Article Text

Abstract
Background CD200, a cell-surface immunoglobulin-like molecule expressed by immune and stromal cells, dampens the pro-inflammatory activity of tissue-resident innate cells via its receptor, CD200R. This interaction appears critical for peripheral immune tolerance, particularly in the airways where excessive inflammation is undesirable. Vitamin D contributes to pulmonary health and promotes regulatory immune pathways, therefore its influence on CD200 and CD200R was investigated.
Methods CD200 and CD200R expression were assessed by qPCR and immunoreactivity of human lymphoid, myeloid and epithelial cells following 1α,25-dihydroxyvitamin D3 (1α,25VitD3) exposure in vitro and in peripheral T cells following 1α,25VitD3 oral ingestion in vivo. The effect of 1α25VitD3 was also assessed in human airway-resident cells.
Results 1α25VitD3 potently upregulated CD200 on peripheral human CD4+ T cells in vitro, and in vivo there was a trend towards upregulation in healthy, but not asthmatic individuals. CD200R expression was not modulated in any cells studied. CD200 induction was observed to a lesser extent in CD8+ T cells and not in B cells or airway epithelium. T cells isolated from the human airway also responded strongly to 1α25VitD3 to upregulate CD200.
Conclusions The capacity of 1α,25-dihydroxyvitamin D3 to induce CD200 expression by peripheral and respiratory tract T cells identifies an additional pathway via which vitamin D can restrain inflammation in the airways to maintain respiratory health.
- Allergic lung disease
- α1 antitrypsin deficiency
- asthma
- asthma epidemiology
- asthma mechanisms
- innate immunity
- paediatric asthma
- lymphocyte biology
- airway epithelium
- eosinophil biology
- histology/cytology
- lymphocyte biology
- macrophage biology
- neutrophil biology
- asthma guidelines
- cystic fibrosis
- exhaled airway markers
- lung physiology
- paediatric lung disease
Statistics from Altmetric.com
- Allergic lung disease
- α1 antitrypsin deficiency
- asthma
- asthma epidemiology
- asthma mechanisms
- innate immunity
- paediatric asthma
- lymphocyte biology
- airway epithelium
- eosinophil biology
- histology/cytology
- lymphocyte biology
- macrophage biology
- neutrophil biology
- asthma guidelines
- cystic fibrosis
- exhaled airway markers
- lung physiology
- paediatric lung disease
Key messages
What is the key question?
What immunological pathways underlie the role of vitamin D in respiratory health?
What is the bottom line?
The induction of CD200, an immunoregulatory molecule, on human peripheral and airway T cells is a novel mechanism whereby 1α,25-dihydroxyvitamin D3 may act to dampen down inflammation in the airway and maintain respiratory health.
Why read on?
To learn about a novel pathway of regulation in the immune system, and about the mechanisms that contribute to the effects of vitamin D in the airway.
Introduction
CD200 is a member of the immunoglobulin superfamily, expressed by immune cells including thymocytes, T and B lymphocytes and dendritic cells (DCs), as well as neurons, retina, placenta, epithelia and endothelia.1–3 CD200 imparts a unidirectional negative signal via its receptor CD200R2 ,4 to suppress the activation of macrophages,5 mast cell degranulation and cytokine secretion6 and basophil histamine release.7 Animal models demonstrate over-expression of CD200 in vivo or administration of CD200-Fc constructs prolong transplant tolerance and protect against experimentally induced autoimmune disease,2 supporting a central role for this pathway in the regulation of immune responses.
Significant interest exists in the immunomodulatory properties of the steroid hormone vitamin D. The vitamin D receptor is widely expressed throughout the immune system; the metabolic machinery to generate active 1α,25-dihydroxyvitamin D3 (1α25VitD3) exists in the tissue environment and 1α25VitD3 has been demonstrated to inhibit inflammatory T helper 1 (Th1) and Th17-type adaptive immune responses. The effects of 1α25VitD3 on Th2 cell differentiation are less conclusive and are reviewed further in Dimeloe et al.8 1α25VitD3 also promotes regulatory immune pathways via generation of tolerogenic antigen presenting cells (APCs), and regulatory T-cell subsets expressing the transcription factor FoxP3 or the anti-inflammatory cytokine interleukin-10 (IL-10).8 ,9
Pulmonary health requires the effective clearance of pathogens with minimal inflammation, which can damage airway structure and impair gaseous exchange. The capacity of 1α25VitD3 to promote immune tolerance and to enhance antimicrobial mechanisms may underlie epidemiological data associating vitamin D sufficiency with optimal pulmonary function and protection against asthmatic disease.10 ,11 These data led us to investigate additional pathways through which 1α25VitD3 might modulate T-cell function relevant to pulmonary homeostasis.
Recent data suggest that the CD200–CD200R interaction is critical for dampening inflammation in the airway. CD200 is highly expressed on murine airway epithelium12 and CD200R is abundant on alveolar macrophages.13 Mice lacking CD200 demonstrate constitutively activated alveolar macrophages, and exhibit significantly greater pulmonary inflammation and morbidity in response to influenza infection.13 The present study investigated whether vitamin D modulates the CD200–CD200R pathway. It demonstrates that the active form of vitamin D potently upregulates CD200 expression by human peripheral and airway T cells in vitro, and may also do so in peripheral T cells in vivo, following ingestion of active vitamin D by healthy people.
Materials and methods
Patient details
Peripheral blood mononuclear cells (PBMCs) were obtained from healthy donors, tonsillar tissue from children undergoing routine tonsillectomy at St Thomas' Hospital, London and human respiratory tract tissue from adult patients undergoing nasal polypectomy at Guy's Hospital, London. Bronchoscopies in children were performed as previously described.14 All volunteers or parents signed a consent form and all studies were fully approved by the Ethics Committee at Guy's Hospital or The Royal Brompton Hospital respectively. See online methods supplement for details of lavage procedure.
Cell purification and culture
Cell purification was performed as previously described15—further details are given in the online methods supplement. T cells (1×106 cells/ml) were stimulated with 1 μg/ml plate-bound anti-CD3 (OKT-3), 50 U/ml IL-2 (Eurocetus, Harefield, UK) in the presence or absence of 1α25VitD3 (Enzo Life Sciences, Exeter, UK). CD19 B cells were stimulated with anti-CD40 (G28.5; 0.5 μg/ml) and rIL-4 (200 U/ml; R&D Systems, Abingdon, UK) with or without 1α25VitD3. Peripheral CD14 monocytes were matured for 7 days in vitro to a macrophage phenotype using macrophage colony-stimulating factor (MCSF, 100 ng/ml, Sigma-Aldrich, Poole, UK) prior to incubation with or without 1α25VitD3 for 24 h. CD326 nasal epithelial cells (0.5×106 cells/ml) were cultured without or with lipopolysaccaride (500 ng/ml, Sigma-Aldrich, UK) and 1α25VitD3. Total bronchoalveolar lavage (BAL) cells were incubated for 72 h in the presence of 1α25VitD3 when indicated. The effects of 1α25VitD3 on CD200 and CD200R mRNA levels were then assessed by real-time PCR, and cell-surface protein expression by immunoreactivity and flow cytometry analysis. See online methods supplement for details.
1α25VitD3 ingestion by healthy volunteers and patients with asthma
CD3 CD4 T lymphocytes from healthy volunteers (n=4) and patients with asthma (n=3, except at day 7, n=2) were analysed for CD200 and CD200R gene expression directly ex vivo before treatment and at 1, 3 and 7 days after starting a course of oral 1α25VitD3 (0.5 μg/day, Rocaltrol, Roche Products Ltd, Welwyn Garden City, UK). The samples were generated as part of a previously published study,15 approved by the Research Ethics Committee of Guy's Hospital, and with full informed consent from volunteers. See online methods supplement for patient baseline characteristics.
Statistics
The statistical tests used are fully described in the figure legends and further details are available in the online methods supplement.
Results
1α25VitD3 treatment enhances CD200 mRNA and protein expression by human peripheral CD4 T cells and CD8 T cells, but not CD19 B cells
Human T cells express CD200,1 but little is known about the regulation of its expression by these cells. Under culture conditions previously identified to promote a regulatory phenotype in CD4 T cells,15 1α25VitD3 significantly upregulated mRNA transcripts for the CD200 gene in human peripheral CD4 T cells following two 7-day rounds of stimulation with anti-CD3 and IL-2. This occurred in a dose-dependent manner and was maximal at 10−7 M 1α25VitD3 (figure 1A).

1α,25-Dihydroxyvitamin D3 (1α25VitD3) enhances CD200 gene and protein expression by human peripheral CD4 T cells. Human purified peripheral CD4 T cells, stimulated with anti-CD3 (aCD3) and interleukin (IL)-2 for two rounds of 7-day stimulation, in the absence or presence of 1α25VitD3, were assessed for CD200 expression by (A) quantitative PCR (qPCR) and (B,C) flow cytometry. (A) n=6, (B) n=1, representative of four experiments, (C) n=4. (A) CD200 gene expression was assessed by qPCR normalised to 18s expression and is shown as a relative quantity compared with expression under neutral conditions (ie, anti-CD3 plus IL-2) in vitro (median±IQR). p Values derive from Friedman's non-parametric repeated measures test. *p<0.050 as assessed by Dunns paired post-test for multiple comparisons. (B) Grey dots = IgG1 isotype control stain; black dots=CD200 stain. (C) CD200 mean fluorescence intensity (MFI) summarised as mean±SD. p Values derived from a repeated measures analysis of variance test. *p<0.050 as assessed by Bonferroni paired post-test for multiple comparisons.
Flow cytometry analysis demonstrated that 1α25VitD3 significantly enhanced surface expression of CD200 by CD4 T cells at day 14 compared with anti-CD3 and IL-2 stimulation (figure 1B,C). T-cell receptor (TCR) ligation alone increased the frequency of CD200 cells from approximately 20% of cells at day 0 to 88% at day 14. Inclusion of 1α25VitD3 in culture further increased this population to 93% (data not shown) and resulted in significantly greater levels of CD200 expression on a per cell basis (figure 1C).
The kinetics of CD200 surface expression on CD4 T cells in culture was examined daily between days 1 and 6 of culture with IL-2 alone, or with plate-bound anti-CD3, in the absence or presence of 10−7 M 1α25VitD3. Stimulation through the TCR by anti-CD3 induced little surface expression of CD200 at these early time points, which was greatly enhanced with 1α25VitD3. The capacity of 1α25VitD3 to promote CD200 expression required TCR stimulation (figure 2A). Parallel cultures of peripheral blood CD8 T cells demonstrated no increase in cell surface CD200 by TCR activation, with or without the inclusion of 1α25VitD3 in culture at this early time point (figure 2B). However, flow cytometry and quantitative PCR (qPCR) analysis at day 14 of culture indicated a significant increase in CD200 mRNA levels in 1α25VitD3-treated CD8 T cells, although to a lesser degree than observed in CD4 T cells (figure 2C). Human peripheral B cells also express CD200,1 but 1α25VitD3 failed to modulate CD200 gene expression by CD19 B cells at day 14 of culture (figure 2C). Analysis at earlier time points, and of cells cultured with the TLR9 agonist, CpG-ODN, also demonstrated no CD200 modulation by 1α25VitD3, despite a marked increase in expression of hCAP-18, which encodes the antimicrobial peptide cathelicidin, a known target of 1α25VitD3-bound vitamin D receptor8 (data not shown). Thus, the capacity of 1α25VitD3 to increase CD200 gene and protein expression appears to be maximal in CD4 T lymphocytes.
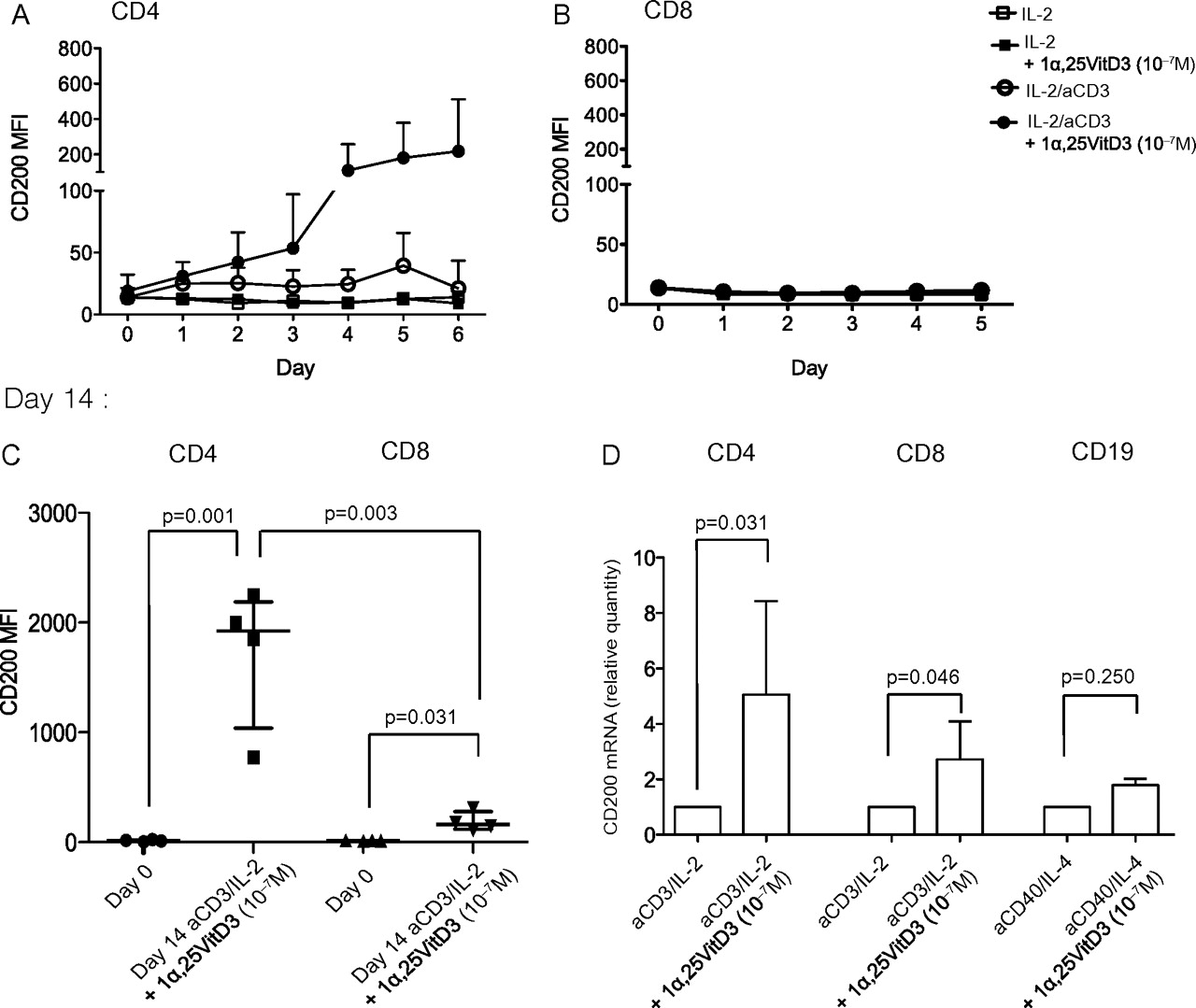
1α,25-Dihydroxyvitamin D3 (1α25VitD3) induces CD200 expression by human peripheral CD4 and CD8 T cells but not CD19 B cells. Human purified peripheral (A,C) CD4 or (B,C) CD8 T cells were cultured with interleukin (IL)-2 alone (50 U/ml) or with additional anti-CD3 (aCD3) stimulation (1 μg/ml, plate bound) in the absence or presence of 10−7 M 1α25VitD3. CD200 expression was assessed by flow cytometry at days 1 to 5 or 6 of culture and at the end of culture at day 14. (A) n=5, (B) n=3, (C) n=4. Data are summarised as mean±SD. p Values derive from paired (within group) or unpaired (between group) t tests. (D) Peripheral CD4 or CD8 T cells cultured as above; or tonsillar CD19 B cells, cultured with anti-CD40 (0.5 μg/ml) and IL-4 (200 IU/ml) in the absence or presence of 10−7 M 1α25VitD3, for two rounds of 7-day stimulation were assessed for CD200 gene expression by quantitative PCR (qPCR). CD200 gene expression was assessed by qPCR normalised to 18s expression as described in the previous figure legend (CD4, n=12; CD8, n=7; CD19, n=3). Data are summarised as median±IQR. *p Values derive from Wilcoxon non-parametric paired tests. MFI, mean fluorescence intensity.
1α25VitD3 does not regulate expression of CD200R by human peripheral T cells, monocyte-derived macrophages or airway-resident cells
The major CD200 receptor, CD200R1 is expressed by T lymphocytes and myeloid cells.16 It is highly expressed in the airway by murine alveolar macrophages,13 in whole human lung tissue17 and human macrophages differentiated with IL-4 and IL-13 to an alternatively activated phenotype, similar to those found in the airway.18 We therefore assessed whether 1α25VitD3 regulated CD200R expression by cells derived from peripheral blood or the human airway. CD4 T lymphocytes were cultured as described above, but qPCR analysis of CD200R mRNA expression failed to demonstrate enhanced CD200R expression by 1α25VitD3 (figure 3A). Flow cytometry analysis at days 0–7 (figure 3B) confirmed this finding and additionally showed a lack of regulation of CD200R expression in response to 1α25VitD3 by CD8 T cells (data not shown). The data do however indicate that CD200R expression is upregulated in both populations following TCR ligation, in agreement with published findings.16

1α,25-Dihydroxyvitamin D3 (1α25VitD3) does not regulate expression of CD200R by human peripheral T cells, monocyte-derived macrophages or airway-resident cells. Human purified peripheral CD4 T cells, stimulated with anti-CD3 (aCD3) and interleukin (IL)-2 for two rounds of 7-day stimulation, in the absence or presence of 1α25VitD3, were assessed for CD200R expression by (A) quantitative PCR (qPCR) at day 14 and (B) flow cytometry at day 7. (A) n=6, (B) n=3 (FACS plots from one representative experiment). (C) n=3. (A) CD200 gene expression was assessed by qPCR normalised to 18s expression. Data are summarised as median±IQR. (B) Grey dots = IgG1 isotype control stain; black dots = CD200R stain. Human purified peripheral CD4 T cells were cultured with IL-2 alone (50 U/ml) or with additional anti-CD3 stimulation (1 μg/ml, plate-bound) in the absence or presence of 10−7 M 1α25VitD3. CD200 expression was assessed by flow cytometry at days 1–6 of culture. Data are summarised as mean±SD. (C) Human purified peripheral CD14 monocytes were cultured with macrophage colony-stimulating factor (100 ng/ml) for 7 days. CD200R and CD206 expression were assessed by flow cytometry at days 1, 3 and 7. Macrophages were then cultured with media alone or 10−7 M 1α25VitD3 for 24 h and CD200R expression was assessed by flow cytometry. Data are summarised as mean±SD. (D) Total bronchoalveolar lavage (BAL) cells were cultured with media alone or 10−7 M 1α25VitD3 for 72 h, CD200R or hCAP-18 gene expression was assessed by qPCR normalised to 18s expression and are shown as a relative quantity compared with expression in media alone. Data are summarised as median±IQR. MFI, mean fluorescence intensity.
CD200R expression was also assessed in monocyte-derived macrophages. Maturation of purified CD14 monocytes with MCSF (100 ng/ml) upregulated surface expression of CD200R in parallel with the prototypical macrophage marker, the mannose receptor (CD206; figure 3C). Subsequent culture of these macrophages in the presence of 10−7 M 1α25VitD3 did not further alter CD200R expression (figure 3C). Finally, CD200R was assessed in human airway-resident cells obtained from BAL fluid, which routinely contain 80–90% macrophages.19 Culture of BAL cells for 72 h demonstrated no significant modulation of CD200R gene expression by 1α25VitD3, although 1α25VitD3 responsiveness was clearly demonstrated by increased hCAP-18 (figure 3D). qPCR analysis was also performed on these cells to assess for regulation of CD200 and the predicted activatory CD200R analogue, CD200R1L. No modulation of CD200 or detectable levels of CD200R1L transcripts were identified in these cells under any culture conditions (data not shown). Together these data suggest that the cell populations studied showed a good response to 1α25VitD3, as assessed by increased hCAP-18, but no modulation of CD200R could be observed.
Effect of ingestion of 1α25VitD3 on CD200 mRNA expression by human peripheral CD4 T cells
To investigate evidence for in vivo modulation of CD200, the effect of ingestion of 1α25VitD3 on CD200 expression by peripheral CD4 T cells was examined in healthy volunteers and patients with asthma in whom a trend towards increased IL-10 and TLR9 gene expression has previously been shown.15 CD200 and CD200R gene expression was assessed in CD3 CD4 T cells purified from freshly derived peripheral blood during the 7-day course of 1α25VitD3 ingestion. There was a clear trend for peripheral CD200 expression to be increased by oral ingestion of 1α25VitD3 in four healthy volunteers studied, to a maximum at day 7. In contrast, CD200 expression was not modulated in three patients with asthma (two at day 7) (figure 4A). CD200R mRNA levels were not altered by oral 1α25VitD3 ingestion in either group, supporting earlier in vitro observations.

Oral 1α,25-dihydroxyvitamin D3 (1α25VitD3) administration in vivo promotes CD200 mRNA expression by human peripheral CD4 T cells. Healthy people (n=4) or people with glucocorticoid-refractory asthma (n=3, except at day 7 when n=2) ingested 0.5 μg 1α25VitD3 daily for 7 days. On days 0, 1, 3 and 7, CD3 CD4 peripheral T cells were isolated and cell pelleted directly ex vivo without any further in vitro manipulation. (A) CD200 and (B) CD200R gene expression were assessed by quantitative PCR normalised to 18s expression and are shown as a relative quantity compared with expression at day 0. Data are summarised as median±IQR. (C, D) Between group differences at day 3 were assessed by Mann–Whitney unpaired non-parametric test.
1α25VitD3 promotes CD200 expression in human respiratory tract CD3 T cells, but not CD326 epithelial cells
Experimental studies have highlighted a protective role of the CD200–CD200R interaction in murine airways. To investigate regulation of this pathway in the human airway, upper respiratory tract tissue was obtained from adults undergoing nasal polypectomy. Nasal polyp tissue on average contains about 40% total leucocytes, of which approximately 25% are CD4 T cells (S Dimeloe, C Hawrylowicz and A Faith, unpublished observations, 2009). However, low cell yields and the requirement to collagenase-dissociate tissue, which cleaves cell surface antigens including CD4, necessitated that total tissue CD3 T cells were isolated (figure 5A). Culture of these purified airway-resident T cells demonstrated that stimulation with anti-CD3 and IL-2 for 7 days modestly increased CD200 gene expression, which was greatly enhanced in the presence of 10−7 M 1α25VitD3 in all four donors tested (figure 5B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1α,25-Dihydroxyvitamin D3 (1α25VitD3) promotes CD200 expression by CD3 T cells, but not CD326 epithelial cells isolated from the human respiratory tract. CD3 T cells were purified from human nasal polyps by cell sorting (typical purity 90%) using a FACSARIA flow cytometer (A). These cells were either unstimulated or stimulated with plate-bound anti-CD3 (OKT-3, 1 μg/ml) and were treated with 1α25VitD3 as indicated. Cell pellets were harvested at day 7. CD200 expression was assessed by quantitative PCR (qPCR) normalised to 18s expression and is shown as a relative quantity compared with expression under neutral conditions (B, n=4). CD326 epithelial cells were purified from human nasal polyps by cell sorting (typical purity 80%) using a FACSARIA flow cytometer. These cells were either unstimulated or stimulated with lipopolysaccharide (LPS, 500 ng/ml) and were treated with 1α25VitD3 as indicated. Cell pellets were harvested at day 7. CD200 and hCAP-18 expression was assessed by qPCR relative to 18s expression and are shown as a relative quantity compared with expression under neutral conditions (C, n=3). Data shown are individual donor responses. IL, interleukin.
Respiratory epithelial cells have been proposed to be a major source of CD200 for interaction with alveolar macrophages.13 Nasal polyp tissue contains approximately 20% CD326 epithelial cells and the capacity of 1α25VitD3 to regulate expression of CD200 in cells enriched for CD326 was therefore assessed (figure 5A). Whilst CD200 mRNA transcripts were detected in sorted CD326 epithelial cells, expression of this gene was not regulated by culture with 1α25VitD3, despite increased transcription of hCAP-18 (figure 5C).
Discussion
Vitamin D sufficiency is associated with reduced incidence and/or better control of respiratory infection, rhinosinusitis, chronic obstructive pulmonary disease and asthma.8 The active form of vitamin D, 1α25VitD3, is produced locally by airway epithelium20 and other airway-resident populations. Local production of 1α25VitD3 is proposed to contribute to airway immunological homeostasis through enhancing innate antimicrobial mechanisms, restraining inflammatory responses and promoting regulatory pathways.8
We now propose a novel mechanism through which vitamin D may maintain immune homeostasis in the airway, by enhancing T-lymphocyte expression of the inhibitory ligand CD200. Interaction of CD200 with CD200R inhibits activation of macrophages, mast cells and basophils.5–7 Functionally, this interaction has been shown to be critical to restrain inflammatory responses in the airways. For example, although influenza infection of mice lacking CD200 results in more rapid clearance of the virus, poorly controlled pulmonary inflammation leads to morbidity, mortality and ‘collateral damage’.13
The capacity of 1α25VitD3 to upregulate CD200 mRNA and protein expression by T cells was dependent upon TCR ligation. Increased CD200 expression following TCR ligation has been proposed to represent a regulatory feedback system to limit effector responses.13 The present study supports this and implies that such a mechanism would be most efficient when vitamin D is replete, in line with epidemiological studies and in vitro data proposing a role for vitamin D in promoting immune tolerance.
Vitamin D circulates in the body almost entirely as the precursor 25(OH)D3, within the range 20–150 nmol/l (2–15×10−9 M). This is converted to 1α25VitD3 in a number of tissues and by stimulated APCs,8 implying that tissue and lymphoid levels may be relatively high, but measurement here is yet to be reported. APC production of 1×10−9 to 6×10−8 M 1α25VitD3 is reported,21 ,22 and CD4 T-cell dose–response studies indicate that 1×10−8 to 1×10−7 M 1α25VitD3 achieves 50–70% interferon γ suppression.23 ,24 Typically doses in the range 1×10−8 to 1×10−6 M are used in vitro8 and our previous studies indicate 1×10−8 to 1×10−7 M 1α25VitD3 optimally promotes IL-10 secretion by human CD4 T cells.15 These observations are echoed here in the case of CD200 expression and both are replicated in vivo using standard prescribed doses of 1α25VitD3.15
Airway-resident CD3 T cells obtained from human upper respiratory tract tissue also responded to 1α25VitD3 for increased CD200 gene expression in culture. Indeed the effect of 1α25VitD3 was more pronounced in airway T cells. These cells may be more responsive to 1α25VitD3—a locally produced mediator during an immune response20—because they comprise a larger proportion of memory T cells, associated with increased vitamin D receptor expression.25 The enhanced inducibility of CD200 expression on airway-resident T cells supports the experimental evidence that this pathway is of particular importance in the tissues.
Enhancement of CD200 by 1α25VitD3 appeared preferential for peripheral and airway-resident CD4 T cells. Airway epithelial CD200 is likely important for basal regulation of alveolar macrophages, which reside in close association with the luminal epithelial surface.13 The current data suggest that in the inflamed airway, local or recruited CD4 T cells, activated through their TCR in the presence of 1α25VitD3, may become an important additional source of CD200. This may facilitate interaction with local DCs, including plasmacytoid DC to reinforce tolerance through the generation of immunosuppressive tryptophan metabolites.26 An important recent paper highlighted a critical role for CD4 and CD8 T cells in the control of innate inflammatory responses in vivo.27 Surface CD200 has been proposed as a mechanism by which lymphocytes can directly interact with innate myeloid cells to regulate their function.28 The relative contribution of epithelial versus lymphocyte CD200 to airway immune homeostasis could be assessed in murine chimaeric studies, however it is interesting that other populations of regulatory CD4 T cells, such as those generated via ligation of the complement receptor CD46, also strongly upregulate CD200.29
In vivo, in healthy human volunteers, a trend towards increased CD200 gene expression in peripheral CD3 CD4 T cells was observed at several time points during a 7-day course of 1α25VitD3 ingestion. This suggests that pharmacological intervention to enhance circulating 1α25VitD3 may promote the CD200–CD200R interaction. However, this may only occur in certain cohorts, as notably increased CD200 was not seen in patients with severe asthma post 1α25VitD3 ingestion, despite increased expression of the IL-10 gene.15 In an independent study profiling gene expression by peripheral lymphocytes during acute asthma exacerbation in children, CD200 was reduced.30 Taken together these data suggest inherent alterations in CD200 expression in asthma, which are not corrected by 1α25VitD3 alone, despite the capacity to promote CD200 expression in healthy people.
In summary, we demonstrate that 1α25VitD3 promotes CD200 expression on human T lymphocytes both in vitro and, in preliminary studies, in vivo. This occurs in peripheral blood T cells, and to a greater extent in T cells isolated from the human airway. We propose this pathway is an additional mechanism by which vitamin D prevents unwanted inflammation in the airways and so maintains pulmonary health. Although the focus of the current study is the respiratory immune system, these findings have broader implications in other physiological systems and disease states. CD200 and CD200R are widely expressed throughout the body and altered expression is associated with a number of other diseases, including multiple sclerosis and Parkinson's disease, for which there are also epidemiological associations with vitamin D.31–34
Acknowledgments
The authors gratefully acknowledge the assistance of the clinical research support staff in the division of Asthma, Allergy and Lung Biology at King's College London and the Department of Pediatric Respiratory Medicine at Royal Brompton Hospital.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Download Supplementary Data (PDF) - Manuscript file of format pdf
Footnotes
Funding SD is the recipient of a Medical Research Council (MRC) funded PhD Studentship through the MRC & Asthma UK Centre for Allergic Mechanisms of Asthma. At the time of this study ZU was funded by an MRC CASE PhD studentship, held in association with Novartis Institute for Biomedical Research. VS is the recipient of an MRC Industrial Collaborative PhD studentship held in association with Novartis, Horsham, UK. AG is the recipient of a British Medical Association James Trust Fellowship. CH acknowledges financial and technical support from the Department of Health via the NIHR comprehensive Biomedical Research Centre award to Guy's & St Thomas' NHS Foundation Trust in partnership with King's College London.
Competing interests None to declare.
Ethics approval Guys Hospital Ethics Committee, Royal Brompton and Harefield NHS Trust Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.