signal anchor transmembrane domain in the propeptide.301 TNF-α is released from cells by proteolytic cleavage of the membrane bound form by a metalloproteinase, TNF-α converting enzyme (TACE). Inactivation of the TACE gene compromises the ability of cells to produce soluble TNF-α. TNF-α is produced by many cells including macrophages, T lymphocytes, mast cells, and epithelial cells, but the principal source is the macrophage. The secretion of TNF-α by monocytes/macrophages is greatly enhanced by other cytokines such as IL-1, GM-CSF and IFN-γ. Human eosinophils are also capable of releasing TNF-α,302 together with airway epithelial cells.303 TNF-β is mainly produced by activated lymphocytes.
Receptors
TNF-α interacts with two cell surface receptors, TNF-R55 and TNF-R75. Both receptors are members of the nerve growth factor receptor superfamily. Soluble p55 and p75 receptors have been described and are derived from the extracellular domain of each receptor. They may act as inhibitors of the effects of TNF.304 TNF receptors are distributed on nearly all cell types except red blood cells and resting T lymphocytes. The p75 receptor is more restricted to haematopoietic cells. TNF-R75 is the principal receptor released by human alveolar macrophages and monocytes in the presence of IFN-γ.305
Several signalling pathways leading to activation of different transcription factors such as NF-κB and AP-1 have been identified. The TNF receptor associated factor (TRAF) family of adaptor proteins, particularly TRAF-2, are involved in signalling from the TNF receptors.306 TRAF-2 may also have a role in the signal transduction pathway from the TNF receptor to the activation of mitogen activated protein (MAP) kinase cascades with subsequent activation of NF-κB and AP-1. TNF activates a sphingomyelinase resulting in the release of ceramide from sphingomyelin which in turn activates a Mg++ dependent protein kinase.307
Effects
Many of the actions of TNF-α occur in combination with other cytokines as part of the cytokine network and the effects of TNF-α are very similar to those of IL-1β as there is close interaction in the signal transduction pathway of these two cytokines.308TNF-α potently stimulates airway epithelial cells to produce cytokines including RANTES, IL-8 and GM-CSF,309-311and also increases the expression of the adhesion molecule ICAM-1.46 TNF-α also has a synergistic effect with IL-4 and IFN-γ to increase VCAM-1 expression on endothelial cells.312 This would have the effect of increasing the adhesion of inflammatory leucocytes such as neutrophils and eosinophils at the airway surface. TNF-α enhances the expression of class II MHC molecules on antigen presenting cells. In addition, it enhances the release of IL-1 by these cells. It also acts as co-stimulatory factor for activated T lymphocytes, enhancing proliferation and expression of IL-2 receptors. TNF-α inhibits bone resorption and synthesis and induces proliferation of fibroblasts.313 It also stimulates bronchial epithelial cells to produce tenascin, an extracellular matrix glycoprotein.314
Role in asthma
TNF-α may have an important amplifying effect on asthmatic inflammation.315 ,316 There is evidence of increased expression in asthmatic airways317 and IgE triggering in sensitised lungs leads to increased expression in epithelial cells in both rat and human lung.318 ,319 Increased TNF-α mRNA expression in bronchial biopsy specimens of asthmatic patients has been reported.320 TNF-α is also present in the BAL fluid of asthmatic patients13 and TNF-α release from bronchoalveolar leucocytes of asthmatic patients is increased.321 TNF-α is also released from alveolar macrophages of asthmatic patients after allergen challenge.322 Furthermore, both blood monocytes and alveolar macrophages show increased gene expression of TNF-α after IgE triggering in vitro and this effect is enhanced by IFN-γ.323 Alveolar macrophages of asthmatic patients undergoing late phase responses after allergen challenge release more TNF-α and IL-6 ex vivo than those from patients with only an early response.322 There are polymorphisms in the promoter of the TNF gene which may be more frequently associated with asthma.324 ,325
TNF-α increases airway responsiveness in Brown-Norway rats326 and in humans in association with an increase in sputum neutrophils.327 It may be an important mediator in initiating chronic inflammation by activating the secretion of cytokines from a variety of cells in the airways. Several approaches to inhibition of TNF-α synthesis of effects are now under investigation in asthma, including monoclonal antibodies to TNF and soluble TNF receptors.
INTERLEUKIN 6
Synthesis and release
IL-6 was originally described for its antiviral activity, its effects on hepatocytes, and its growth promoting effects on B lymphocytes and plasmacytomas. It is secreted by monocytes/macrophages, T cells, B cells, and other cells including fibroblasts, bone marrow stromal cells, keratinocytes, and endothelial cells. Epithelial cells also appear to produce IL-6.328 Human airway smooth muscle cells under activation with IL-1β or TGF-β can release IL-6.329 Major basic protein secreted from eosinophils can interact with IL-1 or TGF to increase IL-6 release from fibroblasts.330 IL-6 has also been localised to eosinophil granules.331
Effects
IL-6 is a pleiotropic cytokine whose role in asthma remains unclear. It has growth regulatory effects on many cells and is involved in T cell activation, growth, and differentiation. It is a terminal differential factor for B cells and induces immunoglobulin (IgG, IgA and IgM) secretion.332 IL-6 is an important co-factor in IL-4 dependent IgE synthesis.333 It upregulates the production of, and its response to, IL-2.
IL-6 may also have anti-inflammatory effects. It inhibits the expression and release of IL-1 and TNF from macrophages in vitro and endotoxin induced TNF production and neutrophil influx in the airways in vivo.334-336 In IL-6 transgenic mice there is a lymphocytic infiltration around airways associated with reduced airways responsiveness.337
Role in asthma
There is increased release of IL-6 from alveolar macrophages from asthmatic patients after allergen challenge322 and increased basal release compared with non-asthmatic subjects.13 IgE dependent triggering stimulates the secretion of IL-6 in both blood monocytes and alveolar macrophages in vitro.323 Increased levels of IL-6 can be measured in nasal washings of children following a rhinovirus infection.14 In addition, IL-6 mRNA expression with increased NFκB-DNA binding activity can be induced by rhinovirus infection of cells in vitro.14
INTERLEUKIN 11
Synthesis and release
IL-11, which is distantly related to IL-6, is produced by fibroblasts, epithelial cells and human airway smooth muscle cells when stimulated by IL-1 and TGF-β1.329 ,338A single class of specific receptor has been described on mouse cells.339
Receptors
The receptor has not yet been cloned. Like IL-6, IL-11 uses the IL-6 signal transducer gp130. On ligand binding, phosphorylation of tyrosine residues in a number of proteins occurs.340 ,341
Effects
Although IL-11 cDNA was cloned on the basis of IL-6-like bioactivity, IL-11 has distinct biological features from IL-6. IL-11 promotes multiple stages of human megakaryocytopoeisis and thrombopoeisis. In combination with stem cell factor or IL-4, IL-11 supports the generation of B cells, similar to IL-6.342IL-11 induces the production of acute phase reactants343and induces the synthesis of tissue inhibitor of metalloproteinase 1. It inhibits IL-12 and TNF-α production from monocytes/macrophages,344 effects mediated at the transcriptional level by inhibition of NF-κB.
Role in asthma
IL-11 can be detected in BAL fluid during upper respiratory viral infections in humans and induces non-specific bronchial hyperresponsiveness in mice.345 Targeted expression of IL-11 in mouse airways leads to a T cell inflammatory response with airway remodelling, local accumulation of myofibroblasts, and airways obstruction.346
GRANULOCYTE-MACROPHAGE COLONY STIMULATING FACTOR (GM-CSF)
Synthesis and release
GM-CSF is one of the colony stimulating factors that act to regulate growth, differentiation, and activation of haematopoietic cells of multiple lineages. GM-CSF is produced by several airway cells including macrophages, eosinophils, T lymphocytes, fibroblasts, endothelial cells, airway smooth muscle cells, and epithelial cells.
Receptors
The GM-CSF receptor consists of a low affinity α chain with a second affinity converting β chain which is also shared by IL-3 and IL-5 receptors.78 ,347 These receptors are usually distributed on granulocytes and monocytes, endothelial cells, and fibroblasts. Upregulation of the expression of α chain of GM-CSFR mRNA in macrophages in airway biopsy specimens of non-atopic asthma but not of atopic asthma has been reported.348 Certain analogues of GM-CSF bind to the α chain of the receptor but not to the β chain complex without agonist effect, indicating that these mutants could act as antagonists of GM-CSF.349
Effects
GM-CSF is a pleiotropic cytokine that can stimulate the proliferation, maturation, and function of haematopoietic cells. It may be involved in priming inflammatory cells such as neutrophils and eosinophils and can prolong the survival of eosinophils in culture.28 ,350 GM-CSF can enhance the release of superoxide anions and cys-LTs from eosinophils351 and can also induce the synthesis and release of a number of cytokines including IL-1 and TNF-α from monocytes. GM-CSF induces non-haematopoietic cells such as endothelial cells to migrate and proliferate.352
Role in asthma
There is increased expression of GM-CSF in the epithelium of bronchial biopsy specimens from asthmatic patients353 and in T lymphocytes and eosinophils after endobronchial challenge with allergen.13 ,354 Increased circulating concentrations have been detected in patients with acute severe asthma,355 and peripheral blood monocytes from asthmatic patients secrete increased amounts.356 GM-CSF accounts for the increased eosinophil survival activity of BAL fluid.31 It is the major LTC4 enhancing activity for eosinophils in the supernatant of cultured asthmatic alveolar macrophages.357 Media obtained from cultured bronchial epithelial cells of asthmatic subjects increases the viability, superoxide production, and LTC4 production by eosinophils in vitro,358 an effect abolished by a neutralising antibody to GM-CSF. Transient expression of the GM-CSF gene to the epithelium of rats using an adenoviral vector led to an accumulation of eosinophils and macrophages associated with irreversible fibrosis.359This indicates that GM-CSF may be involved in the chronic eosinophilia and airways remodelling of asthma.
STEM CELL FACTOR (SCF)
Synthesis and release
SCF (c-kit ligand) is produced by bone marrow stromal cells, fibroblasts (including bronchial subepithelial myofibroblasts and nasal polyp fibroblasts), and epithelial cells such as nasal polyp epithelial cells.360-362
Receptors
The receptor for SCF is c-kit, a receptor protein kinase, and is expressed on early haematopoietic progenitors and allows a synergistic response to SCF and lineage committing growth factors such as GM-CSF for myelocytes.c-kit expression decreases with cell maturation and is absent on mature cells released from bone marrow. However, c-kit expression increases on mast cells as they mature and are abundantly expressed on the surface of mast cells. c-kit is also expressed on human eosinophils.363
Effects
SCF acts as a survival factor for the early haematopoietic progenitor cells and synergises with other growth factors to regulate the proliferation and differentiation of cells. It is a major growth factor for human mast cells.364 ,365 Two alternative spliced variants account for the different forms of SCF: a primarily membrane bound and the other primarily soluble after being released from the cell surface by proteolysis.366 CD34+ bone marrow cells cultured in vitro with rh-SCF and IL-3 induces the development of mast cells and other haematopoietic lineages.367
Membrane bound SCF may influence mast cell adhesion368 and soluble SCF is chemotactic for mast cells.369 Removal of mast cells from either soluble or membrane bound SCF causes mast cells to undergo apoptosis.370 ,371 SCF has modest direct activating capacity on the mast cell but is usually more active in priming mast cell responses to other stimuli such as IgE stimulated mediator release.372-374 It causes the release of small amounts of IL-4 and TNF-α from human lung mast cells.375SCF stimulates VLA-4 mediated cell adhesion to fibronectin and VCAM-1 adhesion molecules on human eosinophils.363
Role in asthma
There is little information on the expression of SCF in asthmatic airways. It is expressed in the epithelium of nasal polyps from patients with allergic rhinitis.
Inhibitory cytokines
INTERLEUKIN 10
Synthesis and release
IL-10, previously known as cytokine synthesis inhibitor factor (CSIF), was originally identified as a product of murine T helper (Th2) clones that suppressed the production of cytokines by Th1 clones responding to stimulation of antigen.376 In humans Th0, Th1, and Th2-like CD4+ T cell clones, cytotoxic T cells, activated monocytes and peripheral blood T cells including CD4+ and CD8+ T cells have the capacity to produce IL-10.377 ,378 Mast cells also have the capacity to produce IL-10. Constitutive IL-10 secretion occurs in the healthy lung with the major source being the alveolar macrophage; however, the circulating monocyte elaborates more IL-10 than the alveolar macrophage.379
Receptors
IL-10R is a member of the class II subgroup of cytokine receptors, the IFN-receptor family. IL-10R has been characterised and cloned from a human lymphoma cell line380 and is expressed in several lymphoid and myeloid cells381 and also on NK cells.382 IL-10R is highly effective in recruiting the signalling pathways of IL-6 type cytokine receptors including STAT1 and STAT3.383 The inhibitory effect of IL-10 on monocytes appears to be dependent on NF-κB.384 The specific signalling pathway of IL-10R has not yet been definitely characterised.
Effects
IL-10 is a pleiotropic cytokine that can exert either immunosuppressive or immunostimulatory effects on a variety of cell types. It is a potent inhibitor of monocyte/macrophage function, suppressing the production of a number of pro-inflammatory cytokines including TNF-α, IL-1β, IL-6, MIP-1α, and IL-8385-387 although the release of MCP-1 is increased.387 IL-10 inhibits monocyte MHC class II, B7.1/B7.2 and CD23 expression and accessory cell function. Accessory signals mediated by B7 molecules through CD28 on the surface of T cells are essential for T cell activation. Expression of IL-10 by antigen presenting cells may be an established pathway for the induction of antigen specific tolerance such as that to allergens.388By contrast, IL-10 upregulates the monocyte expression of IL-1ra, another anti-inflammatory cytokine.389 IL-10 suppresses the synthesis of superoxide anions and NO by activated monocytes/macrophages.390 An IL-10 antibody enhances the release of cytokines from activated monocytes, suggesting that this cytokine may play an inhibitory role when the cell is stimulated.386 IL-10 inhibits the stimulated release of RANTES and IL-8 from human airway smooth muscle cells in culture.116 ,117 It inhibits the production of IFN-γ and IL-2 by Th1 lymphocytes376 and IL-4 and IL-5 production by Th2 cells by interfering with B7-CD28 dependent signals.391 ,392 IL-10 also inhibits eosinophil survival and IL-4 induced IgE synthesis. On the other hand, IL-10 acts on B cells to enhance their viability, cell proliferation, immunoglobulin secretion with the isotype switch and class II MHC expression. It decreases IL-4 induced IgE switch in peripheral blood mononuclear cells but potentiates IgE production in B cells that are already switched to produce IgE.393 IL-10 is also a growth co-stimulator for thymocytes and mast cells,394 as well as an enhancer of cytotoxic T cell development.395 It also activates the transcription of genes for mast cell derived proteases and enhances the production of the tissue inhibitor of metalloproteinases of monocytes and tissue macrophages while decreasing metalloproteinase biosynthesis.396
Role in asthma
There is significantly less IL-10 mRNA and protein expressed in alveolar macrophages of asthmatic subjects than in those from non-asthmatic individuals.25 ,397 Triggering of CD23 molecule by anti-CD23 monoclonal antibodies induces IL-10 production by human monocytes.398 An IL-10 polymorphism on the transcription initiation site could be responsible for reduced IL-10 release.397 Another polymorphism upstream from this site was associated with increased total serum IgE.399 Inhaled corticosteroid treatment restores the reduced IL-10 release from macrophages of asthmatic subjects25 and theophylline increases IL-10 secretion.400 On the other hand, other studies indicate that there are increased numbers of macrophages and T cells expressing IL-10 mRNA in the BAL fluid of patients with asthma.401
IL-10 inhibits the late response and the influx of eosinophils and lymphocytes after allergen challenge in the Brown-Norway rat.402 Co-instillation of IL-10 by the intranasal route significantly inhibits the peritoneal and lung eosinophilia induced by ovalbumin in immunised mice.403 ,404 Given its anti-inflammatory properties and these effects in animal models of allergic inflammation, IL-10 may have beneficial effects in asthma.405 However, no such studies have been performed yet. Administration of IL-10 to normal volunteers induced a fall in circulating CD2, CD3, CD4, and CD8 lymphocytes with suppression of mitogen induced T cell proliferation and reduction of TNF-α and IL-1β production from whole blood stimulated with endotoxin ex vivo.406
INTERLEUKIN 1 RECEPTOR ANTAGONIST (IL-1RA)
IL-1ra has been isolated from supernatants of monocytes cultured on aggregated immunoglobulin or with immune complexes,407 ,408 alveolar macrophages,409and urine of patients with fever or myelomonocytic leukaemia.410-412 IL-1ra shares 26% and 19% amino acid homology with IL-1α and IL-1β, respectively. It binds to the IL-1 receptor with a similar affinity to IL-1α or IL-1β413and inhibits most effects of IL-1 on cells, such as thymocyte proliferation, IL-2 synthesis by T cells, and PGE2 and collagenase production by fibroblasts.413-416 IL1-ra is preferentially produced by alveolar macrophages compared with monocytes,417 which may underlie the diminished IL-1 bioactivity produced by alveolar macrophages compared with monocytes.253 ,417 ,418 Other IL-1 receptor inhibitors have been described.419 ,420
IL-1ra blocks proliferation of Th2 but not Th1 clones in vitro.421 Increased expression of IL-1β and IL-1ra in asthmatic airway epithelium has been reported.295 After treatment with inhaled corticosteroids the expression of IL-1β is reduced but IL-1ra is unchanged, thus tipping the balance away from inflammation.296 In a human airway epithelial cell line corticosteroids increase the expression of IL-1ra.422 In an ovalbumin sensitised guinea pig model an aerosol of IL-1ra given immediately before allergen challenge resulted in protection against bronchial hyperresponsiveness and accumulation of pulmonary eosinophils.423 In a similar model the late phase response with the number of hypodense eosinophils in BAL fluid was inhibited.424 Trials of IL-1ra in asthma are underway.
INTERFERON γ
Synthesis and release
IFN-γ was originally identified as a product of mitogen stimulated T lymphocytes that inhibited viral replication in fibroblasts. The only known sources of IFN are CD4+ and CD8+ T cells and NK cells.
Receptors
IFN-γ receptor is a single transmembrane protein, a member of the cytokine receptor type II superfamily. Although the receptor binds IFN-γ with high affinity, signal transduction requires a species-specific accessory protein that associates with the extracellular domain of the receptor. The receptor is expressed on T cells, B cells, monocytes/macrophages, dendritic cells, granulocytes, and platelets. Epithelial and endothelial cells also express these receptors.
Effects
IFN-γ has extensive and diverse immunoregulatory effects on various cells. It is produced by Th1 cells and exerts an inhibitory effect on Th2 cells.425 IFN-γ inhibits antigen induced eosinophil recruitment in the mouse.426 However, it may also have pro-inflammatory effects and may activate airway epithelial cells to release cytokines and express adhesion molecules.427 IFN-γ has an amplifying effect on the release of TNF-α from alveolar macrophages induced by IgE triggering or by endotoxin323 ,428 and increases the expression of class I and class II MHC molecules on macrophages and epithelial cells. IFN-γ is a powerful and relatively specific inhibitor of IL-4 induced IgE and IgG4 synthesis by B cells.
IFN-γ increases the production of IL-1, PAF, and H2O2 from monocytes, in addition to downregulating IL-8 mRNA expression that is upregulated by IL-2.429-431 IFN-γ also synergises the effects of TNF-α in the production of RANTES from airway smooth muscle cells.116 On the other hand, it inhibits IL-10 production from monocytes,432 which in turn leads to an upregulation of TNF-α transcription.433 Thus, IFN-γ promotes cell mediated cytotoxic responses while inhibiting allergic inflammation and IgE synthesis.
IFN-γ upregulates class II molecules on monocytes/macrophages and dendritic cells and induces de novo expression on epithelial, endothelial and other cells, thus making them capable of antigen presentation.
Role in asthma
There is reduced production of IFN-γ by T cells of asthmatic patients and this correlates with disease severity.434 ,435 No polymorphisms of the IFN-γ gene have been associated with asthma.436 Administration of exogenous IFN-γ prevents the airway eosinophilia and hyperresponsiveness following allergen exposure in mice.437 ,438 Liposome mediated gene transfer of IFN-γ to the pulmonary epithelium in sensitised mice before secondary antigen exposure also inhibited the pulmonary allergic response.439 IFN-γ receptor knock out mice develop a prolonged airway eosinophilia in response to allergen.440IFN-γ inhibits allergic eosinophilia437 ,441 and airway hyperresponsiveness, probably by inducing the formation of IL-10. These studies indicate that IFN-γ has a potential modulating effect on allergen responses. Allergen immunotherapy of asthmatic patients results in increased production of IFN-γ by circulating T cells442 and in an increase in IFN-γ producing T cells in nasal biopsy specimens.443 Corticosteroid treatment also increases IFN-γ expression in asthmatic airways,444but in corticosteroid resistant patients IFN-γ is unexpectedly reduced.71 In asthmatic patients nebulised IFN-γ reduces the number of eosinophils in BAL fluid, indicating its therapeutic potential in asthma.445
INTERLEUKIN 12
Synthesis and release
IL-12 was initially recognised as a cytokine capable of synergising with IL-2 to increase cytotoxic T lymphocyte responses, and also as an inducer of IFN-γ synthesis by resting human peripheral blood mononuclear cells in vitro. IL-12 is secreted by antigen presenting cells including B lymphocytes, monocytes/macrophages, and dendritic cells.446 ,447
Receptors
IL-12 receptors are expressed on T cells and NK cells. One component of the IL-12 receptor complex is related to gp130.448
Effects
IL-12 enhances the growth of activated T cells and NK cells449-452 and enhances cytotoxic T cell and NK activity.449 ,453 ,454 IL-12 stimulates NK cells and T cells to produce IFN-γ,453 ,455-457 promotes in vitro differentiation of mouse and human T cells that secrete IFN-γ and TNF-α,451 ,455 ,458 ,459 and inhibits the differentiation of T cells into IL-4 secreting cells.458 ,459 IL-12 indirectly inhibits IL-4 induced human IgE responses by IFN-γ dependent and independent mechanisms in vitro.460 Thus, IL-12 can primarily regulate Th1 cell differentiation while suppressing the expansion of Th2 cell clones458 by early priming of undifferentiated Th cells for IFN-γ secretion.461 Thus, IL-12 may play an important role in directing the development of Th1-like T cell responses against intracellular pathogens whilst inhibiting the development of Th2-like responses and IgE synthesis. IL-12 may play an important role in inhibiting inappropriate IgE synthesis and allergic inflammation as a result of allergen exposure.
Role in asthma
IL-12 may play an important part in inhibiting inappropriate IgE synthesis and allergic inflammation as a result of allergen exposure. Treatment of mice with IL-12 during active sensitisation reduced antigen induced influx of eosinophils in BAL fluid, inhibited IgE synthesis, and abolished antigen induced bronchial hyperresponsiveness.462 Once an inflammatory response is established there is an inhibition of antigen induced bronchial hyperresponsiveness and inflammation.463 These effects of IL-12 are largely mediated by IFN-γ.464 In mice IL-12 administered at the time of allergic sensitisation decreased specific IgE, tracheal ring responsiveness to acetylcholine, and eosinophilia in BAL fluid after allergen challenge, together with IL-5 and IL-10 downregulation; IL-12 administered after sensitisation did not alter specific IgE levels, had little effect on tracheal ring responsiveness, and a modest effect on the recruitment of eosinophils, together with IL-5 downregulation but IL-12 upregulation.465 Thus, the effect of IL-12 is dependent on the timing of its administration in relation to active sensitisation.
The production of IL-12 and IL-12 induced IFN-γ release is reduced in whole blood cultures from patients with allergic asthma compared with normal subjects.466 There is a reduction of IL-12 mRNA expression in airway biopsy specimens of patients with allergic asthma compared with normal subjects but, following treatment with oral corticosteroids, the levels of IL-12 mRNA increased in patients with corticosteroid sensitive asthma while no significant changes were observed in those with corticosteroid resistant asthma.206This contrasts with the inhibitory effects of corticosteroids on IL-12 production in human monocytes in vitro.88 Allergen immunotherapy results in an increase in IL-12 expression.467 PGE2, β2agonists, and corticosteroids inhibit IL-12 production from monocytes.88 ,468 ,469
Chemokines
Chemokines are chemotactic cytokines of 8–10 kDa involved in attracting leucocytes into tissues. Over 40 chemokines have now been recognised470 and they are divided into families according to their structure. The two major groups are CC chemokines (β chemokines) in which two cysteine residues are adjacent to each other and CXC chemokines (α chemokines) in which these residues are separated by another amino acid. The CC chemokines are involved in chemoattraction of eosinophils, monocytes, and T lymphocytes and are therefore of greatest relevance to asthma.
CC CHEMOKINES
Synthesis and release
Macrophage inflammatory protein 1α (MIP-1α) and MIP-1β were purified from culture media of endotoxin stimulated mouse macrophages471 and their genes can be coordinately expressed after stimulation of T cells (for example, with anti-CD3), B cells, or monocytes and macrophages (for example, with lipopolysaccharide).472-476 MIP-1α gene is rapidly induced in human monocytes following adherence to endothelial cells and to other substrates.477 Monocyte chemoattractant protein 1 (MCP-1) is a monocyte chemoattractant and activating factor and is the best characterised CC chemokine, having been purified and cloned from different sources.478-480 The other CC chemokines—I-309, RANTES, and HC-14—were purified and cloned as products of activated T cells.474 ,481 ,482 Substractive hybridisation was used to find genes expressed uniquely in T cells and this led to the discovery of RANTES (Regulated on Activation, Normal T cell Expressed, and Secreted) cDNA encoding a polypeptide of 91 amino acids with an 8 kDa secreted protein. RANTES gene is expressed in IL-2 dependent T cell lines. In peripheral blood mononuclear cells low but detectable levels of RANTES transcripts can be measured in unstimulated cells and an increase in mRNA 5–7 days after antigen or stimulation with phytohaemagglutinin.481 HC-14, discovered from IFN-γ stimulated monocytes, now called MCP-2, has also been isolated from osteosarcoma cell cultures,483 which also yielded MCP-3 which has been cloned and expressed.484 ,485 MCP-4 has also been identified within a large scale sequencing and expression programme for the discovery of new chemokines.486 ,487Eotaxin is a chemokine selective for eosinophils which was discovered in BAL fluid obtained from an experimental model of allergen exposure of sensitised guinea pigs488 and subsequently shown to be present in humans.489 Another functionally similar chemokine, eotaxin-2, has recently been described.30STCP-1 is another new CC chemokine which is a chemoattractant for Th2 cells.490
In general, monocytes and tissue macrophages are a rich source of CC chemokines, usually associated with de novo synthesis. MCP-1 with MCP-2 is a major stimulated product of monocytes. Lymphocytes are sources of some CC chemokines, particularly RANTES,474 ,481 ,491I-309,474 ,492 MIP-1α,473 ,474 ,491 and MIP-1β.473 ,493 Neutrophils can produce MIP-1α.494 Eosinophils of patients with hypereosinophilic syndrome express mRNA for MIP-1α.302Epithelial cells stimulated with IL-1β or TNF-α produce RANTES,311 MCP-4,495 and eotaxin496 but not MIP-1α. MCP-1, RANTES, and eotaxin immunoreactivity is present in human airway epithelium.311 ,497 RANTES and eotaxin are produced by cultured human airway smooth muscle cells,116 and MCP-1 and RANTES by human eosinophils.498 ,499
Receptors
The chemokine receptors form a family of structurally and functionally related proteins, being members of the superfamily of heptahelical, rhodopsin-like, G protein coupled receptors. At least 10 CC chemokine receptors have been identified. These include CCR1 which binds MIP-1α, RANTES and MCP-3,500 ,501 CCR2 which binds MCP-1 and MCP-3,502 ,503 CCR3 which binds eotaxin, RANTES, MCP-3, and MCP-4,504 CCR4 which binds MCP-1, MIP-1α, and RANTES,505 ,506 and CCR5 which binds MIP-1α, MIP-1β, and RANTES.507 CCR6 is a specific receptor for a new lymphocyte directed CC chemokine called liver and activation regulated chemokine (LARC or MIP-3α),508 CCR7 is a receptor for another novel CC chemokine, EBI1 ligand chemokine (ELC or MIP-3β),509 and CCR8 is a specific receptor for the chemokine I309.510
Chemokine receptor usage by eosinophils has generated considerable interest because of the possibility of using receptor antagonists to block eosinophil influx and degranulation in asthma. CCR3 is considered to be the eotaxin receptor mediating mainly chemotaxis and has been identified as being the major CC chemokine receptor on eosinophils and basophils. An increase in CCR3 expression is observed in bronchial biopsy specimens obtained from asthmatic subjects511. A monoclonal antibody selective for CCR3 inhibits eosinophilia.512 Basophils also express CCR3 which mediate chemotaxis. However, the release responses of basophils are mediated by activation of the MCP-1 receptor (CCR2) expressed on basophils but not on eosinophils. Eosinophils also express CCR1, which is responsible for the MIP-1α and partly for the RANTES response. CCR5 is not expressed on eosinophils or basophils, but on monocytes which also express CCR1, CCR2, and CCR4. Several cytokines including IL-2, IL-4, IL-10, and IL-12 can upregulate CCR1 and CCR2 receptors in CD45RO+ blood lymphocytes associated with an increase in chemotactic activity of RANTES and MCP-1 on these cells.513 Differential chemokine receptor usage on different types of T helper cells is now also recognised. Th2 cells preferentially express CCR4 and CCR3514 ,515 while Th1 cells express CCR5.514 ,516
Effects
Chemokines may play a major part in activating migrating leucocytes and endothelial cells to increase adhesiveness and in establishing a chemotactic gradient. MIP-1α immobilised by binding to proteoglycans binds to endothelium to trigger adhesion of T cells, particularly CD8+ T cells to VCAM-1.517 It has been localised to lymph node endothelium and could act as a tethered ligand on endothelial cells and could therefore provide the required signals for activation of lymphocyte integrins for adhesion to endothelium and migration.
RANTES is a powerful eosinophil chemoattractant, being as effective as C5a and 2–3 times more potent than MIP-1α.518 ,519RANTES upregulated the expression of CD11b/CD18 on eosinophils.520 RANTES and MIP-1α induce exocytosis of eosinophil cationic protein from cytochalasin B treated cells, although RANTES is relatively weak in this effect.518 When injected into the skin of dogs RANTES induced an infiltration of eosinophils and monocytes.521 RANTES, but not MIP-1α, also elicited a respiratory burst from eosinophils.518 MCP-2, MCP-3, and MCP-4 are potent chemoattractants for eosinophils.486 ,522-524 Eotaxin and eotaxin-2 have selective chemoattractant activities for eosinophils in vitro and in vivo in the skin.30 Eotaxin also induces α4and β1 integrin expression on eosinophils.525 ,526 Cooperation between IL-5 and CC chemokines such as RANTES and eotaxin is increasingly recognised with IL-5 being essential for mobilising eosinophils from the bone marrow during allergic reactions and for local release of chemokines to induce homing and migration into tissues.32 ,152 ,165 Eotaxin also induces release of eosinophils and their progenitors from the bone marrow.33
RANTES is also a chemoattractant for memory T cells in vitro.527 Human MIP-1α and MIP-1β are chemoattractants for distinct subpopulations of lymphocytes with MIP-1α towards CD8+ and MIP-1β towards CD4+ T lymphocytes.528 RANTES attracts both phenotypes and acts on resting and activated T lymphocytes, while MIP-1α and MIP-1β are effective on anti-CD3 stimulated cells only.529 On the other hand, MIP-1β but not MIP-1α is chemotactic for resting T cells and enhances the adherence of CD8+ but not CD4+ cells to VCAM-1.530 MCP-1, MCP-2, MCP-3, and MCP-4 induce T cell migration.531 ,532NK cells migrate vigorously in response to RANTES, MIP-1α, and MCP-1.533 ,534 Human recombinant IP-10 is a chemoattractant for human monocytes and promotes T cell adhesion to endothelial cells.535 The C chemokine lymphotactin also shares T lymphocyte chemoattractant activity.536
CC chemokines are powerful stimulants of basophils. MCP-1 is as potent as C5a in stimulating exocytosis in human basophils,537-539 with release of high levels of histamine. In the presence of IL-3, IL-5, or GM-CSF there is enhanced release of histamine and production of LTC4.537 ,539RANTES and MIP-1α are less effective releasers of histamine from basophils. MIP-1α is inactive on basophils.540 RANTES is the most effective basophil chemoattractant,538 ,540 ,541 while MCP-1 is more effective as an inducer of histamine and leukotriene release.540 Eotaxin-1 and eotaxin-2 also are chemoattractant for basophils, in addition to stimulating the release of histamine and LTC4.30
CC chemokines MCP-1, RANTES, I-309, MCP-2, and MCP-3 attract monocytes in vitro,483 ,527 ,542-546 and MCP-1, MCP-2, and MCP-3 induce a selective infiltration of monocytes in animal skin.483 ,547 All CC chemokines stimulate [Ca++]i release.540 ,546 MCP-1 also induces a respiratory burst, an expression of β2integrins (CD11b/CD18 and CD11c/CD18), and the production of IL-1 and IL-6.544 ,547 ,548
Dendritic cells increased intracellular calcium release and migrated in response to MCP-3, MCP-4, MIP-1α, and MIP-5.549
Role in asthma
The potential role of chemokines in asthma is supported by observations that many cell types present in the asthmatic airway have the potential of generating chemokines, in particular monocytes/macrophages, T cells, airway smooth muscle cells, and airway epithelium. Increased levels of MCP-1, RANTES, and MIP-1α have been reported in BAL fluid from asthmatic subjects, and the eosinophil chemoattractant activity of BAL fluid of asthmatic subjects was blocked by antibodies to RANTES, MCP-3, and IL-5.155 ,550The increased levels of MCP-1 and RANTES were not confirmed in other studies551 ,552; eotaxin and RANTES may preferentially bind to components of sputum. Levels of MIP-1α, MCP-1, and RANTES were raised in BAL fluid following segmental allergen challenge.12 ,552 RANTES but not MIP-1α mRNA expression has been shown to be increased in bronchial biopsy specimens from patients with mild asthma.553 Epithelial cells, but not endothelial cells, preferentially secrete RANTES through the apical cell surface in asthma, thereby establishing a chemical gradient for chemotaxis across the epithelium.554 No differences in MIP-1α mRNA expression were observed in alveolar macrophages obtained from normal and asthmatic subjects but MIP-1α release is increased from alveolar macrophages of asthmatic patients.25Increased expression of RANTES and MCP-3 mRNA has been reported in the airway submucosa of patients with allergic and non-allergic asthma.555 Although RANTES immunostaining is present in the airway epithelium with no differences between normal and asthmatic subjects,553 MCP-1 is overexpressed in asthmatic epithelium.497 Increased plasma levels of RANTES are present during asthma exacerbations.556 Eotaxin mRNA and protein expression is increased in the airways of asthmatics, mainly in epithelium, T cells, macrophages, and eosinophils.511 ,557 ,558 In the guinea pig allergen challenge induces eotaxin expression, mainly in airway epithelium and macrophages.559 Increased expression and release of eotaxin occurs after allergen challenge in asthmatic subjects.560
Targeted disruption of eotaxin partially reduces antigen induced tissue eosinophilia in the mouse.156 An anti-MCP-3 antibody inhibits allergen induced eosinophilic inflammation in the mouse.561 Met-RANTES is an antagonist of eosinophil functions following stimulation with RANTES, MCP-3, and eotaxin, antagonising effects mediated through CCR1 rather than CCR3.562 The availability of specific CC chemokine receptor antagonists, particularly of the CCR3 receptor, will make it possible to examine the contribution of these chemokines in allergic inflammation and asthma.
CXC CHEMOKINES
Synthesis and release
IL-8 (also referred to as neutrophil activating protein 1, NAP-1) has major actions as a neutrophil chemoattractant and activator. Other CXC chemokines similar to IL-8 include neutrophil activating protein 2 (NAP-2)563 arising from N-terminal processing of platelet basic protein, GRO-α, GRO-β, and GRO-γ,564 ,565epithelial cell derived neutrophil activating protein (ENA-78),566 and granulocyte chemotactic protein 2 (GCP-2).567 A secreted protein produced by lipopolysaccharide stimulated murine macrophages called macrophage inflammatory protein 2 (MIP-2) is a chemoattractant for human neutrophils and is closely related to GRO.568 In general, monocytes and tissue macrophages are a rich source of CXC chemokines, usually associated with de novo synthesis. Monocytes respond to a large variety of pro-inflammatory agents including IL-1β, TNF, GM-CSF, IL-3, lipopolysaccharide, immune complexes to release IL-8. IL-8 has also been induced following adherence of monocytes to plastic and by changes in ambient oxygen.569 ,570 Eosinophils also release IL-8 after stimulation with calcium ionophore A23187, but not with TNF-α or IL-1β.571 Airway epithelial cells and airway smooth muscle cells stimulated with IL-1β or TNF-α produce IL-8.117 ,572-575 IL-8 expression by epithelial cells is increased by respiratory syncytial virus infections576 and on exposure to neutrophil elastase.577
Several transcriptional regulatory elements can bind to the region preceding the first exon including NF-κB, NF-IL-6, AP-1, glucocorticoid element, and an octamer binding motif.578NF-IL-6 and NF-κB-like factors may act ascis-acting elements in IL-8 mRNA expression.579 IL-8 mRNA expression after stimulation with IL-1β or TNF-α is rapid and results, at least partly, from transcriptional activation as shown by nuclear run-on assays.575 ,580-582 A secondary phase of IL-8 mRNA expression following an early rapid increase induced by IL-1 is observed with cultured human airway epithelial cells.
Receptors
At least four CXC receptors have been identified. For IL-8 there are high affinity (CXCR1) and low affinity (CXCR2) receptors which are mainly expressed on neutrophils.583 ,584 CXCR3 is the receptor for IP-10, MIG, and I-TAC and is found mainly on activated T cells, particularly Th1 cells and natural killer cells,585 ,586 while CXCR4 is the receptor for SDF-1 and is localised to dendritic cells.549 These receptors are a family of structurally and functionally related proteins, being members of the superfamily of heptahelical, rhodopsin-like, G protein coupled receptors. IL-8 on neutrophils also induces G protein activation.587 CXCR1 is specific for IL-8 and other CXC chemokines do not bind to it.583 Its sequence is 77% identical to that of CXCR2.583 CXCR2 can be activated by CXC chemokines containing the sequence Glu-Leu-Arg in the N-terminal domain, including IL-8, GRO, and NAP-2, but not by CC chemokines.584 ,588
Effects
IL-8 is a mainly neutrophil chemoattractant and activator which induces shape change, a transient rise in [Ca++]i, exocytosis with release of enzymes and proteins from intracellular storage organelles, and respiratory burst through activation of NADPH oxidase.589 It also upregulates the expression of two integrins (CD11b/CD18 and CD11c/CD18) during exocytosis of specific granules.590 ,591 IL-8 activates neutrophil 5-lipoxygenase with the formation of LTB4 and 5-HETE,592 and also induces the production of PAF.593
IL-8 can also induce an increase in [Ca++]i, shape change, and release of eosinophil peroxidase from the eosinophils of patients with hypereosinophilic syndrome.594 It has a small amount of chemotactic activity for both CD4+ and CD8+ T lymphocytes595 but intradermal injection of IL-8 in humans does not attract lymphocytes.596 ,597 IL-8 induces the release of histamine598 ,599 and cys-LTs598from human blood basophils with enhanced release with IL-3, IL-5, or GM-CSF pretreatment.600 It induces release of a small amount of [Ca++]i and respiratory burst.601
Role in asthma
Enhanced co-expression of IL-8 and GM-CSF in bronchial epithelial cells of patients with asthma has been reported.602 Free IL-8 has been detected in the serum and bronchial tissue of subjects with severe atopic asthma but not in normal subjects or those with mild atopic asthma, suggesting that IL-8 may be a marker of severe asthma. IL-8 was also found to be complexed with IgA, levels of which were raised in bronchial tissue in asthma.603 However, in segmental local challenge studies of patients with allergic asthma IL-8 increased levels correlated with neutrophil influx,604indicating that IL-8 may be responsible for neutrophil chemotaxis. Enhanced release of IL-8 has been demonstrated from alveolar macrophages obtained from patients with mild asthma compared with those from normal subjects.605 In patients with mild asthma there is no increase in IL-8 levels in induced sputum, in contrast to the markedly raised levels in patients with COPD and bronchiectasis.606 ,607 Increased levels of IL-8 have been measured in the BAL fluid of patients with asthma and bronchitis.607
IL-8 possesses chemotactic activity for primed eosinophils.608 Human IL-8 is able to induce accumulation of eosinophils in guinea pig skin609 and an anti-IL-8 antibody inhibited IL-1 induced eosinophil accumulation in rat skin.298 Local instillation of rh-IL-8 to the nose caused an extravascular accumulation of eosinophils in the nasal mucosa of atopic but not in normal subjects.610
Growth factors
PLATELET-DERIVED GROWTH FACTOR (PDGF)
Synthesis and release
PDGF is released from many different cells in the airways and consists of two peptide chains so that AA, BB, or AB dimers may be secreted by different cells. Both PDGF-A and PDGF-B chains are synthesised as high molecular mass precursors which are then extensively processed before secretion.611 ,612Post-translational glycosylation and proteolytic cleavage611 ,613 ,614 both contribute to the heterogeneity of the apparent molecular mass of mature proteins. Most of the PDGF present in human platelets (from which PDGF was originally isolated) has been identified as AB dimer, although BB and AA dimers also exist.615 ,616 PDGF-like activity in the conditioned media of various cells such as those derived from smooth muscle consists predominantly of the AA dimer.617 The sources of PDGF include platelets, macrophages, endothelial cells, fibroblasts, airway epithelial cells, and vascular smooth muscle cells. Various stimuli such as IFN-γ on alveolar macrophages, hypoxia, basic FGF, mechanical stress on endothelial cells, serum, TNF-α, IL-1, and TGF-β on fibroblasts can induce PDGF release.
Receptors
The PDGF receptors belong to a family of closely related receptor proteins that include the receptor for monocyte-colony stimulating factor and the c-kit receptor.618 PDGF exerts its actions through a family of at least two classes of PDGF receptors, α and β.619-621 They are single transmembrane glycoproteins with an intracellular tyrosine kinase domain.622 Binding of PDGF dimers induces receptor dimerisation with three possible configurations (αα, αβ, ββ). The PDGF receptor α subunit binds both PDGF-A and -B chains, whereas the β receptor subunit binds only PDGF-B chains. Thus, PDGF-AA binds only to PDGF receptor αα dimers, PDGF-AB to receptor αα and αβ dimers, and PDGF-BB to all three configurations.623 ,624 These receptors are widely distributed on cells of mesenchymal origin including fibroblasts and smooth muscle cells. Because of their critical role in cell growth, expression of PDGF receptors is usually tightly controlled. However, they can be regulated by TGF-β which can increase the expression of PDGF receptor α on human skin fibroblasts625 ,626 or by basic fibroblast growth factor which induces the expression of PDGF α receptor on bronchial smooth muscle.627
Effects
PDGF is a major mitogen whose main regulatory role is directed at the cell cycle, acting as a competence factor triggering early events of the cell cycle leading to DNA synthesis and mitosis.628PDGF induces the expression of competence genes including the proto-oncognes c-myc,c-fos, andc-jun.629 ,630 PDGF may activate fibroblasts to proliferate and secrete collagen631 and may also stimulate proliferation of airway smooth muscle48 which is mediated via the receptor.632 PDGF is a chemoattractant for connective tissue cells633 ,634 and can stimulate fibroblasts to contract collagen lattices.635
Role in asthma
Levels of PDGF-AA, -AB, and -BB are not raised in asthma and immunohistochemistry for PDGF-AA and -BB, and for PDGFR-α and PDGFR-β are not increased.636 Expression of PDGF in the bronchial mucosa of asthmatic patients with thickened lamina reticularis is not increased compared with normal subjects.637 Similar results were obtained in another study in which PDGF was immunolocalised to tissue macrophages.638 One source of PDGF B-chain is the eosinophil in nasal polyps or bronchial biopsy specimens from patients with asthma.639 This has raised the possibility that eosinophils, together with their ability to express TGF-β, are involved in airway remodelling in patients with asthma.
TRANSFORMING GROWTH FACTOR β(TGF-β)
Synthesis and release
Monocytes express TGF-β mRNA constitutively but only release the protein when activated.640 ,641 Pulmonary macrophages may store large amounts of TGF-β during pulmonary inflammation.642 Lung fibroblasts themselves may be a source of TGF-β,643 but it is also secreted by inflammatory cells including eosinophils,644 ,645neutrophils,646 and airway smooth muscle cells, and structural cells such as epithelial cells.647 Mast cells may also be another source.254 TGF-β is present in the epithelial lining fluid of the normal lower respiratory tract.648 TGF-β mRNA and protein have been found to be abundantly expressed in human lung, with TGF-β1 precursor being immunolocalised throughout the airway wall including the epithelium and in alveolar macrophages, and the mature protein localised mainly within the connective tissue of the airway wall.649
Receptors
TGF-β receptor exists in three forms: a high affinity type I and II, and a low affinity type III.650 The high affinity receptors are serine/threonine kinases related to the activin receptor and are thought to associate to mediate signal transduction probably through serine/threonine phosphorylation. The type II receptor includes β-glycan and endoglin and does not transduce signals but may concentrate TGF-β on the cell surface and present the ligand to the other receptors.
Effects
TGF-β comprises a family of growth modulating cytokines with an important influence on the turnover of matrix proteins.651They may either inhibit or stimulate proliferation of fibroblasts depending on the presence of other cytokines. TGF-β induces the transcription of fibronectin which can function as a chemotactic agent and growth factor for human fibroblasts.652 ,653 It may also be involved in the repair process of airway epithelial damage characteristic of asthma, since TGF-β is a potent inducer of differentiation for normal epithelial cells.654 TGF-β is a potent profibrotic cytokine that stimulates fibroblasts to promote the synthesis and secretion of many proteins of the extracellular matrix.655 It is also a potent chemoattractant for many cell types including monocytes, fibroblasts, and mast cells.656 ,657 TGF-β activates monocytes to produce other cytokines such as TNF-α, TGF-α, TGF-β, PDGF-B, and IL-1. TGF-β has complex actions on the immune system. In general, TGF-β1 inhibits both T and B cells. Thus, TGF-β inhibits IL-1 dependent lymphocyte proliferation287 and blocks IL-2 mediated induction of IL-2 receptors on T cells.658 TGF-β1 can both decrease and increase airway smooth muscle proliferation.659 ,660
Role in asthma
Expression for both forms of TGF-β1 is reported to be similar in lungs from normal and asthmatic subjects.649However, a greater number of airway mucosal eosinophils expressing TGFβ1 mRNA and protein has been reported, correlating with the severity of asthma and the degree of subepithelial fibrosis.661 This was not confirmed in other studies of asthmatic subjects in whom an increase in the lamina reticularis of the basement membrane was reported.637 ,662TGF-β1 immunoreactivity has been observed in the epithelium and submucosal cells such as eosinophils and fibroblasts, with a greater expression in biopsy specimens from patients with chronic bronchitis than those with asthma.663 Release of TGF-β1 into the BAL fluid has been observed following segmental allergen challenge.664 The possibility remains that TGF-β (together with PDGF) may be involved in the remodelling process of asthma, although it may also participate in modulating the T cell response.
FIBROBLAST GROWTH FACTOR (FGF)
FGF represents a family of heparin binding growth factors consisting of seven polypeptides including acidic (aFGF) and basic FGF (bFGF).665 aFGF and bFGF are potent modulators of cell proliferation, motility, and differentiation. They are found associated with extracellular matrix. A major role has been proposed for FGF in the induction of angiogenesis.666 bFGF induces an invasive phenotype in cultured endothelial cells, enabling them to penetrate the basement membrane in vitro.667 It induces increased production of proteolytic enzymes, plasminogen activators, and collagenase.667-669 bFGF binds to heparan sulphate proteoglycans in basement membranes in vivo.670 In the human adult lung bFGF has been localised to vascular smooth muscle and in endothelial cells of blood vessels of the lungs.671 It has also been detected in high levels in epithelial cells of the trachea and bronchi. bFGF increases the expression of PDGF-Rα in human airway smooth muscle and therefore indirectly stimulates proliferation.627
EPIDERMAL GROWTH FACTOR (EGF)
EGF and TGF-α, which do not bind heparin, also stimulate angiogenesis.672 EGF expression is increased in the epithelium of bronchitic subjects and the submucosa of patients with asthma.663 It increases airway smooth muscle proliferation49 and endothelin-1 potentiates EGF induced airway smooth muscle proliferation.673 Since an increased number of blood vessels has been described in asthmatic airways,9 these growth factors may be implicated. EGF expression is reported to be increased in the epithelium of patients with chronic bronchitis and in the submucosa of asthmatic patients.663
INSULIN-LIKE GROWTH FACTOR (IGF)
IGF is produced by airway epithelial cells.674 It is a potent mitogen for airway smooth muscle proliferation675and appears to mediate the proliferative effect of LTD4 on airway smooth muscle proliferation.676 IGF is a potent mitogen and activates MAP kinases in airway smooth muscle.677There is no difference in the expression of IGF in the airways submucosa of patients with asthma who have a thickened reticularis lamina of their basement membrane.678 Inhaled corticosteroid treatment is associated with a reduction in the expression of IGF in the airways of patients with asthma, associated with a reduced lamina reticularis.679
Conclusion
The field of cytokine biology is moving very rapidly with the discovery of new molecules and of the basic mechanisms by which these cytokines act. Our review details the involvement of many cytokines in asthma and their contribution to the pathogenesis of asthma appears to be very diverse. Despite a large amount of information on the biological activities of cytokines and on their potential involvement in various aspects of asthmatic inflammation, much remains to be learnt about cytokines in day-to-day asthma. While current interest is focusing to a large extent on the immunoregulatory polarisation of T cells into Th2 cells as a cause of asthma, little is known about the stratification of the cytokines into the clinical stages of asthma. What are the profiles of the cytokine network at the “pre-asthma” stage prior to the onset of symptoms? When the clinical disease is established, do certain cytokines (or cytokine networks) become more prominent? In the presence of established airway wall remodelling, cytokines with proliferative activities on resident cells may be more important. This also raises the possibility that the severity of asthma may be related to differential cytokine expression.
An understanding of cytokine biology has led to the identification of several targets that could be developed as potential treatment for asthma.680 Indeed, there is currently a large drive to test several inhibitors of cytokine effects in asthma, such as inhibitors of IL-5 which is a cytokine central to the development of tissue eosinophilic inflammation. Clinical studies of an antibody to IL-5 are currently underway. However, it is unclear to what extent blocking the effects of one single cytokine out of all the cytokines that are potentially involved will lead to important therapeutic effects. For example, in terms of eosinophilic inflammation, not only IL-5 but also the eosinophil selective chemokines may contribute. Testing such inhibitors will also address the question of, in this instance, the importance of eosinophils in causing clinical asthma, as distinct from the studies in animals. A study of the actions of these cytokines may provide other ways of blocking the effects of groups of cytokines. Corticosteroids may have beneficial effects in asthma by their ability to block the generation of several pro-inflammatory cytokines. Blocking the effects at an early stage of asthma may control or cause remission in the disease. Therefore, not only will greater understanding of asthma pathogenesis be achieved by studying cytokine biology, but also the potential for newer more effective treatments, perhaps even a cure.
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵