Article Text

Abstract
BACKGROUND Acute exposure of healthy subjects to swine house dust causes increased bronchial responsiveness to methacholine but no acute bronchoconstriction. The role of cysteinyl leukotrienes and mast cells in increased bronchial responsiveness is unclear.
METHODS Ten non-asthmatic subjects were exposed to swine dust for three hours while weighing pigs in a piggery. Urine was collected prior to and for up to 12 hours after entering the piggery and at the same times five days before and the day after exposure. As indices of whole body leukotriene production and mast cell activation, urinary levels of leukotriene E4(LTE4) and 9α,11β-PGF2, the earliest appearing urinary metabolite of prostaglandin D2(PGD2), were measured. Bronchial responsiveness to methacholine was determined five days before and the day after the exposure.
RESULTS Methacholine PD20FEV1 decreased from 1.32 mg (95% CI 0.22 to 10.25) before exposure to 0.38 mg (95% CI 0.11 to 1.3) after exposure (p<0.01). Associated with the increase in bronchial responsiveness there was a significant mean difference between post- and pre-challenge levels of LTE4 (difference 38.5 ng/mmol creatinine (95% CI 17.2 to 59.8); p<0.01) and 9α,11β-PGF2 (difference 69 ng/mmol creatinine (95% CI 3.7 to 134.3); p<0.05) on the day of exposure to swine dust. Swine dust exposure induced a 24-fold increase in the total cell number and a 12-fold increase in IL-8 levels in the nasal lavage fluid. The levels of LTB4 and LTE4 in nasal lavage fluid following exposure also increased 5.5-fold and 2-fold, respectively.
CONCLUSIONS The findings of this study indicate that cysteinyl leukotrienes and other mast cell mediators contribute to the development of increased bronchial responsiveness following inhalation of organic swine dust.
- swine dust
- leukotrienes
- prostaglandin D2
Statistics from Altmetric.com
Cysteinyl leukotrienes (LTC4, LTD4, and LTE4) mediate bronchoconstriction evoked in asthmatic subjects by different environmental trigger factors such as allergen, exercise or dry cold air as well as by aspirin in aspirin intolerant individuals.1 The role of leukotrienes in the development or expression of increased bronchial responsiveness is less clear. It has been suggested that decreased bronchial responsiveness occurs during treatment with antileukotrienes.1 However, responsiveness to histamine or methacholine following allergen challenge has produced variable results with regard to the effectiveness of antileukotrienes.2 3
In an attempt to provide more information on the possible role of leukotrienes in increasing bronchial responsiveness we have used the swine house dust exposure model. A brief three hour stay in a piggery induces an inflammatory reaction in the airways of healthy volunteers, as documented by measurement of markers of inflammation in nasal and bronchoalveolar lavage (BAL) fluid and in blood.4 5Interestingly, there is no acute bronchoconstriction but a conspicuous increase in bronchial responsiveness to methacholine after exposure in individuals not previously exposed to swine dust.6 The model thus provides the opportunity of studying the development of increased bronchial responsiveness without having to consider the confounding effects of ongoing or prior bronchoconstriction.
The primary aim of this study was to determine whether evidence of activation of the leukotriene pathway could be obtained following exposure to swine dust, and if such a change was associated with the development of increased bronchial responsiveness. The end metabolite of cysteinyl leukotrienes, LTE4, has previously been established as a reliable marker of leukotriene generation in the lung, and urinary LTE4 levels are increased, for example, following allergen bronchoprovocation and exacerbations of asthma.7 8
The cellular response to inhaled swine dust is dominated by neutrophils as assessed by BAL.9 Leukotriene B4(LTB4) is the predominant leukotriene generated in neutrophils. There are currently no methods available to study urinary excretion of LTB4 or its metabolites so its concentration was determined in nasal lavage fluid. Although the chemoattractant LTB4 is devoid of bronchoconstrictive properties in asthmatic subjects,10 it may nevertheless be involved in cell recruitment preceding the appearance of increased bronchial responsiveness. Inhalation of LTB4 is known to cause cellular changes in the airways and blood of healthy subjects11 and asthmatic subjects.10
Finally, previous studies have suggested a relationship between increased bronchial responsiveness and mast cell mediator release.12-14 We therefore also investigated whether or not exposure to swine dust was associated with evidence of mast cell activation in terms of increased urinary excretion of 9α,11β-PGF2, the earliest metabolite of the mast cell product prostaglandin D2 (PGD2) to appear in the urine.15
Methods
STUDY DESIGN
Ten healthy non-smoking subjects (two men) of mean age 39 years (range 26–60) participated in the study. The subjects were non-atopic and non-asthmatic as determined by history and a questionnaire. All participants gave their informed consent and the study was approved by the ethics committee at the Karolinska Institutet.
The subjects were exposed to swine dust for three hours (between 08.00 and 11.00 hours) while weighing pigs in a piggery containing approximately 900 pigs. Bronchial responsiveness to methacholine was determined five days before exposure (pre-exposure day) and the day after the exposure (post-exposure day).
Urine was collected at roughly hourly intervals prior to and for up to 12 hours after entering the piggery and at the same time on the pre- and post-exposure days—that is, when methacholine challenges were performed. All subjects underwent nasal lavage on the exposure day prior to entering the piggery and again seven hours after the start of exposure.
PULMONARY FUNCTION MEASUREMENTS AND METHACHOLINE BRONCHOPROVOCATION
Forced expiratory volume in one second (FEV1) and vital capacity (VC) were measured according to the American Thoracic Society (ATS) criteria with a wedge spirometer (Vitalograph, Buckingham, UK). The method for the bronchial provocation has been designed for use in normal volunteers and has been thoroughly validated. The methodology is described in detail elsewhere.16 Bronchial responsiveness was expressed as the dose of methacholine that caused a 20% reduction in FEV1compared with the value obtained after inhalation of diluent (PD20). One of the 10 participating subjects had a pre-exposure PD20 value below the fifth percentile (0.21 mg, n = 82) of the reference values established for this method.
NASAL LAVAGE
Nasal lavage was performed as described previously.4
ENZYME IMMUNOASSAY (EIA) PROCEDURES
Analysis of 9α,11β-PGF2 and LTE4 in the urine and LTE4 and LTB4 in the lavage fluid was performed with polyclonal antisera and acetylcholinesterase linked tracer as described previously.15 17 Urinary data are expressed as ng compound per mmol filtered creatinine.17The concentration of LTB4 and LTE4 in the nasal lavage fluid is reported as pg per ml recovered fluid.
Automation of the EIA procedure was performed with a pipetting robot, MultiProbe 104 (Canberra Packard, Merideen, Connecticut, USA). The detection limit was approximately 8 pg/ml for all compounds.
Interleukin 8 (IL-8) was measured by ELISA using a commercial immunoassay (Quantikine, R&D Systems Europe, Abingdon, Oxon, UK) with a lower detection limit of 31.3 pg/ml.4
SWINE DUST EXPOSURE
The subjects carried personal exposure dosimeters as previously described.4
DRUGS AND MATERIALS
Synthetic 9α,11β-PGF2, LTE4, and LTB4 were obtained from Cayman Chemical Company, Ann Arbour, Michigan, USA and Cascade Biochemicals, Reading, UK. Rabbit polyclonal antisera against 9α,11β-PGF2, LTE4, and LTB4 acetylcholinesterase linked 9α,11β-PGF2, LTB4 and LTE4tracer, mouse monoclonal anti-rabbit IgG, and Ellman’s reagent were all purchased from Cayman Chemical Company. Rabbit polyclonal antiserum against LTE4 was obtained from Cascade Biochemicals, UK. Kits from Sigma Chemical Company, St Louis, USA were used for analysis of creatinine. Methacholine chloride (Sigma) was diluted in phosphate buffer saline (pH 7.4) to a concentration of 64 mg/ml and further diluted in isotonic saline.
DATA ANALYSIS
Total airborne dust is expressed as the median and the 25th and 75th percentiles for the three hours of the exposure. Baseline LTE4/9α,11β-PGF2 measurements were made on urine samples collected in the early morning (07.00 hours) prior to entering the piggery on the day of exposure. The peak levels of both urinary mediators following exposure (or at the same time points on the pre- and post-exposure days), as well as the baseline measurements, are expressed as mean (SE). Mediator levels in nasal lavage fluid are presented as median values with 25th to 75th percentiles. The provocative dose (PD20) of methacholine causing a 20% fall in FEV1 was derived by linear interpolation from the log cumulated dose response curves. Calculations of geometric mean (and 95% CI) PD20 values were performed on log transformed raw data.
When data were found to be normally distributed, group mean values were compared using the Student’s t test with p values of <0.05 being considered statistically significant. Differences between baseline concentrations on the three study days were assessed by one way ANOVA analysis. Median, 25th and 75th percentiles for nasal lavage were calculated by the Wilcoxon signed rank test. The statistical calculations were controlled by the use of a validated statistical software package for personal computers (SigmaSuite, Jandel Scientific, USA).
Results
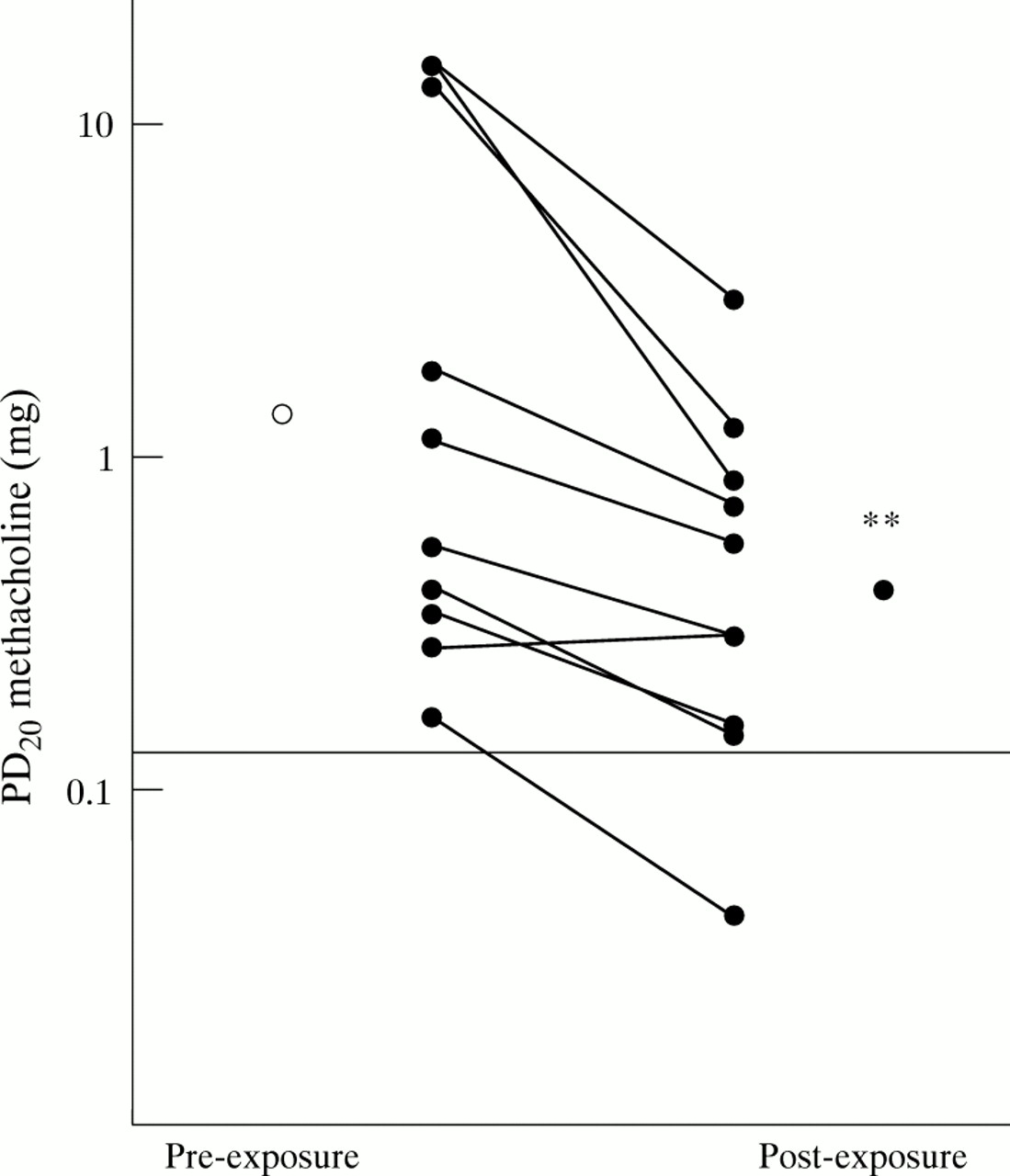
During exposure the median total airborne dust level was 28.5 mg/m3 (26.5–29.5), similar to levels previously documented with this model.18 Bronchial responsiveness to methacholine increased in all but one subject on the day following the exposure (fig 1). PD20FEV1 decreased from 1.32 ( 95% CI 0.22 to 10.25) mg on the pre-exposure day to 0.38 (95% CI 0.11 to 1.304) mg on the post-exposure day (p<0.01; fig 1). The mean (SE) prechallenge baseline FEV1 was the same on both days (3.53 (0.16) l and 3.46 (0.15) l on the pre- and post-exposure days, respectively).

Change in bronchial responsiveness to methacholine after three hours of exposure to swine dust. Data are expressed as the geometric mean. Bronchial responsiveness to methacholine was determined five days before and the day after exposure. Horizontal line indicates the fifth percentile for the reference material (n = 82). **p<0.01.
There were no changes in urinary levels of LTE4 (fig2A) or 9α,11β-PGF2 (fig 2B) during the pre-exposure day. In contrast, urinary excretion of both LTE4 and 9α,11β-PGF2 increased significantly (p<0.01) on the day of exposure (fig 2A and B). For LTE4, the maximum increases were observed 4–6 hours after the subjects had entered the piggery (fig 3). The magnitude of the increase corresponded to a doubling of the mean baseline concentration of urinary LTE4. A similar pattern was seen for urinary excretion of 9α,11β-PGF2 on the exposure day but the relative increase was larger, approximately three times that of the mean baseline value (fig 2B). Urinary levels of 9α,11β-PGF2and LTE4 returned to pre-exposure levels by late evening of the exposure day (fig 3). There was no change in the levels of either LTE4 or 9α,11β-PGF2 during the pre- or post-exposure days (fig 2A and B). Baseline urinary values (mean values in the urine samples collected between 06.00 and 09.00 hours) of the two mediators did not vary between the three study days.

Mean and individual urinary levels of (A) LTE4 and (B) 9α,11β-PGF2 in samples collected from 10 healthy subjects on the pre-exposure day (upper panel), post-exposure day (lower panel), or on the day of exposure to swine house dust (middle panel). Open bars show group mean (SE) morning values in samples collected before methacholine challenge and solid bars show group mean (SE) peak values of samples collected after the respective challenges. Individual values are also shown. A significant increase was seen in levels of LTE4 and 9α,11β-PGF2 following exposure to swine house dust but not following methacholine bronchoprovocation (**p<0.01, paired t test).
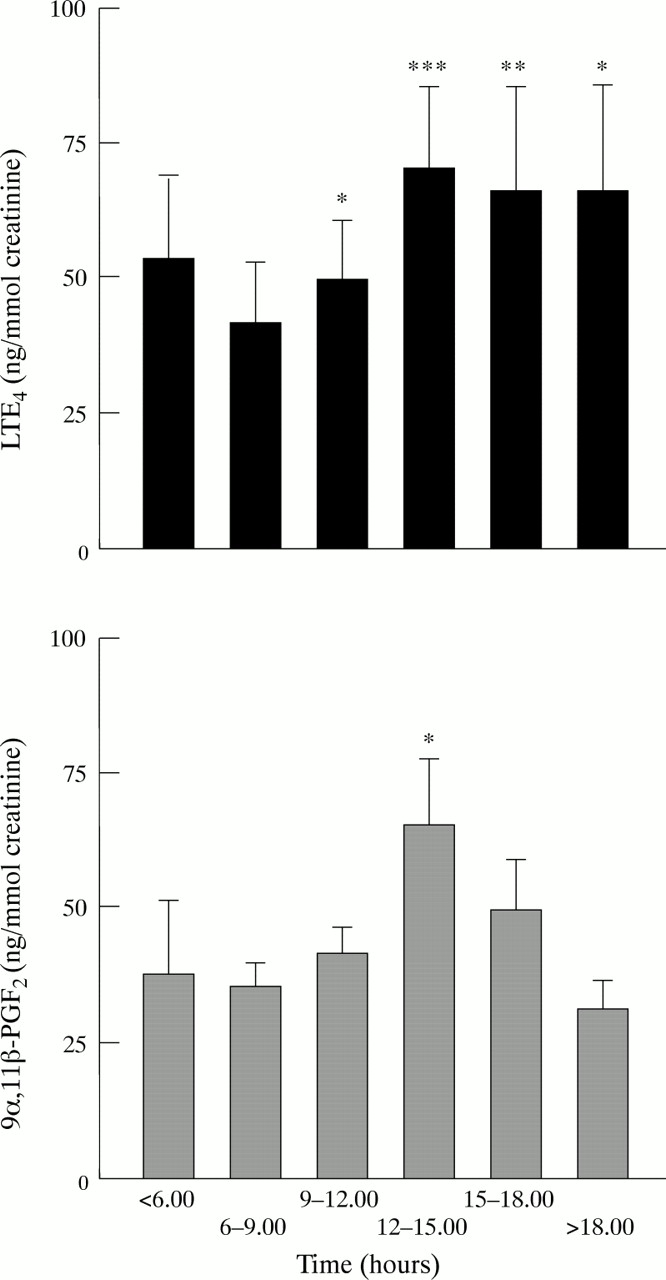
Time course of mean (SE) urinary excretion of LTE4 and 9α,11β-PGF2 during day of exposure to swine house dust. Pre-exposure levels are defined as those in the 06.00–09.00 hours period. Significant increases in both metabolites were observed approximately four hours after the start of exposure to swine dust. ***p<0.001, **p<0.01, *p<0.05.
The median amount of nasal lavage fluid recovered was 7.8 ml (6.8–8.4). The cellular findings in nasal lavage fluid are summarised in table 1. Exposure to swine dust induced a 24-fold increase in the total cell concentration in nasal lavage fluid (p<0.01), mainly due to the increase in neutrophils (p<0.01). The level of IL-8 increased approximately 12-fold following exposure (fig 4). There was more than a doubling of the concentration of LTE4 in nasal lavage fluid collected seven hours after the exposure to swine dust (fig 4). The concentration of LTB4 increased to an even greater extent, and was 5.5 times higher than in the lavage fluid samples taken before exposure to swine house dust (fig 4).
Numbers of cells (× 103ml) in nasal lavage fluid before and seven hours after exposure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Median concentrations (horizontal lines) of (A) IL-8, (B) LTE4 and (C) LTB4 in nasal lavage fluid collected in the morning before entering the piggery and seven hours after exposure to swine house dust. Significant increases were seen in the levels of IL-8 (**p<0.01), LTE4 (**p<0.01), and LTB4 (***p<0.001) following the exposure.
Discussion
The results of this investigation confirm previous indications that a brief three hour exposure of non-atopic healthy volunteers to swine dust is sufficient to elicit a pronounced increase in bronchial responsiveness to methacholine on the following day.6 9The reaction is known to be triggered by airborne dust, and several microbial and non-microbial factors in the air seem to contribute.18 The exposure generates a cascade of inflammatory changes in the airways,4 9 as well as systemic reactions, including increases in serum levels of cytokines such as tumour necrosis factor α and IL-6.5 However, the mechanism behind the increase in bronchial responsiveness is not understood.
The increased formation of cysteinyl leukotrienes observed following the exposure to swine dust therefore raises the possibility of leukotriene involvement in the increased bronchial responsiveness. Cysteinyl leukotrienes certainly possess a profile of biological actions which are consistent with such a role. In addition to direct bronchoconstrictive effects, they induce plasma exudation, mucus secretion, and infiltration of inflammatory cells into the airways, and inhalation of cysteinyl leukotrienes leads to an enhanced airway response to other bronchoconstrictors.19 20
The increase in cysteinyl leukotriene formation was documented both as an increase in the level of urinary LTE4 during the afternoon following the exposure and as enhanced release of LTE4 into nasal lavage fluid. In line with previous observations in healthy subjects17 there was no diurnal variation in urinary excretion of LTE4 during the control day preceding the exposure, nor during the day following the exposure. Since methacholine bronchoprovocation was performed on both these days, the findings confirm earlier observations that directly acting bronchoconstrictors such as histamine8 and methacholine20 do not induce production of leukotrienes or 9α,11β-PGF2.15
The turnover of cysteinyl leukotrienes in the lung is rapid21 and inhaled leukotrienes are recovered in the urine within a few minutes.8 22 Allergen bronchoprovocation of subjects with isolated early reactions is associated with an increase in urinary LTE4 excretion which is maximal within the first two hours after challenge.8The finding of an increase in urinary LTE4 levels in the afternoon following the swine dust exposure therefore indicates that this trigger induced a long lasting activation of cells which produce cysteinyl leukotrienes. The relatively late increase in the appearance of LTE4 in nasal lavage fluid also supports this hypothesis.
The urinary excretion of 9α,11β-PGF2 also increased after the exposure to swine dust. In humans PGD2 appears to be almost exclusively produced in mast cells.15 Increased formation of 9α,11β-PGF2 and other PGD2metabolites has been reported in different circumstances when mast cells are activated or increased in numbers—for example, allergen induced bronchoconstriction,15 systemic mastocytosis, and circulatory shock.23 Our findings suggest that mast cell activation may be one of the factors behind the enhanced generation of cysteinyl leukotrienes. In addition, since 9α,11β-PGF2and PGD2 are both relatively potent bronchoconstrictors,24 their release may have contributed to the increased bronchial responsiveness. Inhalation of PGD2 and LTC4 have additive bronchoconstrictive effects in asthmatics.19 Our findings support previous work suggesting a relationship between increased bronchial responsiveness and mast cell activation. Bronchoalveolar lavage fluid from non-challenged asthmatics has increased concentrations of histamine,13 PGD2, and tryptase, proportional to baseline airway obstruction and increased bronchial responsiveness.12
The increased levels of LTB4 in the nasal lavage fluid may relate to previous findings that neutrophils are a prominent feature of the response to swine dust,5 as is the increase in nasal lavage fluid levels of IL-8,4 which was also confirmed in this study. However, other cells may have contributed to the increased levels of LTB4.25
The increased bronchial responsiveness which occurs after exposure to swine dust differs from that following allergen exposure in that it is not associated with an acute bronchoconstrictor response. However, the exposed individuals did not have asthma and it is not known whether asthmatic subjects will display an exaggerated reaction involving acute airway obstruction if subjected to this particular challenge. Asthmatics are hyperresponsive to cysteinyl leukotrienes26 as well as other bronchoconstrictors. The increase in urinary LTE4 levels after exposure to swine dust was within the range observed after allergen challenges in asthmatics.2 8 This suggests that the leukotriene production we detected would be adequate to cause bronchoconstriction in subjects with a sufficient degree of airway responsiveness. The isolated development of increased bronchial responsiveness may therefore represent a threshold effect of increased production of cysteinyl leukotrienes in the airways of healthy volunteers. The second methacholine challenge was given when the subjects, according to the measured urinary excretion of LTE4, had been exposed to a sustained increase in the leukotriene levels in the airways during the preceding day. It is known that inhalation of cysteinyl leukotrienes leads to an enhancement of the response to histamine or methacholine.19 27 However, since inhalation of cysteinyl leukotrienes may also initiate other cascades, including activation of eosinophils28 and release of proinflammatory cytokines,29 there are several mechanisms which may have contributed to the development of increased bronchial responsiveness.
Obviously, our findings warrant pharmacological intervention to delineate the role of leukotrienes, as well as other mediators, in the increased bronchial responsiveness following exposure to swine dust. Nevertheless, the study raises the possibility that both cysteinyl leukotrienes and LTB4 may be involved in this process, and that mast cell activation is another factor to consider in the reaction. The development of a considerable degree of increased bronchial responsiveness without an obligatory prior bronchoconstriction or changes in baseline pulmonary function would seem to be one particularly useful asset of this human model for the exploration of increased bronchial responsiveness.
Acknowledgments
We thank Lilian Larsson, Siw Siljerud and Britt-Marie Sundblad for excellent technical assistance. The research was supported by Karolinska Institutet and the following Swedish foundations: Work Environment Fund, Farmers’ Foundation for Agricultural Research, Heart Lung Foundation, Swedish Council for Work Life Research 94-1383, Association Against Asthma and Allergy, Medical Research Council (Project 14X-9071), and the Foundation for Health Care Sciences and Allergy Research (Vardal).