Article Text

Abstract
Background: The immune response to bacterial antigens on mucosal surfaces may be modified in individuals allergic to aeroallergens due to a maturational or genetic difference or from the interaction between inhaled allergens and bacteria at the mucosa.
Methods: Plasma from children and adults allergic (n = 97) and non-allergic (n = 54) to aeroallergens were initially tested for IgG1 (Th1) and IgG4 (Th2) reactivity to P6, a conserved outer membrane protein of Haemophilus influenzae. IgE binding was measured for some allergic donors. The development of the antibody responses to P6 was subsequently examined in the plasma from 35 children aged 1, 2 and 5 years taken from a prospective birth cohort.
Results: IgG4 antibodies to P6 were more readily detected in allergic subjects than in non-allergic subjects (p<0.001), with a strong bias to the male gender. Some allergic subjects (35%) also had IgE antibody (1–10 ng/ml) that was not associated with IgG4 or gender. In the cohort study of infants, subjects who developed skin prick test positivity to mite allergens by 5 years of age had an 85% reduction in the IgG1 anti-P6 antibody at year 2 (p<0.05) and, unlike skin test negative infants, this group had IgG4 anti-P6 antibodies at 5 years of age.
Conclusions: The antibodies of subjects allergic to a bacterial antigen included IgE and IgG4 (particularly for males) compared with the almost exclusive IgG1 response of non-allergic subjects. The IgG1 responses of 2-year-old children who became skin test positive was markedly reduced and P6-specific IgG4 became detectable at 5 years of age.
Statistics from Altmetric.com
The T cell responses of children who develop allergic sensitisation have been shown to mature slowly. The late maturation has been demonstrated in T cell precursor analyses,1 responses to mitogens2 3 and in the production of Th1 and Th2 cytokines induced by allergens.4 5 Both Th1 and Th2 responses are lower in allergic infants,4 but the Th1 cytokine production has been a centre of focus because interferon-γ (IFNγ) responses are very slow to mature in children,6 leaving the possibility that Th2 imprinting in the neonatal period can persist into infancy. Most of the studies of cytokine regulation of allergic sensitisation have examined the response to allergens. An effect on responses to other antigens could also be expected. This could result from a genetic or environmentally-induced predisposition to producing Th2 responses in general, or be mediated via bystander interactions with the responses to allergens. The latter possibility has been demonstrated by experiments with viral and bacterial infections of sensitised mice.7–9 A greater involvement of the Th2 component of the response of children with aeroallergies to childhood vaccines has been observed.10–12 The responses to mucosal antigens where co-presentation could occur with allergens would therefore be of interest.
To study immune responses to an antigen presented at the respiratory mucosa, the IgE and IgG subclass antibodies induced by a conserved outer membrane protein of Haemophilus influenzae has been examined. These bacteria are highly infectious for children and infect everybody at least once by the age of 2 years.13 The possibility that immune responses to bacteria may be different in people with aeroallergies has been indicated by studies examining IgE antibodies or allergic responses to bacterial extracts.14 The studies here have used assays with defined polypeptide antigens to provide quantitative assays and to differentiate the anti-protein antigen responses from the lipopolysaccharides. Viral infections have been associated with the development of asthma15 16 while, except for Chlamydia and Mycoplasma, bacterial infections have not been considered to be important.16 It is, however, important to distinguish between measuring an association between allergy and infections manifest by disease and examining immune responses to frequent non-pathogenic infections that are essentially part of the normal flora.
METHODS
Study population
The first study compared anti-P6 antibodies of 97 subjects with allergy and 54 without allergy to inhaled allergens. The male to female quotient was 1.3 in the allergic sample and 1.2 in the non-allergic sample. Allergy was defined by symptoms of rhinitis or asthma and by allergen-specific IgE to mite (Dermatophagoides pteronyssinus), cat and/or grass aeroallergens >0.35 kU/l, these being the most relevant allergens in the study area. Non-allergic subjects were those without symptoms of asthma or rhinitis and without serum IgE antibodies to D pteronyssinus, grass pollen or cats. These samples were also divided further with subjects aged >16 years being classified as adults. The adults (58 allergic, 36 non-allergic) were recruited from the Perth general community. Of the 57 children (39 allergic and 18 non-allergic), 20 (13 allergic, 7 non-allergic) were recruited from the general population and 37 (26 allergic and 11 non-allergic) were recruited from an allergy clinic at the Princess Margaret Hospital. The mean ages of the children were 7.1 years (range 1.9–15.2) for allergic subjects and 4.3 years (range 1.3–11.4) for non-allergic subjects, and the mean ages of the adults were 32.7 years (range 16.1–55.2) for allergic subjects and 34.8 years (range 22.3–55.5) for non-allergic subjects. The male:female ratio was 23:16 and 32:26 for the allergic children and adults and 13:5 and 16:20 for the non-allergic children and adults, respectively. The H influenzae type b (Hib) vaccine was introduced to the immunisation schedule in 1991 so most children (49/57) were vaccinated but the adults were not. Data regarding infections and use of antibiotics were not collected for the first study. IgE to D pteronyssinus mite extract, cat and grass extract was measured using the CAPS system (Pharmacia Diagnostics AB, Sweden) performed by the Immunology Department at Princess Margaret Hospital (Subiaco, Western Australia).
The development of the antibody responses was examined in plasma from infants randomly selected from a prospective birth cohort (Childhood Asthma Study) at 1, 2 and 5 years of age. Infants with a positive skin prick test (SPT) were selected solely on the presence of a positive skin test to mite and those with a negative response were selected on the absence of a skin test response to mite, cat and grass at 5 years of age. The infants were recruited prenatally to the cohort based on parental history of atopy as defined by a standard questionnaire4 and a positive doctor’s diagnosis of asthma, hay fever or atopic dermatitis for one or both parents. Preterm babies (<36 weeks), babies with major abnormalities and infants whose family had moved and were unavailable for follow-up were excluded from the cohort. Plasma was collected at time intervals from the age of 12 months. All the infants received the standard West Australian immunisation schedule which included acellular and/or whole cell pertussis (DTPa and/or DTPw), polio and Hib vaccines at ages 2, 4 and 6 months and the measles-mumps-rubella vaccine at 12 months. A further dose of DTPa or DTPw and Hib vaccination was also given at 18 months. Twenty infants (12 boys) were SPT positive to house dust mite extract based on a wheal diameter of ⩾2.5 mm at 5 years of age and 15 infants (8 boys) were SPT negative to mite and grass allergens. Most of the SPT positive infants (15/20) had symptoms of asthma, rhinitis and/or eczema at 5 years of age. There was no difference in the number of upper and lower respiratory tract infections between the SPT negative and positive groups. Two-thirds (66%) of the SPT negative group reported using antibiotics in the first year of life compared with 20% of the SPT positive group (p<0.05), and 75% and 100% respectively of the SPT positive and negative groups had used antibiotics by the second year of life. Antibiotic use at the age of 5 years was 28% and 26% in the SPT positive and negative groups, respectively. The Princess Margaret Hospital human ethics committee gave approval for the procedures in this study.
Antigens
The P6 outer membrane protein of H influenzae was produced as a fusion polypeptide with glutathione-S-transferase (GST) using the pGEX-2T vector (P6-GST). The P6 coding sequence was amplified using DNA from the type b isolate Eagen. A control of the GST was produced directly from pGEX-2T. The proteins were expressed in Escherichia coli TG-1 and isolated from the soluble cytosolic fraction with glutathione coupled sepharose (Amersham Pharmacia Biotech, Uppsala, Sweden). Natural Der p 1 was purified from spent mite medium by antibody affinity chromatography. Natural Der p 3 was purified according to the method of Heymann et al17 and then further substrate affinity purified with benzamidine Sepharose 6B (Amersham Pharmacia Biotech). Natural Der p 4 was similarly purified by substrate affinity with β-cyclodextrin Sepharose 6B (Amersham Pharmacia Biotech). Recombinant Der p 2.0101, Der p 5, Der p 7 and Der p 20 were produced as recombinant polypeptides with a N-terminal hexa-histidine tag and purified by Ni-NTA (Qiagen, Hilden, Germany). P6 was also made with a N-terminal hexa-histidine tag (P6-H6) to confirm binding was due to the P6 component and not to the GST tag. All proteins were further purified by high-resolution size exclusion chromatography and passed over Mustang E filters (Pall Life Sciences, New York, USA) to remove residual endotoxin from the preparations.
Quantification of antibody binding
DELFIA (Wallac, Turku, Finland) assays were used to quantify IgE, IgG1 and IgG4 binding to the P6 antigen and to a panel of mite allergens (Der p 1, 2, 3, 4, 5, 7, 20). The quantity of antibody was interpolated from a standard curve created with human/mouse anti-Der p 2 chimeric antibodies (Indoor Biotechnologies Ltd, Cardiff, UK) by the procedures of Schuurman et al.18 19
The IgE DELFIA assay is described in detail elsewhere.20 21 The IgG DELFIA assays were performed with minor modifications.21 The plates were blocked with 3% human serum albumin (Sigma, USA) in 0.5% bovine serum albumin (CSL, Australia), Tris-HCl, 0.05% Tween 20 buffer and the allergens diluted to 100 ng/ml in DELFIA assay buffer (Wallac). The chimeric standards were either the anti-Der p 2 chimeric IgG1 (diluted 2.5-fold from 0.52 to 5000 ng/ml) or the anti-Der p 2 chimeric IgG4 (diluted from 0.52 to 5000 ng/ml). The biotinylated mouse-antihuman IgG1 or IgG4 monoclonal antibodies (BD Pharmingen, USA) were diluted 1:2000 in assay buffer (Wallac). Antibody binding to P6-GST was confirmed using the P6-H6. The lower limit of detection was approximately 10 ng/ml for IgG1 and 1 ng/ml for IgG4. The sensitivity and specificity of the IgG assays were determined by comparing the titration of the IgG1 and IgG4 chimeric anti-Der p 2 antibodies with titrations of patient sera as described by Shuurman et al19 and by the reactivity to human myeloma IgG1 and IgG4 proteins (Biodesign International, Saco, Maine, USA). The IgG binding could also be inhibited by the specific allergens. The isotype specificity was demonstrated by comparing the IgG1 and IgG4 assays measured with both IgG1 and IgG4 anti-Der p 2 antibodies (only the relevant reactivities were found).
Statistical analysis
As antibody data were highly skewed, they were log-transformed prior to analysis. Differences in the level of antibody binding by selected groups were compared using the non-parametric Mann-Whitney or χ2 tests. All analyses were done using SPSS V.6.1 statistical software (Chicago, Illinois, USA).
RESULTS
The IgG1 and IgG4 titres in the plasma from 97 subjects (39 children, 58 adults) with allergies to common aeroallergens were compared against titres for 54 non-allergic subjects (18 children, 36 adults) to the bacterial P6 outer membrane protein (fig 1A and B). The allergic subjects considered together had significantly more IgG4 in their plasma to the P6 antigen (p<0.001). The adults alone showed a significant difference between allergic and non-allergic subjects (p<0.005), but the difference in the smaller group of children was not significant (p = 0.109). The children and adults had similar IgG1 and IgG4 titres. The subjects were also stratified by gender (fig 2) which showed that the IgG4 response to P6 had a strong bias to allergic males who had more IgG4 than allergic females (p<0.005), although the allergic females also had increased IgG4 (p = 0.07) compared with non-allergic females. The sizes of the IgG4 responses were similar in the allergic male adult and allergic male children groups. IgG1 was readily quantitated in the plasma of both allergic and non-allergic subjects regardless of age and gender, but non-allergic males had lower IgG1 titres than the allergic male (p<0.05) and female groups (p<0.01). The IgG4 titres for the allergic male children and adults correlated strongly with the IgG1 titres (r = 0.45, p<0.001). IgE binding to the P6 antigen was also measured in a selection of plasma from 34 children and 23 adults in the allergic group. As shown in fig 3, the plasma from 35% of allergic donors had IgE reactivity to the bacterial protein ranging from 0.3 to 10 ng/ml. IgE binding was similar between the children and adults (fig 3A) and between males and females (fig 3B).
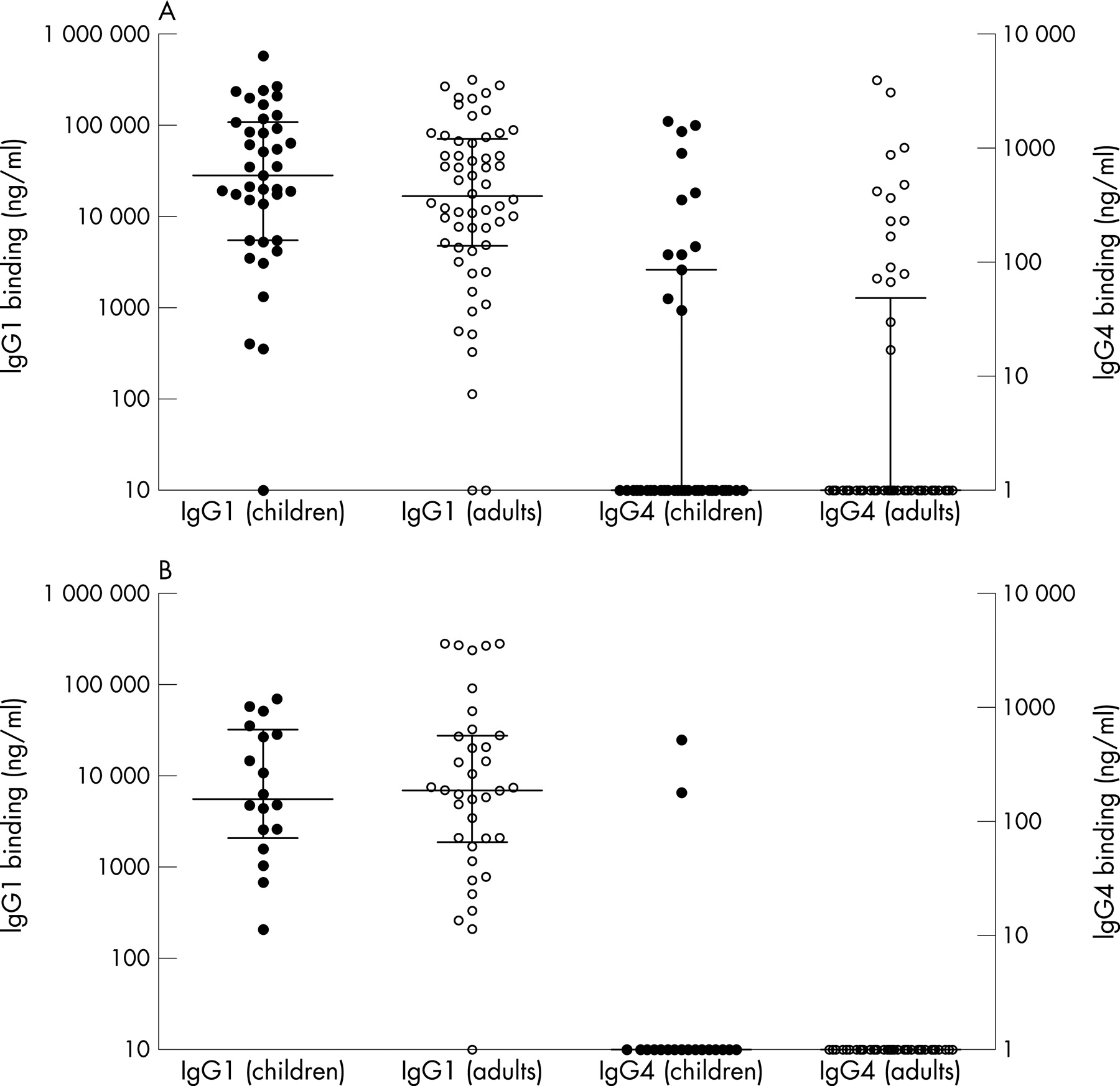

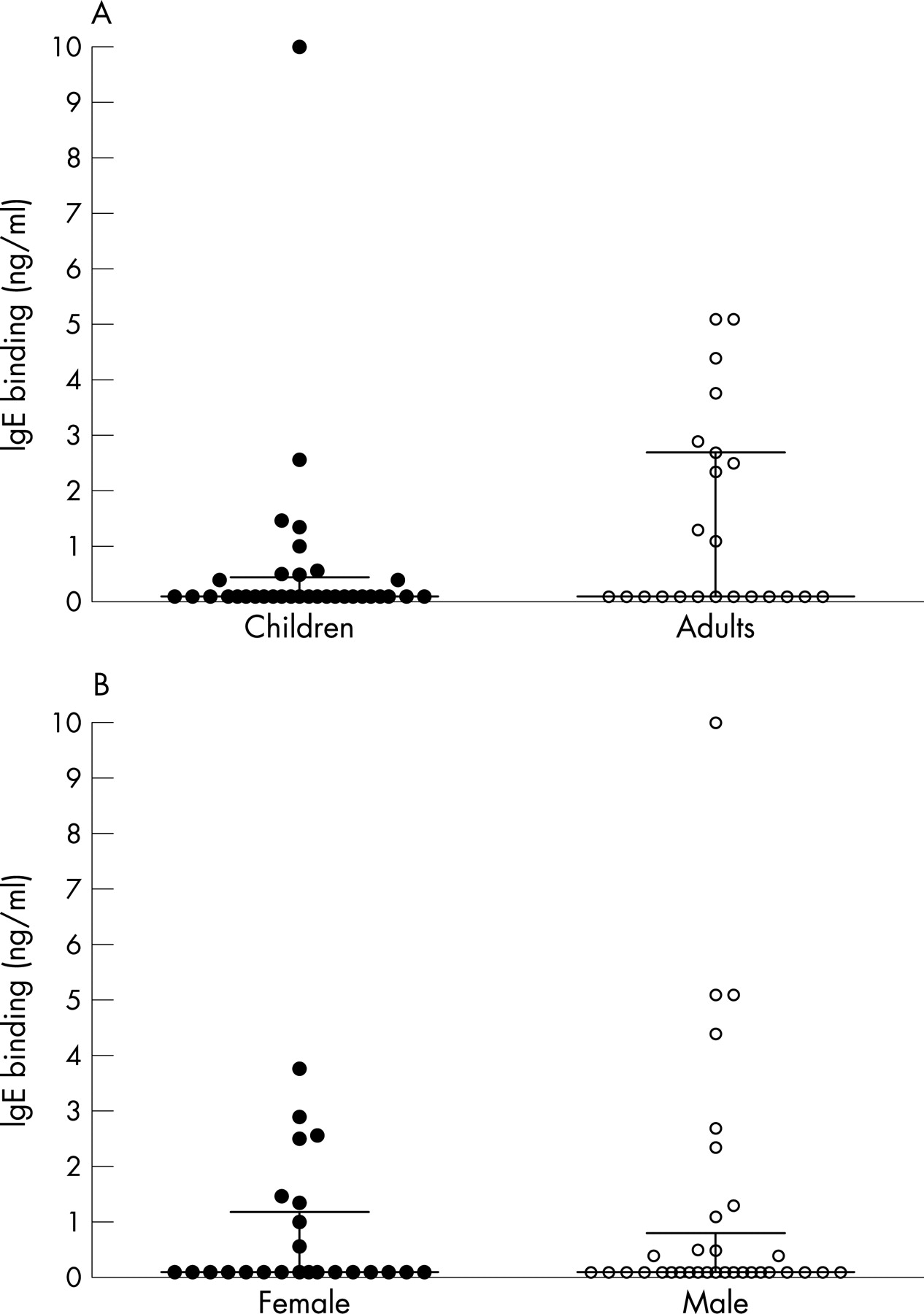
{kind=link}
{kind=link}
{kind=link}
The development of IgG antibodies to the bacterial antigen was examined in the plasma from infants aged 1, 2 and 5 years who were either SPT positive or negative to house dust mite extract at 5 years (table 1). IgG4 against P6 was highest and more prevalent in SPT positive infants at 5 years (p<0.05) and the majority (80%) of the IgG4 responders were male (p = 0.057). IgG1 antibody was readily detected in the first year with a sequential increase in titre at 2 and 5 years. The IgG1 titres were 85% lower at age 2 for the SPT positive group (p<0.05). Low levels of IgE below 3 ng/ml were found against P6, but only at age 5 and in both the SPT positive and negative groups.
Antibody development was also measured to a panel of house dust mite allergens (Der p 1, 2, 4, 5, 7, 20) as described by Hales et al21 at 1, 2 and 5 years. The median titre of IgE or IgG4 binding (ng/ml) based on the summation of binding to each of the six allergens for each SPT positive subject is summarised in table 2. The table is divided into subjects with or without IgG4 to P6. Only SPT positive subjects who had IgG4 responses to one or more individual house dust mite allergens (n = 11) had IgG4 to P6 (n = 5). The IgG4 response to house dust mite was more frequent (p<0.05), and the IgG4 and IgE titres tended to be higher in infants who had IgG4 to P6 (table 2). The IgG4 titres to the mite allergens were typically higher than those found to P6 (up to 3500 ng/ml) at age 5, and IgG4 to mite allergens could be detected earlier at age 2 in subjects with high titres at age 5. As expected, only SPT positive infants had IgE responses to the individual mite allergens of D pteronyssinus which were rarely detected at age 1 and progressively increased in titre by age 2 and 5.
DISCUSSION
The P6 outer membrane protein of H influenzae is highly conserved with respect to sequence and antigenicity.22 It has frequently been used as a marker of H influenzae infection.23 24 Other proteins are also immunogenic, but some such as the major porin show isolate-specific variations.25 As shown here, most of the IgG antibodies found to P6 were of the IgG1 subclass, which is the isotype associated with Th1 immunity. This can be expected of a bacterial antigen. The IgG1 titres are similar to levels reached after immunisation and infection with other bacteria, although it is probable that higher titres result from recent infection.26 27 It was most noticeable that allergic subjects had IgG4 antibodies to P6 whereas they were rare in non-allergic subjects. This was highly significant when comparing the responses of subjects by the simple division of allergic and non-allergic. Further examination found the effect was mostly in males, with 45% showing this Th2-dependent isotype. IgG4 in females was significantly less, but it was also restricted to the allergic group who had a prevalence of 12% with a probability value of 0.07 for the allergic vs the non-allergic groups. Similar trends persisted when the data were divided into those >16 years and those <16 years of age, but the numbers were small and were only significant for the allergic male adults. However, this shows how the altered antibody responses persist with age. Within the allergic groups the size of the IgG4 responses was highly correlated with the size of the IgG1 response, and the IgG4 titres were distributed over a 100-fold range with no association between the size of the response and the presence of IgG4. There was little difference in the antibody response of children and adults both in respect to the magnitude of the titres and the presence of IgG4 antibodies in the allergic population. Most of the children (but not the adults) would have been vaccinated with a Hib conjugate vaccine which indicates that infection with type b H influenzae has had little bearing on the anti-P6 response which is presumably induced by the more ubiquitous untypeable isolates. In addition to the IgG4 antibodies, a number of subjects produced IgE antibodies. The presence of the IgG4 antibodies was confirmed in independent samples of subjects who were SPT positive to house dust mite extracts and examined in infants. The IgG4 antibody was found at 5 years of age but not in samples from those aged 1 and 2 years. The IgG4 titres were only found in children with IgG4 titres to house dust mite allergens and the IgG4 titres to P6 were highest in children with high IgG4 titres to mite. IgG1 to P6 appeared earlier at 1 year of age, with progressively higher titres at 2 and 5 years. The IgG1 titres were lower at 2 years in the plasma from infants who developed a positive SPT, indicating that the development of IgG1 could be slower to mature in this group. Although IgG1 antibodies were readily detectable in all subjects, the allergic group had higher responses of this isotype after infancy and this was also highly significant for males. The antibody titres found in infancy differed because, although the IgG4 was increased in the allergic group, the young children had decreased levels of IgG1.
The increased propensity of infant males to produce IgG4 antibodies and have reduced IgG1 is reminiscent of their increased prevalence of allergic sensitisation in childhood and reports of increased otitis media.28 The development of otitis media and allergy in children is strongly associated with allergic rhinitis.29 This has primarily been attributed to malfunction of the Eustachian tube induced by allergic reactions or depressed mucociliary clearance. The presence of the IgG4 antibodies and reduced IgG1 in early life adds evidence for an immunological mechanism leading to a diminished protective response. Asthma and allergies have been reported as risk factors for carriage of H influenzae30 and Streptococcus pneumonia,31 and there are numerous reports of increased isolation of bacteria from the lungs or nose of subjects with asthma or allergic rhinitis. Asthma and allergies are also risk factors for the development and severity of pneumonia and sinusitis. The diminished IgG1 responses at 2 years of age would be consistent with a decreased capacity to clear infections early in life. The results are of interest with respect to the hygiene hypothesis linking infection with protection from allergic sensitisation. It is possible that the higher IgG1 titres found in infants who did not become allergic indicates that they had more frequent or more severe infections and thus developed a protection from allergic sensitisation. Alternatively, since the immune system has been found to be less mature in children of atopic parents,1 the decreased IgG1 in infancy may be constitutional. The higher rates of infection reported for H influenzae and S pneumonia in allergic individuals30 31 would be consistent with this.
The antibiotic use of the donors in the first study is not known. The fact that the differences were readily detectable in people over 16, who rarely have clinical symptoms from H influenzae, would suggest that the IgG4 antibodies were either induced by subclinical colonisation or were strongly influenced by the increased early exposure found in infancy. Antibiotic use in infants was documented and was found to be greater in the non-allergic subjects in the first year but there was no difference at 2 and 5 years. This does not seem to be associated with H influenzae infection because IgG1 titres were found in all infants at 12 months and the prevalence was the same for the allergic and non-allergic groups. This is consistent with the known high prevalence of subclinical colonisation by H influenzae in young infants.23 Antibiotic use could be expected to reflect a greater susceptibility to infection, but it is not clear whether it would be associated with a consistent pattern of antibody responses and none has been found.32 The treatment of experimental H influenzae infection has been shown to slightly decrease the primary IgG response and slightly increase the response to a second infection.32
It is mechanistically possible for bacterial infections to trigger asthma attacks. H influenzae endotoxin can directly trigger the release of inflammatory mediators from basophils33 and eosinophils,34 and there is evidence from the release of eosinophil cationic protein that they do this in the lungs.35 The IgE shown to be produced to the bacteria could also induce release by an immunological mechanism. Treatment with antibiotics has not, however, given credence for more than an occasional role for asthma triggered by non-atypical bacteria,36 but it has been pointed out that inflammatory effects induced by the bacteria would take time to subside.37 Viral infections are more frequently found to trigger asthma attacks, although this may be easier to identify from the epidemic nature of these diseases and, of course, viruses increase nasopharyngeal bacterial growth. Bacterial infections could have a greater role in the induction of allergic sensitisation rather than exacerbation. H influenzae endotoxin is a ligand for both TLR4 and TLR2.38 Both of these have been implicated in the induction of Th2 responses, especially TLR2.39 It is also of interest that the P6 protein itself is a lipoprotein with a TLR2-binding motif.40
Infants can make IgG4 antibody as shown in food allergy41 42 and responses to vaccines10 and virus infection.43 The IgG4 subclass is, however, slow to mature in ontogeny,44 so it would be expected that the IgG1 subclass would be detected first (as found in this study). Although IgE antibodies to inhalant allergens do not consistently manifest until 5 years of age, the underlying Th2 cell cytokine is detectable early and thus could have bystander influences.5 The early IgG4 anti-P6 responses therefore could just as easily result from bystander influences of cytokine as genetic predisposition, but it clearly shows that the antibacterial responses are present from the earliest time of sensitisation and could have a continued influence. Previous studies on the in vitro cytokine responses of peripheral blood mononuclear cells of adults to P6 only detected IFNγ release and little interleukin (IL)-5 or IL-13 from cells from allergic and non-allergic donors.45 These studies, however, only examined groups of 10 subjects and would not be expected to detect a Th2 component if it was only present in 30% of subjects, as described here. It is also likely that that the high levels of IFNγ found in both allergic and non-allergic subjects would feed back on Th2 cytokine production in the culture system used in that study.
There is a strong association between asthma and IgE antibody titres to the prevailing aeroallergens. While the association is strong on grouped data in children of preschool age with relative risks of developing asthma of 8–12, this is still not predictive in individual children.46 Children with high titres of IgE antibody have only twice the probability of developing asthma as children with 100-fold fewer antibodies. There is thus a strong contribution from other factors. These could be environmental influences or genetic or developmental differences in subjects with asthma in the nature or expression of their inflammatory or tissue response genes. The presence of Th2 responses and a diminished Th1 response to the bacterial antigen could help drive an allergic sensitisation or be a marker of differences in the development of the immune system in allergic subjects. Not all of the sensitised subjects had the IgG4 response, so it could be a marker for a particular allergic phenotype. This could result from a particular genetic predisposition to the induction of Th2 responses or from bystander effects of the responses to the bacteria, or both.
Acknowledgments
The authors thank the study families, study nurses and personnel for recruitment and follow-up of the families; and Lee Hazell for producing the proteins used in the study.
REFERENCES
Footnotes
-
Funding: This work was supported by the National Health and Medical Research Council of Australia.
-
Competing interests: None.