Article Text

Abstract
BACKGROUND Mast cells (MCs), which are a major source of cytokines and growth factors, have been implicated in various fibrotic disorders. To clarify the contribution of MCs to fibrogenesis, lung tissue from patients with the acute respiratory distress syndrome (ARDS) was examined during exudative through to fibroproliferative stages.
METHODS Lung tissue was obtained from 17 patients with ARDS who had pathological features of the early exudative stage (n = 6) or the later reparative stages (n = 11), from four patients with idiopathic pulmonary fibrosis, and from three patients with normal lung tissue. Immunohistochemical localisation of tryptase (found in all human MCs), chymase (found in a subset of human MCs), α-smooth muscle actin (identifies myofibroblasts), and procollagen type I was performed.
RESULTS Normal lung tissue exhibited myofibroblast and procollagen type I immunolocalisation scores each of <5 and MC scores of 1. Increased scores were defined as myofibroblast and procollagen type I scores of >10 and MC scores of ⩾2. Eighty percent of lung tissue samples from the early exudative stage of ARDS exhibited increased numbers of myofibroblasts, 50% had increased numbers of procollagen type I producing cells, while only 17% had increased numbers of MCs compared with control samples. All samples from the later reparative stages of ARDS had increased numbers of myofibroblasts and procollagen type I producing cells. Increased numbers of MCs were seen in 55% of samples from the reparative stages. There was no significant shift in MC phenotype in the ARDS samples.
CONCLUSIONS Increased numbers of myofibroblasts and procollagen type I producing cells were frequently found early in the course of ARDS. MC hyperplasia was unusual during this stage, but was often a feature of the later reparative stages. MCs do not appear to initiate fibroproliferation in ARDS.
- acute respiratory distress syndrome
- mast cells
- pulmonary fibrosis
Statistics from Altmetric.com
Although mast cells (MCs) are often located in areas of established fibrosis, it is unclear whether they actively contribute to fibrogenesis or are merely attracted to areas of injury in a non-specific way.1 Experimental evidence suggests a complex relationship between MCs and fibroblasts. For example, fibroblasts elaborate an important chemotaxin and growth factor for MCs, stem cell factor, which may stimulate the recruitment and differentiation of MCs in developing fibrotic tissue.2 In turn, MCs induce cultured fibroblasts to lose contact inhibition, proliferate and synthesise increased amounts of type I collagen.3 We and others have recently reported that human MCs represent a major source of basic fibroblast growth factor, a potent angiogenic and mitogenic growth factor for a variety of cells which may contribute to fibrogenesis.4 ,5 Two main types of human MCs have been described based on the types of neutral serine proteases in their granules: MCs which contain tryptase only (MCT cells) and MCs which contain both tryptase and chymase (MCTC cells). MCT and MCTC subsets express different cytokine profiles.6 A shift in the distribution of MC phenotypes has been described in several chronic fibrotic lung disorders.7 Thus, it may be that different MC subsets are involved at different stages of fibrogenesis.
The range of pathological changes from exudative diffuse alveolar damage (DAD) to fibroproliferation in patients with the acute respiratory distress syndrome (ARDS) offers a unique opportunity to study fibrogenesis over a relatively compressed period of time. The oedema and alveolar haemorrhage seen in the early exudative stage of ARDS become less prominent as repair is initiated by the organisation of exudative material and the proliferation of alveolar type II cells.8 Myofibroblasts appear as part of the repair process, similar to that seen in healing cutaneous wounds.9 Total lung collagen is increased in ARDS patients who survive more than 14 days.8 Previous investigators have shown that type III collagen is often found in the organising phase of lung injury, while type I collagen predominates in the fibroproliferative phase.10 Lung tissue from ARDS patients with features of different pathological stages may yield clues as to whether MCs participate in fibroproliferation in ARDS and, if so, during which stage of the process. MCs are difficult to locate in routinely processed clinical specimens because they often lose their characteristic metachromatic staining properties with formalin fixation. However, they can now be readily identified using immunohistochemical localisation of tryptase, an enzyme present in all human MCs.11
The aim of this study was to determine whether fibroproliferation in ARDS is preceded by an influx of MCs. We hypothesised that, if MCs are important in the initiation of fibroproliferation, they would probably be present in increased numbers before markers of fibroproliferation were identified using immunohistochemical techniques. We further investigated whether there was a change in the types of MCs populating the lung during this process.
Methods
STUDY POPULATION
Paraffin embedded tissue sections were obtained from archived lung samples of patients with ARDS (n = 17) and idiopathic pulmonary fibrosis (IPF; n = 4). ARDS was defined as respiratory distress requiring intubation and mechanical ventilation, diffuse pulmonary infiltrates, hypoxaemia, and no evidence of congestive heart failure. Normal lung tissue samples were obtained from two patients without pulmonary disease who died of accidental trauma and from one patient who had normal lung tissue surgically removed distal to a lung tumour. Tissue samples from patients with ARDS were selected as representing various pathological stages of the disorder from prior pathological reports. These pathological stages have recently been described in detail by Tomashefski8: the exudative phase(includes capillary congestion, interstitial and alveolar oedema, intra-alveolar haemorrhage, and hyaline membrane formation); themixed/organising (proliferative) phase (includes organising granulation tissue, alveolar type II cell proliferation, with initial migration of mesenchymal cells); and thefibroproliferative phase (includes lung remodelling with increased mesenchymal cells and extracellular matrix). Tissue samples with predominantly exudate, mixed/organising, or fibroproliferative features were obtained from two institutions (nine of 17 ARDS samples from University of Texas Health Sciences Center at San Antonio (UTHSCSA) courtesy of Drs Jacqueline J Coalson and Jay I Peters, and all other samples from the Oregon Health Sciences University (OHSU) Pathology Department). The UTHSCSA samples were obtained as part of a prospective study of acute respiratory failure.12 OHSU subjects met similar criteria for ARDS, although the OHSU patients were not included in the UTHSCSA prospective study. Since the tempo of lung injury and repair can vary from patient to patient, histological criteria were used to classify stages of ARDS rather than the number of days of mechanical ventilation required. Necropsy specimens with apparent autolysis were not included in this study. Because of a scarcity of available tissue samples, not all subjects were able to have samples evaluated with all listed primary antibodies.
PRIMARY ANTIBODIES USED IN STUDY
Mouse monoclonal antibody to tryptase (an enzyme found in all human MCs) was purchased from Dako (Carpinteria, California, USA). No cells other than MCs contain tryptase in appreciable quantities.13 Mouse monoclonal antibody to chymase was purchased from Chemicon (Temecula, California, USA). Chymase is only found in a subset of human MCs and in the vascular endothelial cells of human myocardium. Since myocardial tissue was not included in our samples, any chymase identified was confined to the MCTCsubset of MCs. Rat monoclonal antibody specific to human intracellular procollagen type I was also purchased from Chemicon. Mouse monoclonal antibody to α-smooth muscle actin (ASMA, the dominant isoform of actin expressed in myofibroblasts) was purchased from Sigma (St Louis, Missouri, USA). Titres of antibodies used were determined by exposing slides to varying concentrations of antibodies and visualising the optimal balance between a consistent staining pattern with low background staining at the highest possible dilutions.
TISSUE PROCESSING AND IMMUNOHISTOCHEMISTRY
Tissue samples were formalin fixed, embedded in paraffin, and cut into 5 μm sections. Immunolocalisation of tryptase, chymase, procollagen type I, and ASMA was demonstrated using a three step alkaline phosphatase-avidin-biotin complex (ABC-AP) method (Vector Laboratories, Burlingame, California, USA) as previously described.4 Antigen retrieval was performed by pretreating slides either with hyaluronidase (anti-tryptase and anti-chymase) or trypsin (anti-procollagen type I) as required to optimise staining conditions. No antigen retrieval techniques were needed for localisation of ASMA. The antigen-antibody complexes were visualised using either Fast Red (BioGenex Laboratories, San Ramon, California, USA) or Vector Blue (Vector) as substrates, with 2 mmol/l levamisole added to inhibit endogenous alkaline phosphatase activity. Tissue specimens were then counterstained with Gill III haematoxylin and mounted with crystal mount (Biomeda Corporation, Foster City, California, USA). Control slides were treated similarly, but the primary antibody was replaced with non-immune globulin of the same isotype at the same concentration from the same species as the primary antibody. No staining was evident in any of the control slides incubated with the non-immune immunoglobulins.
For double immunolabelling of tryptase and chymase, slides were first labelled for chymase using the ABC-AP method with Vector Blue as a substrate. Potential non-specific binding resulting from the first labelling was blocked by using the biotin-avidin blocking system as directed by the vendor (Vector). The sections were then incubated with the anti-tryptase antibody which was then localised using the avidin-fluorescein isothiocyanate method as previously described.4
ANALYSIS OF STAINED SLIDES
A semiquantitative analysis of stained slides was undertaken. The number of MCs in the lung parenchyma was determined by viewing the tissue at 250× by light microscopy and counting the number of tryptase positive cells in 30 non-contiguous and non-overlapping fields. Areas around large blood vessels and bronchi and near pleural surfaces were avoided. The total cell count was then assigned a score as follows: 0 = 0–200 cells, 1 = 201–400 cells, 2 = 401–600 cells, 3 = 601–800 cells, 4 = 801–1000 cells, 5 = 1001–1200 cells, 6 = 1201–1400 cells, 7 = 1401–1600 cells, 8 = 1601–1800 cells, 9 = 1801–2000 cells, 10 = ⩾2001 cells total in 30 250× microscopic fields. In order to determine the distribution of MC subsets slides were double labelled with both anti-tryptase and anti-chymase antibodies. The percentage of tryptase positive cells which were also immunoreactive to chymase (MCTC subtype) was determined by examining at least 100 tryptase positive cells in a minimum of six fields on each slide.
The number of cells which immunolocalised procollagen type I or ASMA were counted in 30 non-contiguous and non-overlapping fields, avoiding large blood vessels, bronchi and pleural surfaces. However, because of the difficulty in identifying individual stained cells at a lower power, these slides were examined at 400× by light microscopy. Fields were scored according to the number of stained cells identified: 0 = ⩽5 cells, 1 = 6–20 cells, 2 = ⩾21 cells/400× field. The total score was determined by adding together the individual scores for each of the 30 fields. Thus, the highest score achievable was 60 (a score of 2 for each of the 30 fields). Increased scores were defined as values at least twice that seen in normal tissue. Slides were analysed by a single observer who was unaware of the pathological classification of the tissue.
Data are expressed as mean (SE) and were compared using the Mann-Whitney U test, p values of <0.05 being considered significant.
Results
CLINICAL FEATURES OF STUDY POPULATION
The clinical and pathological characteristics of patients included in the study are summarised in table 1. The mean (SE) number of days on which patients received mechanical ventilation before lung tissue was obtained was 4.7 (1.0) for the exudative DAD samples, 10.5 (4.8) for mixed/organising DAD samples, and 19.0 (3.3) for fibroproliferative DAD samples. All of the patients who had exudative DAD findings had received mechanical ventilation for seven days or less. Thirteen of the 17 ARDS patients (76%) had a proven or suspected infectious aetiology for their ARDS.
Characteristics of study subjects
DESCRIPTIVE IMMUNOHISTOCHEMISTRY
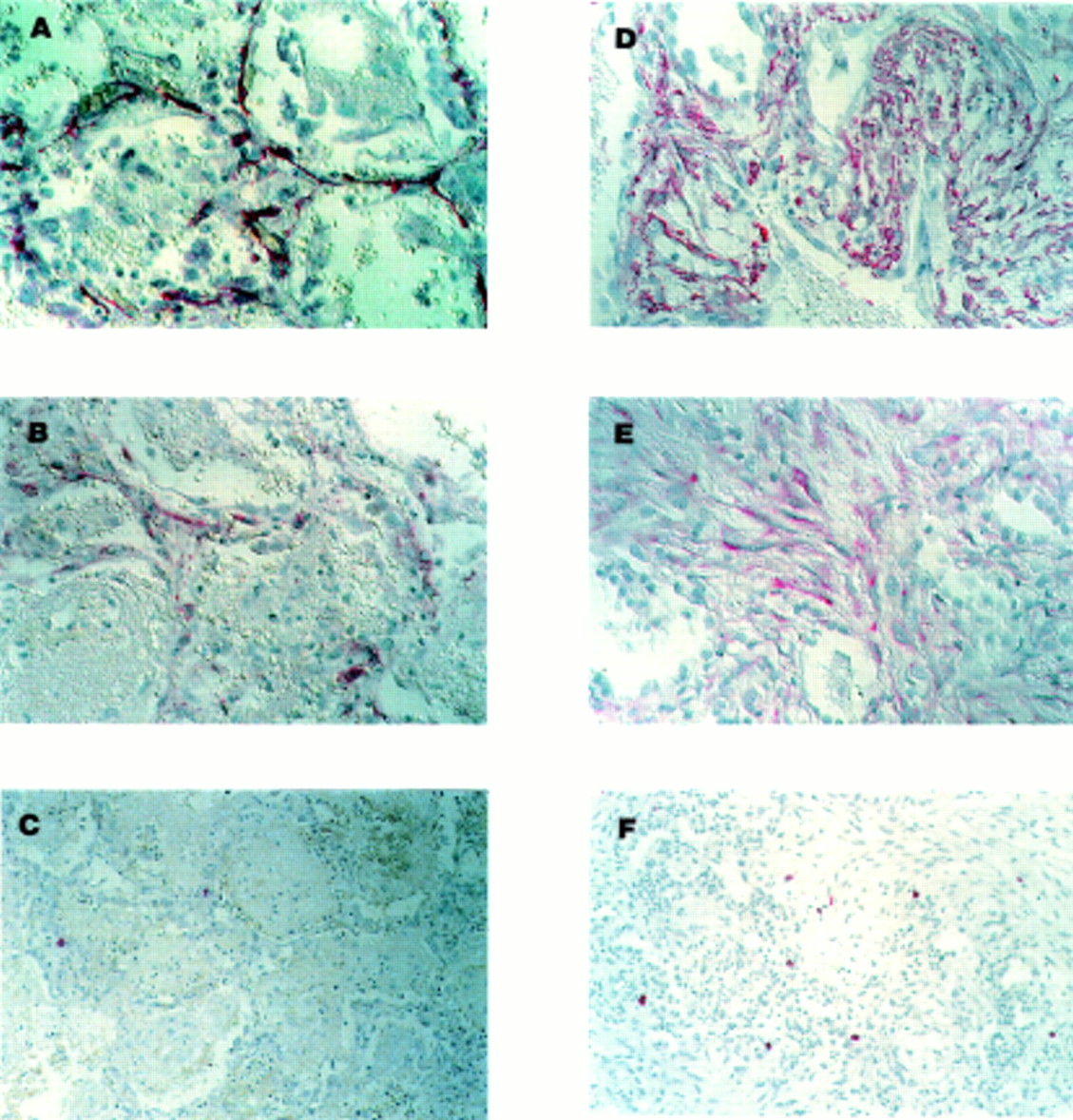
In the early, exudative stage of ARDS the injured tissue exhibited thickening of the alveolar septum, alveolar haemorrhage, and hyaline membrane formation. To identify myofibroblasts and type I collagen producing cells, slides were labelled with antibodies to ASMA and procollagen type I, respectively. Strong cytoplasmic staining for ASMA was found in thin spindle-shaped cells in the alveolar walls (fig 1A). These ASMA positive cells had the morphological appearance of myofibroblasts. No ASMA staining was associated with capillaries, alveolar epithelial cells, or inflammatory cells. Cells that were immunoreactive for procollagen type I in the injured alveolar wall were also seen at this stage (fig 1B). To determine whether there was a simultaneous increase in MC numbers, slides were also labelled with antibody to tryptase which identifies all human MCs. Strong cytoplasmic immunolocalisation of tryptase was seen in a few rounded cells in the alveolar septum (fig 1C). These tryptase positive cells had the morphological appearance of MCs.
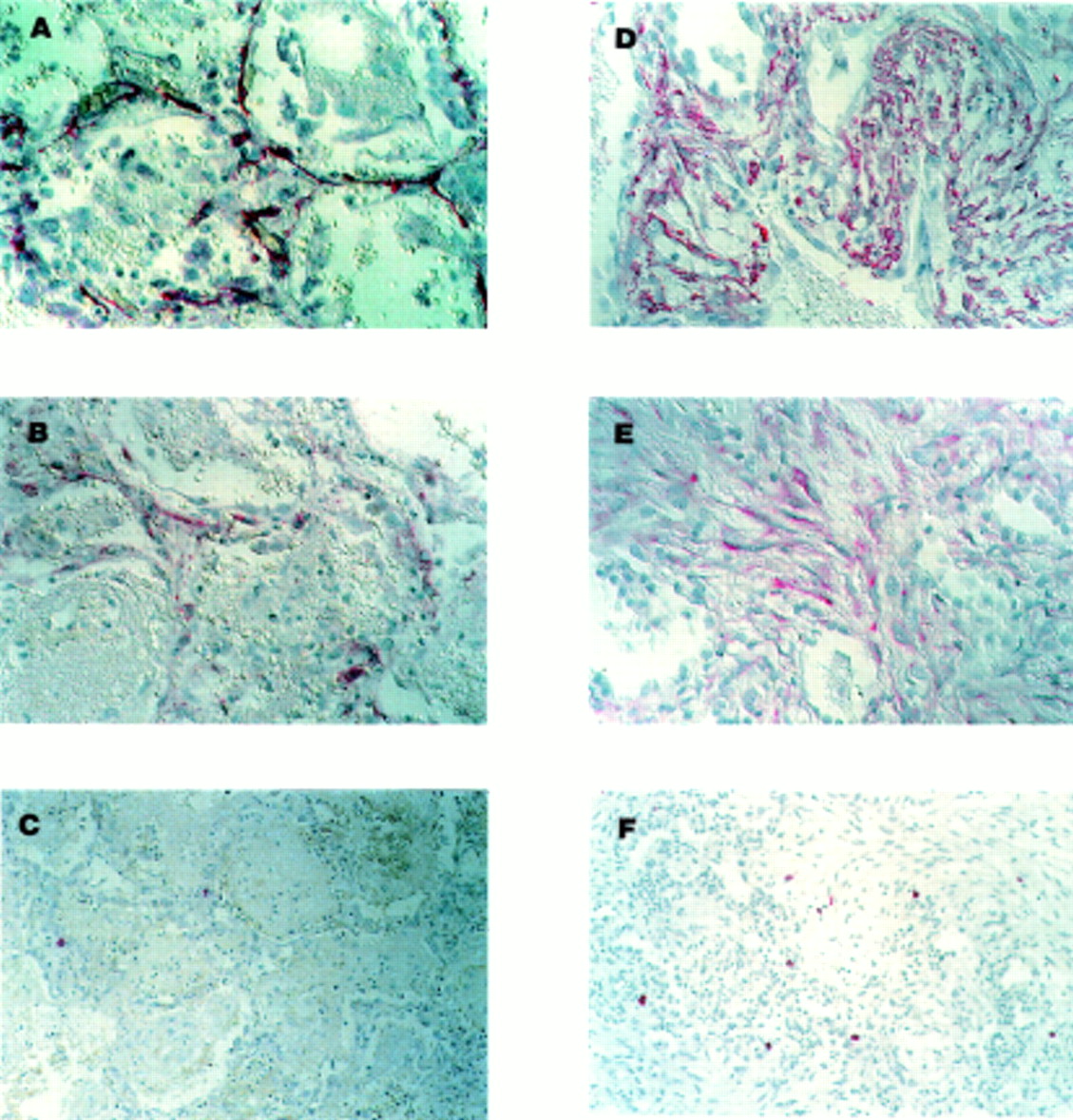
Representative photomicrographs of lung tissue with (A–C) features of the early exudative phase of ARDS (ARDS (E)) and (D–F) features of the reparative stage of ARDS (mixed/organising or fibroproliferative features; ARDS (R)). Immunohistochemical localisation (indicated by red colour) of α-smooth muscle actin (ASMA), procollagen type I, and tryptase is shown as described in the Methods section. ASMA positive myofibroblasts (A) and procollagen type I producing cells (B) are present in the alveolar walls of the injured tissue in the early exudative stage. Few tryptase positive MCs are present at this stage (C). By comparison, nests of ASMA positive myofibroblasts (D) and procollagen type I producing cells (E) are present in samples of the later reparative stage. Increased numbers of MCs are found in areas of fibroproliferation (F). All slides processed with alkaline phosphatase conjugated to avidin-biotin complex with Fast Red as chromogen. Gill’s haematoxylin was used as the counterstain. Magnification 400× for (A), (B), (D), and (E) and 250× for (C) and (F).
In the later reparative stages of lung injury alveolar spaces were frequently replaced with granulation tissue. Nests of loosely organised fibroblasts were often found. Most of the cells in these areas of developing fibrosis were immunoreactive for ASMA (fig 1D). Procollagen type I positive cells were also seen in areas of active fibroproliferation (fig 1E). A modest increase in the number of tryptase positive MCs was identified at this stage (fig 1F). These cells were often found along the edges rather than within the most fibrotic part of the lesions.
SEMIQUANTITATIVE ANALYSIS OF IMMUNOHISTOCHEMISTRY SLIDES
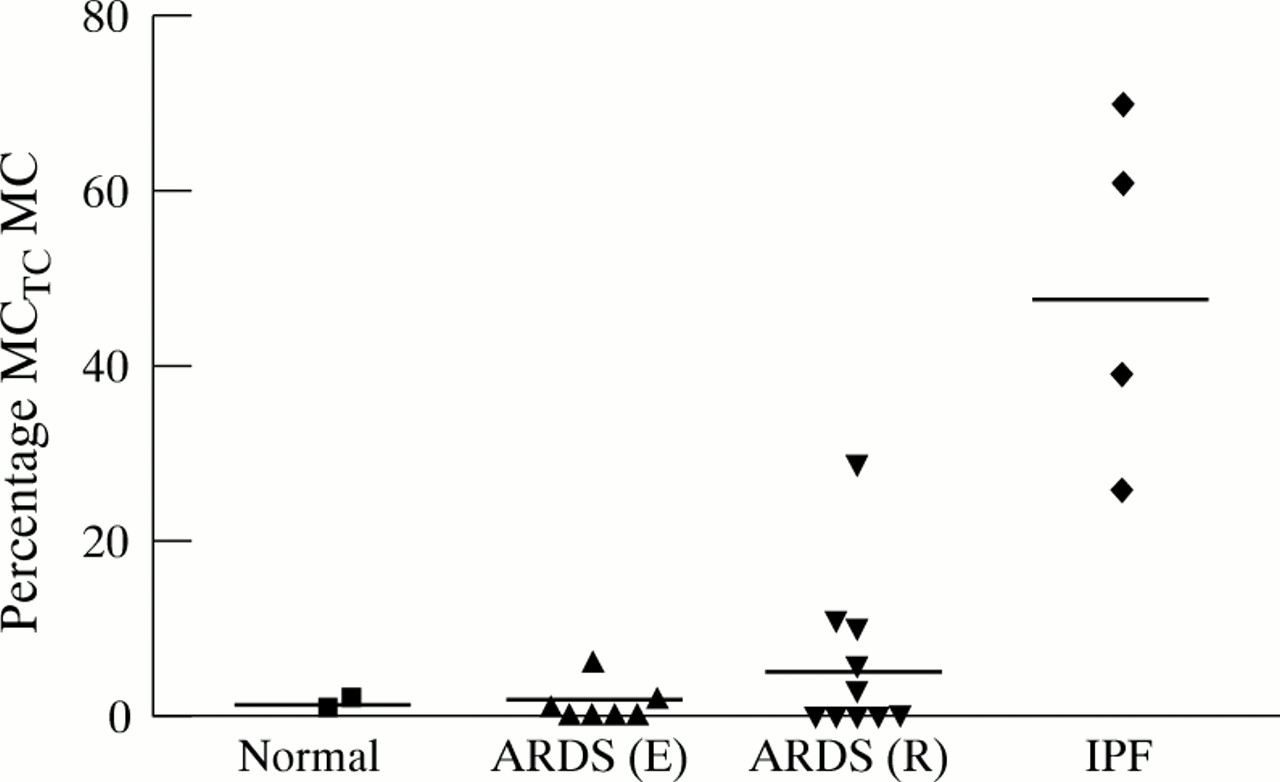
To determine whether there was a relationship between numbers of MCs and the development of fibroproliferation, a semiquantitative analysis was carried out. MC scores from the normal tissues examined were all equal to 1 (fig 2). Although there was a significant increase in the mean MC score in lung tissue from the reparative stages of ARDS (ARDS (R) which includes samples with mixed/organising and fibroproliferative findings) compared with lung tissue from the early stages of ARDS (ARDS (E) which includes samples with exudative findings only), the mean scores from both the ARDS (E) and ARDS (R) samples were not significantly different from normal tissue. Of note, MC scores were raised (⩾5) in all samples from patients with end stage IPF, far exceeding values obtained from any of the ARDS samples.

Relationship between individual mast cell (MC) scores and stage of ARDS using pathological criteria alone. A semiquantitative method was used to determine the MC score in lung parenchyma as described in the Methods section. Horizontal lines represent means of each group. There was no significant difference between MC scores from normal tissue and from samples from the early exudative phase of ARDS (ARDS (E)) or from the later reparative phase (ARDS (R)). There was a significant difference in MC score between normal tissue and idiopathic pulmonary fibrosis (IPF) samples (p<0.05).
In the normal lung at least 90% of MCs exist as MCT subset MCs and the remainder as MCTC subset cells.14To determine whether there was a shift in the distribution of MC subsets with different stages of ARDS, dual labelling of slides with tryptase and chymase antibodies was undertaken. We confirmed the predominance of MCT cells in normal lung tissue (mean (SE) MCTC subset cells 1.5 (0.5)%; fig 3). There was a marked increase in the MCTC subset in the end stage IPF samples (mean (SE) 49 (10)%). Although there was a trend towards an increased percentage of MCTC subset MCs in the ARDS (R) samples (5.9 (2.9)%), the percentage was not significantly different from normal samples.
Percentage of mast cells (MCs) in lung parenchyma expressing the MCTC phenotype in individual samples using pathological criteria alone. Horizontal lines represent means of each group. There was no significant difference in the percentage of MCTC MCs from normal tissue and from samples from the early exudative phase of ARDS (ARDS (E)) or from the later reparative phase (ARDS (R)). There was a significant difference in the percentage of MCTC MCs between normal tissue and idiopathic pulmonary fibrosis (IPF) samples (p<0.05).
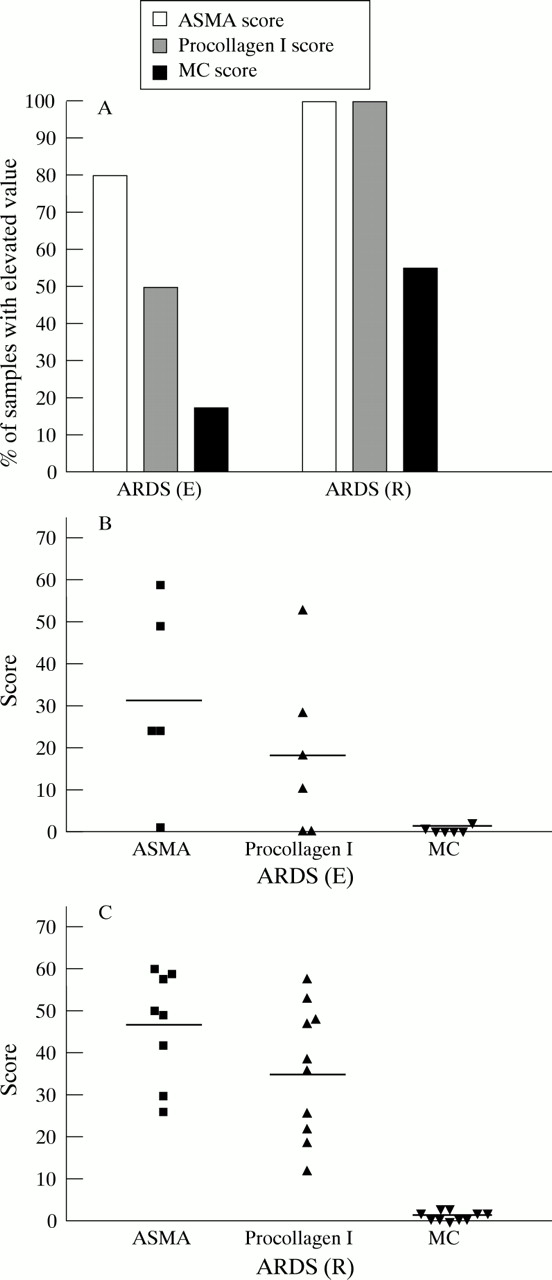
Previous investigators have shown that only airway and vascular smooth muscle cells exhibit immunoreactive ASMA in normal tissue while no immunoreactive ASMA was noted in alveolar walls.15 We also found that ASMA staining in normal lung parenchyma was minimal (ASMA score <5). Increased numbers of ASMA positive cells were found in 80% of the ARDS (E) and 100% of the ARDS (R) samples studied (defined as ASMA score >10), as shown in fig 4A. The mean ASMA score for the ARDS (E) samples was more than three times higher and the ARDS (R) samples more than five times higher than control tissue (figs 4B and4C).

(A) Percentage of samples with increased α-smooth muscle actin (ASMA), procollagen type I and mast cell (MC) scores according to stage of ARDS. Scores for ASMA and procollagen type I stained slides of >10 were defined as increased (normal tissue scores were <5) and MC scores of ⩾2 were defined as increased (normal tissue scores = 1). Individual ASMA, procollagen type I and MC scores for samples with characteristics of (B) the early exudative stage of ARDS (ARDS (E)) and (C) the later reparative stage (ARDS (R)). Horizontal lines represent means of each group.
Mesenchymal cells actively involved in collagen type I synthesis are identified with the anti-procollagen type I antibody. We found minimal immunoreactive procollagen type I localised to alveolar structures in normal tissue (procollagen type I scores <5). Increased numbers of procollagen type I producing cells were found in 50% of the ARDS (E) and 100% of the ARDS (R) samples studied (defined as procollagen type I score >10), as shown in fig 4A. The mean procollagen type I score was less than twice as high in ARDS (E) samples but nearly four times higher in the ARDS (R) samples than in control tissue (figs 4B and 4C). As also shown in fig 4A, there was an increase in MC scores (defined as MC score >2) in only 17% of samples from patients with ARDS (E). The number of patients with increased MC scores rose to 55% in the ARDS (R) samples. However, the change in MC numbers was modest compared with the large increases in ASMA and procollagen type I positive cells (figs4B and 4C).
The relationship between myofibroblast infiltration and MC hyperplasia for all ARDS samples regardless of pathological diagnosis is shown in fig 5A. High ASMA scores were present at all levels of MC infiltration. By comparison, there was a significant increase in the procollagen type I score associated with MC scores of >2 (p <0.05), as shown in fig5B.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Relationship between mean mast cell (MC) scores and mean α-smooth muscle actin (ASMA) scores in ARDS samples regardless of pathological diagnosis. The differences between ASMA scores and the different MC scores were not significant. (B) Relationship between mean MC scores and procollagen type I scores in ARDS samples, regardless of pathological diagnosis. *p<0.05.
Discussion
Although the exudative stage of ARDS is dominated by findings of haemorrhage and oedema, increased numbers of myofibroblasts were present in 80% of samples taken during that early injury phase. Similarly, increased numbers of procollagen type I producing cells were found in half the samples examined. MC hyperplasia was found in only 17% of the exudative stage samples, suggesting that the infiltration of MCs is not a primary event. Thus, the fibroproliferative response appears to be initiated early after the acute injury in many patients with ARDS and occurs without an initial increase in MCs.
All samples taken from the later reparative stages of ARDS showed increased numbers of myofibroblasts and procollagen type I producing cells. Increased numbers of MCs were found in only half of these same samples. There was no significant difference in the percentage of MCs expressing the MCTC phenotype in the ARDS samples compared with normal tissue, although there was a trend towards an increased percentage in the later reparative stage samples. MC hyperplasia was most closely related to the presence of increased numbers of procollagen type I producing cells. Type I collagen, thicker and more resistant to degradation than type III collagen, is generally found in more advanced fibrotic lesions.10 This relationship between MCs and procollagen type I producing cells further suggests that MC infiltration is a later event in fibrogenesis.
We were also interested in learning how the relationship between MCs and fibroproliferation in ARDS compared with that seen in the advanced fibrosis of end stage IPF. In no case did lung tissue from ARDS patients demonstrate the profound MC hyperplasia seen in the IPF samples. In addition, there was a pronounced shift in MC phenotype in the IPF samples, with a eight times greater proportion of MCs of the MCTC subset compared with samples from patients with reparative stage ARDS. Of interest, MCTC subset MCs have been described as “non-immune system-related” in that they are more often associated with angiogenesis and tissue remodelling than with allergic disorders.16 Although the process of fibroproliferation in ARDS and IPF shares some similarities, it is also different in important ways such as initiating factors, time course, and possible contribution of mediators from non-pulmonary organ systems. It is unknown whether the differences in MCs are due to different disease mechanisms or merely reflect a difference in the relative maturity of the fibrotic process.
Previous studies using animal models have supported a role for MCs in the pathogenesis of fibrosis. Using an experimental model of bleomycin induced pulmonary fibrosis in the rat, Goto et al found that MC density was below baseline at day 7, but increased markedly by day 14 when early fibrotic lesions were present. MC density continued to rise for the duration of the study.17 Other investigators have found that MC-deficient animals are less susceptible than controls to lung injury and fibrosis under experimental conditions.18 ,19 A possible explanation for these findings is that MCs are not important in the initiation of fibroproliferation, but are recruited by cells which have already set the stage for mesenchymal cell influx and increased matrix deposition. MCs may then promote the continuation of the fibrotic process.
There are several limitations to studying fibroproliferation and MCs in ARDS patients using archived lung tissue. Individual patients were not studied prospectively and could not be sampled serially over the course of their disease process. Study of the dynamic process of MC degranulation in fixed lung tissue is hampered by the inability to measure MC degranulation products (histamine, proteases, others) in biological fluids. However, several investigators have supported the ability of histological techniques to identify both intact and degranulating MCs.11 ,20 ,21 Since it is difficult to obtain lung samples immediately after the initiation of mechanical ventilation in patients with ARDS, very early time points (within the first few hours) in the course of ARDS have been missed. It would be interesting in future studies to look for evidence of MC degranulation—for example, tryptase or histamine levels—in bronchoalveolar lavage fluid from patients at the early time points. Our study population included predominantly patients who died with ARDS and may not provide information regarding the potential reversibility of this process. Previous investigators have learned that features of fibroproliferation in patients with ARDS are associated with a poor outcome.22 ,23 However, our tissue samples provide some clues as to the possible temporal relationship between MCs and fibroproliferation.
In summary, we found that markers of fibroproliferation were often present early in the course of ARDS. Fibroproliferation was initiated before increased numbers of MCs were evident. We speculate that MCs do not initiate fibrogenesis in ARDS, but may support the continuation of the process by releasing potent mediators into the local environment.
Acknowledgments
Dr Rosenbaum is the recipient of a Senior Scholar Award from Research to Prevent Blindness, New York and Dr Powers is the recipient of a Physician Scientist Award from the National Eye Institute. Lung tissue from the University of Texas Health Science Center at San Antonio was obtained as part of the NIH grants HL-23578 and HL-07221. The authors would like to thank Joshua Eubanks for his technical support during this project.
References
Footnotes
Supported by the American Lung Association of Oregon and the Oregon Health Sciences Foundation Donor Fund.