Article Text

Abstract
BACKGROUND Pollens are important triggers for asthma but the mechanism of sensitisation to their proteins remains poorly understood. The intrinsic protease activity of some allergens may contribute to sensitisation by disrupting the integrity of the airway epithelial barrier. Pollens release a variety of enzymes, including proteases, upon hydration. The hypothesis that such enzymes might be able to damage airway epithelial cells was therefore tested.
METHODS Diffusates from pollens ofLolium perenne (ryegrass), Poa pratensis(Kentucky bluegrass), Acacia longifolia (Sydney golden wattle), or Casuarina distyla (she-oak) were incubated with mouse tracheal epithelial cells in culture and cellular detachment was quantified using a methylene blue dye binding assay.
RESULTS Diffusates prepared using 100 mg/ml of pollen caused detachment of 30–90% of airway epithelial cells in separate experiments. Within each experiment comparable detachment was observed with all diffusates tested, although total protein in the diffusates varied markedly between species. Viability of the cells recovered after exposure to Acacia diffusate was higher than after detachment by exposure to Loliumdiffusate. Cellular detachment by all of the diffusates could be almost completely inhibited by addition of 10% serum. Aprotinin, an inhibitor of serine proteases, partially blocked activity in diffusates ofLolium pollen but not of Acacia pollen. In contrast, α1-protease inhibitor and secretory leucocyte protease inhibitor (SLPI) were not able to block the activity of either diffusate at concentrations which inhibited cellular detachment by trypsin.
CONCLUSIONS Proteases released by pollens are able to cause detachment of airway epithelial cells from their substratum in vitro and may not be effectively inhibited by endogenous antiproteases.
- pollens
- allergens
- proteases
- tracheal epithelium
Statistics from Altmetric.com
Clinical asthma is usually related to underlying IgE mediated allergic inflammation.1 However, the mechanisms of sensitisation to inhaled allergens remain incompletely understood. Tight junctions between airway epithelial cells apparently constitute a physical barrier between inhaled antigens and the immune system. Experimental evidence suggests that, if this barrier is breached at the time of initial exposure to antigen by concurrent infection or exposure to environmental pollutants, an IgE response is more likely to be induced.2 This may be a result of facilitated access of allergenic protein components to the subepithelial antigen presenting dendritic cells.3 In this context it is therefore noteworthy that at least some allergens have intrinsic enzymatic activity—for example, major house dust mite4 and cockroach5 allergens exhibit protease activity. Exposure to such proteases in vitro causes epithelial detachment and increases the permeability of epithelial sheets and cultured monolayers of airway epithelium.6 These observations have led to the suggestion that disruption of intercellular junctions by the enzymatic activity of inhaled allergens might cause loss of integrity of the airway epithelial barrier in vivo and thus promote sensitisation to these antigens.4 ,6
Airborne allergens derived from plant pollens are major triggers for asthma.7 However, information about whether these might function as proteases is lacking.8 Pollens do contain a variety of enzymes including proteases, and very high local concentrations of pollen solutes are likely to be achieved when pollen grains are deposited on mucosal surfaces because 20–25% of the pollen mass is released rapidly upon hydration.9 ,10 We hypothesised that proteases released by inhaled pollens might be able to damage the airway epithelium. To test this hypothesis, we investigated whether diffusates of potentially allergenic pollens were able to cause detachment of murine airway epithelial cells from their substratum in vitro.
Methods
PREPARATION OF POLLEN DIFFUSATES
Pollens of the allergenic species Lolium perenne(ryegrass), Poa pratensis (Kentucky bluegrass), andAcacia longifolia (Sydney golden wattle) were obtained from Bayer Australia, Sydney. Pollen of Casuarina distyla(she-oak) was collected in the Royal National Park, Sydney (with the assistance of Dr Peter Stricker, School of Biological Science, The University of New South Wales). Diffusates of the pollens were prepared under physiological conditions by thoroughly mixing the dry pollen with basal medium MCDB 201 (Sigma, St Louis, Missouri, USA) at ratios of 10–100 mg pollen per ml. The suspension was then incubated at 4°C for 30 minutes without further agitation, followed by centrifugation at 16 000g for eight minutes to pellet the pollen grains, and the supernatants were used immediately. The protein content of an aliquot of each diffusate was measured using a dye binding assay (Bio-Rad, Sydney) with bovine serum albumin (0.1–1.6 mg/ml) as the standard.
CULTURE OF MURINE AIRWAY EPITHELIAL CELLS
Mouse airway epithelial cells were cultured from explants of tracheal tissue in serum-free medium as previously described.11 Briefly, fragments of tracheal tissue from 8–10 week old specific pathogen-free BALB/c mice were cultured at 37°C in an atmosphere of 2% CO2 in air in serum-free medium MCDB 201 with appropriate supplements including bovine serum albumin, lipids, transferrin, epidermal growth factor, and insulin-like growth factor 1. Primary cultures were established weekly using tracheas from three mice. As reported previously,11 each culture yielded approximately 2 × 106 cells at the second passage, thus providing sufficient cells for a single assay.
CELLULAR DETACHMENT ASSAY
To assess the ability of pollen diffusates to detach airway epithelial cells from their substratum, cells at the second passage were seeded in growth factor-deficient medium at 1.25 × 104 per well of a collagen coated 96-well culture plate (Nunc, Roskilde, Denmark) for two hours, by which time ⩾95% of the cells had attached to the substratum. The medium was then removed and diffusates were added to the wells in quadruplicate. Following incubation for four hours the wells were gently washed and fixed overnight with 10% phosphate buffered formalin. The proportion of cells remaining attached was assessed using a methylene blue dye binding assay.12 Briefly, cells were stained with 1% methylene blue in 0.01 M borate buffer, pH 8.5, for 30 minutes and washed three times in buffer. The bound dye was subsequently eluted with a 1:1 mixture of absolute ethanol and 0.1 M hydrochloric acid and absorbances of individual wells were read at 650 nm. Percentage detachment was calculated relative to the absorbance of wells incubated in basal medium. Cells incubated with 10 μg/ml of trypsin served as a positive control for the cellular detachment.
ASSESSMENT OF VIABILITY OF DETACHED CELLS
The viability of cells detached by incubation with diffusates ofAcacia and Lolium was assessed using trypan blue exclusion, compared with cells detached by 10 μg/ml of trypsin as a positive control. Because trypan blue exclusion underestimates the proportion of non-viable cells, the ability of the detached cells to survive in subculture was also assessed. For these experiments, diffusates were incubated for one hour with airway epithelial cells which had previously been seeded at 5 × 105 cells per well of a six-well culture plate. The detached cells were re-seeded in a 24-well culture plate and after 24 hours of incubation the number of attached cells was determined.
EFFECT OF PROTEASE INHIBITORS
Preliminary characterisation of the cellular detachment activity included non-selective inhibition of enzyme activity by the addition of newborn calf serum (Trace Biosciences, Sydney) to a final concentration of 10% as well as selective inhibition of proteases.13For the latter experiments we compared the non-cytotoxic inhibitors aprotinin (Bayer) (an inhibitor of serine proteases), α1-proteinase inhibitor (α1-PI) (an endogenous antiprotease) (both from Sigma), and secretory leucocyte proteinase inhibitor (SLPI) (an endogenous inhibitor of trypsin-like enzymes) (R&D Systems, Minneapolis, Minnesota, USA).
ANALYSIS OF DATA
To assess the statistical significance of any inhibitory effect, analysis of variance was performed for each assay followed by Newman-Keuls test for comparison between groups.
Results
Pollen diffusates of all species tested caused detachment of airway epithelial cells from their substratum. In separate experiments using different primary cultures of tracheal epithelial cells as targets, maximum cellular detachment by diffusates prepared with 100 mg pollen per ml of basal medium varied between 30% and 90% (results from at least three experiments for each species of pollen). However, in a given experiment detachment by different pollens was of comparable magnitude even though the total protein concentration in diffusates prepared with an equal dry weight of pollen differed markedly between species (table 1). Detachment activity was related to the concentration of solute and diminished with serial dilution of the diffusate (fig 1).
Cellular detachment activity and concentration of protein in diffusates of various pollens
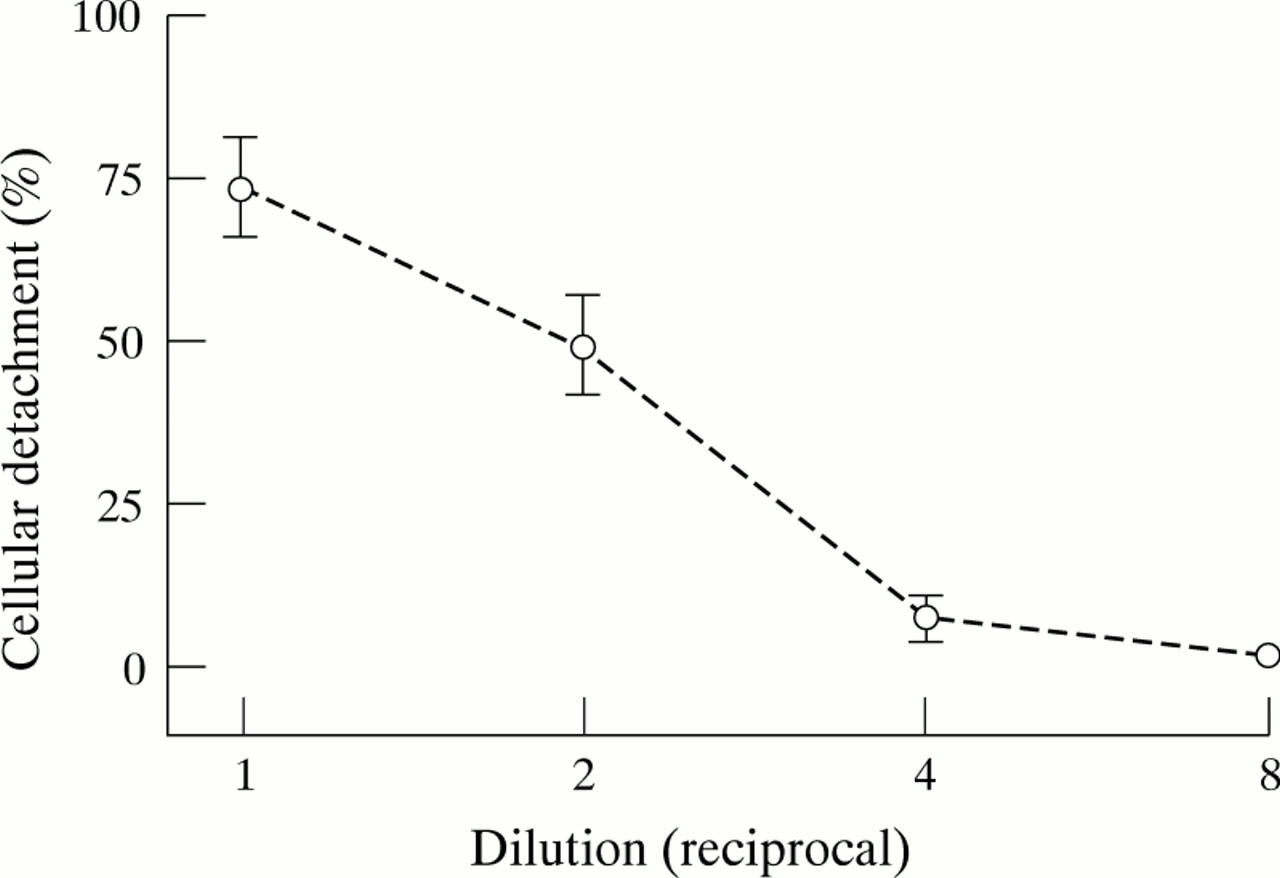
Effect of dilution of Lolium diffusate prepared using 100 mg/ml pollen on percentage detachment of murine airway epithelial cells from a collagen substratum. Data are mean (SE) of three separate assays.
Incubation with a 30 mg/ml diffusate of Acacia, which typically caused detachment of 30–60% of cells, yielded a population with approximately 50% viability as assessed by trypan blue exclusion. Upon re-seeding in culture approximately 20% of the detached cells re-attached and remained viable after a further 24 hours. In contrast, incubation with a 100 mg/ml diffusate of Lolium caused detachment of a similar proportion of the cells but all of these were non-viable, suggesting that this diffusate was cytotoxic. In comparison, incubation with 10 μg/ml trypsin detached all of the cells with ⩾90% viability.
Cellular detachment by trypsin was substantially inhibited by 1000 U/ml aprotinin and wholly inhibited by 10% serum. Detachment activity in diffusates of Lolium pollen was partially blocked by aprotinin and almost completely blocked by serum in three separate assays (fig 2A). However, aprotinin had no effect upon the activity in Acacia pollen although serum inhibited cellular detachment by this diffusate (not shown).

{kind=link}
{kind=link}
Effect of protease inhibitors on cellular detachment by trypsin or by pollen diffusates. (A) Both aprotinin and newborn calf serum caused significant inhibition of detachment by trypsin (*p<0.05; **p<0.01) and by Lolium diffusate (#p<0.05; ##p<0.01). (B) α1-PI and SLPI completely inhibited cellular detachment by trypsin (**p<0.01) but there was no statistically significant inhibitory effect of α1-PI or SLPI on Lolium or Acacia diffusate. In each case data are mean (SE) of replicates from a single assay.
Cellular detachment by trypsin was also completely blocked by incubation with 200 μg/ml α1-PI or 10 μg/ml SLPI. Although α1-PI caused modest inhibition of the activity of Lolium diffusate (fig 2B), this was not statistically significant in three of four assays. SLPI did not block the activity ofLolium diffusate (fig 2B), nor did it potentiate the inhibition by α1-PI (not shown). Neither protease inhibitor had any effect upon cellular detachment byAcacia diffusate (fig 2B).
Discussion
These experiments show that solutes released by a variety of allergenic pollens are able to cause detachment of murine airway epithelial cells from their substratum in vitro. We deliberately used an approach which recovered soluble components released after hydration without mechanical disruption because most of the solutes that are likely to be pathophysiologically relevant are released from the outer layers of the pollen grain within minutes of hydration and these constitute more than two thirds of the total extractable solutes.9 ,10 Prolonged extraction as is usually employed for studies of pollen solutes—for example, in hypotonic solution rather than a buffer of physiological osmolality, with stirring or grinding—liberates proteins from the inner layers of the pollen grain which might be less readily available to cause tissue injury.
Cellular detachment activity in the pollen diffusates was substantially inhibited by addition of serum and, at least in the case ofAcacia pollen diffusate, permitted the detachment of a proportion of viable cells. It is therefore likely that activity is related to the action of enzymes in the pollen diffusates, most probably proteases. Nevertheless, pollen diffusates contain a variety of solutes and we cannot exclude a role for other components in causing detachment of airway epithelial cells. Indeed, non-enzymatic components probably account for the cytotoxicity that was observed. Activity was not simply a function of total solute release, as comparable cellular detachment was observed using diffusates which differed markedly in their total protein content. In this context it is of interest that the concentration of protein was lowest in diffusates of Acacia longifolia, as this is the only insect pollinated species among those tested. Whether this is also a characteristic of other insect borne pollens is as yet unknown.
Preliminary characterisation indicated that a component of the detachment activity in Lolium perenne diffusates was inhibited by aprotinin, suggesting the presence of a trypsin-like or chymotrypsin-like serine protease. Whether such a protease would be active in vivo is uncertain as the test system contained none of the endogenous antiprotease activities that might inhibit its activity. However, the diffusate could not be reproducibly inhibited by α1-PI which is a major endogenous antiprotease nor was it blocked by SLPI which is an inhibitor of trypsin-like proteases produced by airway epithelial cells that may complement the activity of endogenous α1-PI.14 Moreover, epithelial detachment by Acacia diffusates was not blocked by any of the selective inhibitors tested. These findings are of interest in the context of the recent isolation of novel serine proteases from the pollens of mesquite and ragweed.15 ,16 Although their ability to injure airway epithelial cells has not been studied, the peptidase recovered from mesquite pollen was not inhibited by any of the known endogenous mammalian antiproteases15 while the peptidase from ragweed could cleave and inactivate human α1-PI.16 Taken together with our observations of the relative lack of effect of endogenous antiproteases, these findings reinforce the notion that pollen derived proteases have significant potential to cause airway epithelial injury.
Using high concentrations of dithiothreitol activated Der p 1 protein, Herbert et al 6 demonstrated proteolytic disruption of canine airway epithelial cells and suggested that tissue damage in vivo could potentiate sensitisation to this antigen. The extent to which comparable injury by pollen derived enzymes might be relevant to allergen sensitisation is difficult to ascertain. However, very high local concentrations of pollen solutes (up to 50 mg/ml) may be achieved when pollen grains are deposited on mucosal surfaces.9 In any case, epithelial denudation is not necessarily involved in increasing epithelial permeability because proteolytic enzymes may cause disruption of tight junction associated proteins.17 This could be tested using cells grown in culture well inserts,18 although we have encountered technical difficulties relating to the use of mouse tracheal epithelial cells.
If pollen proteases do contribute to epithelial injury and sensitisation, it is unlikely that this occurs in the lower respiratory tract as most intact pollen grains are likely to be trapped within the nasopharynx because of their size. Rupture of pollen grains can release granules of respirable dimensions which might facilitate access to more distal airways,19 although the enzymatic activity associated with such granules is undefined. Nevertheless, upper respiratory tract exposure would certainly be sufficient to cause sensitisation, and disruption of the epithelial barrier by proteases in pollens might promote sensitisation even to those pollen antigens which do not have intrinsic enzymatic activity.