Article Text

Abstract
Whilst patient to patient spread of the respiratory pathogenBurkholderia cepacia is well recognised between patients with cystic fibrosis, prompting a strict segregation policy, cross colonisation between cystic fibrosis patients already infected withB cepacia has not been described and surveys show a very low incidence of patients with more than one strain. Five adult cystic fibrosis patients with B cepacia are presented who became cross colonised with a second B cepacia (UK epidemic) strain, four of whom then died, three from the cepacia syndrome. These cases show that, amongst segregated patients, cross colonisation with different B cepacia strains is possible, and even in these patients the acquisition of the UK epidemic strain may have a fatal outcome. In future it may be necessary to segregate cystic fibrosis patients colonised with the UK epidemic strain from all other patients with cystic fibrosis.
- Burkholderia cepacia
- cystic fibrosis
- cross infection
Statistics from Altmetric.com
Respiratory colonisation of patients with cystic fibrosis with Burkholderia cepacia can cause an accelerated fall in pulmonary function and 20% of cases develop fatal acute fulminant pneumonia (the cepacia syndrome).1 The spread of B cepacia between individuals with cystic fibrosis due to social contact is well recognised2 ,3 and, because of this, a strict segregation policy between B cepacia colonised (BC+) and non-colonised (BC–) patients is advocated in all cystic fibrosis units.4
In the UK a very transmissible strain of B cepacia has been identified and labelled electrophoretic type 12 (ET 12)5 or “UK epidemic” strain, and is present in over 50% of clinics in the UK.6 Furthermore, patients who are already colonised with B cepacia are usually allowed to mix freely with each other, raising the possibility that cross colonisation with this strain may occur. Despite this, there have been no studies to determine whether cross colonisation with B cepacia occurs in this patient group, and surveys in Britain and Ireland have shown a very low incidence of patients colonised with more than one B cepacia strain.6
A number of adult cystic fibrosis patients have been transferred to our clinic from the local paediatric centre who were already colonised with the UK epidemic B cepacia strain, and five other patients colonised with non-epidemic B cepacia have been transferred mainly from peripheral paediatric clinics. In keeping with the national policy, we segregated these BC+ patients from those who were BC–, but all BC+ patients were allowed to mix socially, attend the same outpatient clinics, and were admitted to the same wards for inpatient treatment. All five of the non-epidemic BC+ patients subsequently became cross colonised with the UK epidemic B cepacia strain. Furthermore, four of these rapidly succumbed, three from the “cepacia syndrome”. We present the methods used to type the B cepacia strains in our clinic and also the case histories of these five patients.
B cepacia typing method
The separate B cepacia strains were differentiated by pulsed field gel electrophoresis (PGFE) of macrorestricted chromosomal DNA and polymerase chain reaction (PCR) amplifying the intergeneric region of the 16S and 23S ribosomal RNA genes. PCR amplification of the cable pilus gene was used to identify the strains.
POLYMERASE CHAIN REACTION (PCR) RIBOTYPING
Chromosomal template DNA was extracted by the Chelex-100 resin method.7 Ten colonies of organism were emulsified in 5% (w/v) Chelex solution, boiled for 10 minutes, vortexed for 30 seconds and then centrifuged at 10 000g for one minute. The supernate was pipetted into Eppendorf tubes and refrigerated for use in PCR. Oligonucleotide primers were designed to span conserved regions of the 16S and 23S regions of the rRNA operon.8 The sequences of primers used were 5′-TTGTACACACCGCC CGTCA-3′ for the 16S region and 5′-GGTACCTTAGATGTTTCAGTTC-3′ for the 23S region. Amplification was performed in a mixture of 10mM Tris-HCl (pH 8.8), 2.0 mM magnesium chloride, 0.1% W-1, 200 μM deoxynucleoside triphosphates, and 100 pmol each of primer. Approximately 20 ng of template DNA was used in each amplification. The total volume of the reaction was brought up to 50 μl with distilled H2O and 2.5 U ofTaq DNA polymerase. An initial denaturing step of 95°C for three minutes was followed by 30 cycles of denaturation at 94°C for one minute, annealing at 55°C for one minute, and extension at 72°C for one minute, except for an extension step of four minutes during the last cycle. The products were then separated by electrophoresis for 2.5 hours at 100 V in 1.8% agarose. 100 bp lambda ladders (Bio-Rad) were used as DNA size standards.
PULSED FIELD GEL ELECTROPHORESIS (PFGE)
The bacterial cells were grown overnight in BHI broth, harvested by centrifugation, washed in cell suspension buffer (10 mM Tris, 20 mM NaCl, 50 mM EDTA) and mixed with equal volumes of pulsed field certified agarose (Bio-Rad) containing 4 μl lysozyme. The mixture was dispensed into 100 μl insert molds and allowed to solidify on ice. The plugs were incubated in 40 μl lysozyme buffer (10 mM Tris, 50 mM NaCl, 0.2% sodium deoxycholate, 0.5% sodium lauryl sarcosine) for one hour at 37°C. After washing in wash buffer (20 mM Tris, 50 mM EDTA) the plugs were placed in proteinase K buffer (100 mM EDTA, 0.2% sodium deoxycholate, 1.0% sodium lauryl sarcosine) containing 1 mg of proteinase K and incubated overnight at 50°C. The plugs were then washed in wash buffer with 20 μl of 100 mM PMSF for 0.5 hour. After four further washes of 0.5 hour in wash buffer the plugs were cut with 2 μl Spe 1(Bio-Rad) in 300 μl restriction enzyme buffer (20 mM Tris-HCl, 5 mM MgCl2, 50 mM KCl) and incubated at 37°C for 18 hours. Restriction fragments were separated by PFGE using a CHEF DRII system (Bio-Rad) through 1% (w/v) pulsed field certified agarose in 0.5 × TBE (1 × TBE is 0.9 M Tris-HCl, 0.9 M boric acid, and 1.0 mM EDTA). Electrophoresis was performed for 20 hours in 0.5 × TBE buffer at 14°C with an initial and final pulse time of five and 35 seconds, respectively. Lambda concatamers were used as DNA size standards. The resulting gel pattern was analysed using a Bio-Rad fingerprinting programme to produce a dendrogram of genetic relatedness. The results were corroborated independently by the Epidemiological Typing Laboratory, Central Public Health Laboratory, London.
CABLE PILUS TYPING BY PCR
The method for amplification is identical to that for PCR ribotyping except primers spanning the cable pilus gene9replace the 16S and 23S primers. The sequence of primers used was: sense primer 5′-CCAAAGGACTAACC CA-3′ and antisense primer 5′-ACGCGATG TCCATCACA-3′.It produces an amplicon of 676 bp.
Case histories
CASE 1
An 18 year old man who had cystic fibrosis (DF508/DF508) diagnosed at the age of 20 months had been colonised by Pseudomonas aeruginosa for many years. In 1992 he became colonised with a unique strain of B cepacia. Lung function initially fell and then stabilised and he was requiring four courses of intravenous antibiotics per year. He was admitted in July 1996 with a two week history of increased sputum production and a two day history of increasing breathlessness. He was hypoxaemic (Pao 2 5.6 kPa breathing air), pyrexial (38°C), and had a neutrophil count of 23 000. There were widespread coarse crepitations on auscultation and the chest radiograph showed florid bilateral consolidation. He was commenced on high dose intravenous co-trimoxazole, colomycin, and piperacillin to which previous organisms cultured from sputum were sensitive. Sputum cultured on admission grew a second strain of B cepacia (UK epidemic strain). He deteriorated and died on the ninth day of admission from “cepacia syndrome”.
CASE 2
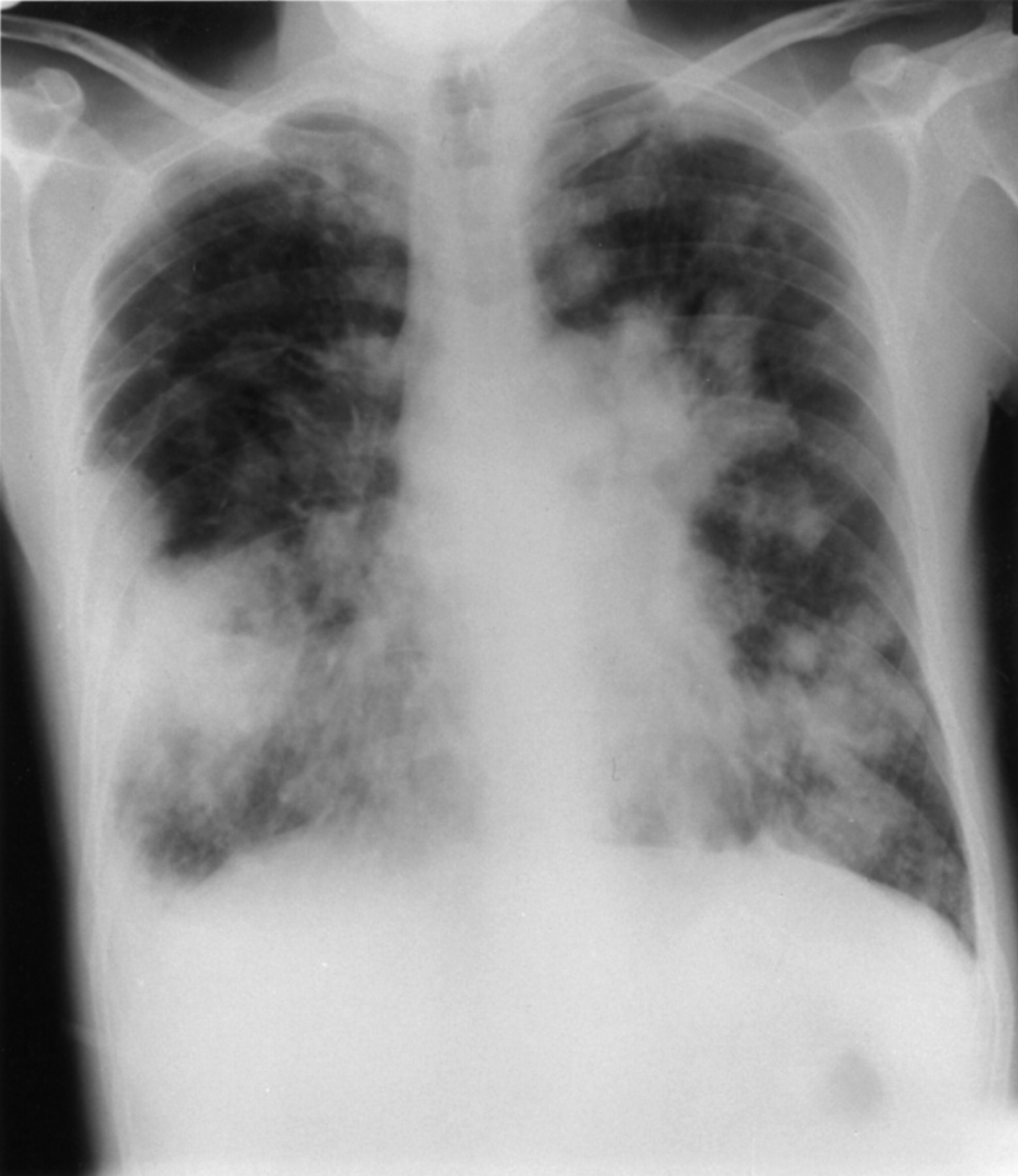
A 34 year old man diagnosed with cystic fibrosis (DF508/DF508) at the age of four months had been colonised with Pseudomonas aeruginosa for many years. A unique strain of B cepacia was intermittently cultured from sputum between November 1992 and November 1993 and continually from then on. Lung function initially fell and then stabilised. By 1996 he was requiring four admissions per year for intravenous antibiotic therapy. He presented in September 1996 with a one week history of increasing sputum production and haemoptysis. On examination he was clubbed, afebrile, and auscultation of the chest revealed coarse crepitations in all zones. He had a mild neutrophilia (8800) and mild hypoxaemia (Pao 2 10.0 kPa on breathing air). His chest radiograph was unchanged from recent previous radiographs. He was started on high dose intravenous ceftazidime and tobramycin to which organisms previously cultured were sensitive. On the tenth day of admission his temperature spiked to 39°C, his sputum production increased, and he became tachypnoeic. His neutrophil count rose to 12 100 and he had worsening hypoxaemia (Pao 27.1 kPa on breathing room air). The chest radiograph showed florid bilateral consolidation (fig 1) and sputum culture revealed a pure growth of B cepacia (UK epidemic strain). P aeruginosa and the original B cepacia strain were now absent. The antibiotics were changed to intravenous colomycin, ceftazidime, co-trimoxazole, and oral chloramphenicol. Despite this, he rapidly succumbed to the “cepacia syndrome”.

Case 2: chest radiograph showing florid bilateral consolidation typical of “cepacia syndrome”.
CASE 3
A 21 year old man was diagnosed with cystic fibrosis (DF508/N1303K) at the age of 18 months. He had been growing P aeruginosa in his sputum for many years. In April 1994 he became colonised by B cepacia (unique strain). Lung function remained stable until July 1994, requiring five courses of intravenous antibiotics per year, when a second strain of B cepaciawas isolated (UK epidemic strain) and the original B cepacia strain could no longer be cultured. Lung function deteriorated and he required continuous intravenous antibiotics until his death from respiratory failure in May 1997.
CASE 4
A 20 year old man was diagnosed with cystic fibrosis (DF508/R553X) at the age of eight months. He had been growing P aeruginosain his sputum for many years. He was colonised by B cepacia (unique strain) before referral to our unit in 1994. In July 1994 he presented with worsening shortness of breath and increasing sputum production. He was pyrexial (39.1°C) with a neutrophilia (37 400) and hypoxaemia (Pao 2 4.8 kPa on breathing room air). There were widespread crepitations and wheezes throughout his chest and the chest radiograph revealed extensive shadowing bilaterally with confluent shadowing at the right base. Sputum culture showed a second strain of B cepacia (UK epidemic strain). Despite treatment with intravenous co-trimoxazole, colomycin and ceftazidime he rapidly succumbed to the “cepacia syndrome”.
CASE 5
A 25 year old man was diagnosed with cystic fibrosis at the age of 10 months. He had been growing P aeruginosa in his sputum for many years. He was colonised by B cepacia (unique strain) in March 1994. Lung function initially fell and then stabilised at a low level (FEV1 30% predicted). In April 1997 a second strain of B cepacia was isolated (UK epidemic strain) and the original B cepacia strain could no longer be cultured. To date there has been no change in his clinical condition.
Both PFGE and PCR ribotyping distinguished each original isolate to be of a unique genotype (figs 2 and 3), whilst the subsequent dendrogram proved the secondary isolates to be indistinguishable from each other and to the prevalent strain in the clinic. Only these secondary isolates possessed the cable pilus gene (fig 4), as did those from all of the other patients in the clinic colonised with B cepacia, therefore confirming the strain as the transatlantic ET 12 (UK epidemic) clone.

PCR ribotyping products analysed by 1.8% agarose gel electrophoresis. Lane 10, 100 bp lambda DNA ladder. Lane 1, original isolate from patient 1; lane 2, secondary isolate from patient 1; lane 3, original isolate from patient 2; lane 4, secondary isolate from patient 2; lane 5, original isolate from patient 3; lane 6, secondary isolate from patient 3; lane 8, original isolate from patient 4; lane 7, secondary isolate from patient 4.

Pulsed field gel electrophoresis in 1% pulse field agarose after cutting with Spe 1. Lane 3, original isolate from patient 1; lane 2, secondary isolate from patient 1; lane 8, original isolate from patient 2; lane 7, secondary isolate from patient 2; lane 9, original isolate from patient 3; lane 1, secondary isolate from patient 3.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cable pili PCR products analysed by agarose gel electrophoresis. Lane 1, 1 kb lambda DNA ladder; lane 2, original isolate from patient 1; lane 3, secondary isolate from patient 1; lane 4, original isolate from patient 2; lane 5, secondary isolate from patient 2; lane 6, original isolate from patient 3; lane 7, secondary isolate from patient 3; lane 12, original isolate from patient 4; lane 11, secondary isolate from patient 4; lane 14, original isolate from patient 5; lane 13, secondary isolate from patient 5.
Discussion
The “epidemic” strain of B cepacia was first isolated in the UK in August 19892 and by 1996 it had been isolated in 50% of UK cystic fibrosis centres and from 38% of allB cepacia samples submitted for genotypic analysis.6 The prevalence of B cepacia in some UK centres has now reached the 40% described in some North American studies.6 In our centre 37 of a total clinic population of 121 adult cystic fibrosis patients have been colonised by B cepacia, and 24 are still alive. Once colonised byB cepacia patients can have varying clinical outcomes. Whilst some remain asymptomatic, others have an accelerated decline in lung function and up to 20% succumb to an overwhelming fatal pneumonia and septicaemia, the “cepacia syndrome”.1 Recent studies from the USA have suggested that the increased risk associated with colonisation with B cepacia in patients with cystic fibrosis is three times that of uncolonised individuals, and that the average life expectancy is almost halved.10
The transmission of B cepacia in patients with cystic fibrosis depends on many factors, and different strains of Bcepacia vary greatly in their rate of transmissibility and transient colonisation can occur. Whilst in some cystic fibrosis patients the source of B cepaciacolonisation is unclear, there is no doubt that patient to patient transmission of the epidemic strain can occur6 ,11-13 and individuals who acquire this strain invariably remain chronically colonised (authors’ unpublished data). This epidemic strain has a high rate of transmission: a patient harbouring two strains of B cepacia transmitted only the epidemic strain to his girlfriend.2
High numbers of B cepacia (>108 cfu/ml) are present in the saliva of colonised patients12 ,13 and indirect spread via contaminated fomites is possible.13 B cepacia has been grown from nebulisers used by colonised individuals14 ,15 and in a study looking at airborne dissemination up to 158 cfu/ml of organisms were recovered from the air of a room occupied by a colonised patient.16 Contamination of the environment by sputum has been shown to persevere for weeks and thus indirect transmission may occur from contaminated surfaces.12 However, the highest risk occurs in the direct exchange of respiratory secretions associated with kissing and the intimate social contact which occurs between family members.2 ,12 Because of these factors, most centres now advocate a strict segregation policy between B cepacia colonised and non-colonised cystic fibrosis patients, in an attempt to limit the spread of this organism throughout the cystic fibrosis population.4 However, there have been no attempts to separate patients colonised with different genotypes ofB cepacia from each other. Indeed, in our clinic cepacia patients were free to socialise with each other whilst inpatients and many of them maintained social contact when in the community.
Colonisation with more than one strain of B cepacia is unusual, having been reported in less than 10% of patients,6 and there have been no reported cases of cross colonisation with different B cepacia strains in individuals who already carry one such genotype. This may be because, although other authors have noted phenotypic variations in B cepacia strains from the same patient, conventional ribotyping techniques are unable to separate them.17 In order to overcome this three molecular techniques were applied in our study to the B cepacia strains to establish their relatedness to each other and to the UK epidemic strain. Firstly, PCR amplification patterns of the intergeneric spacer regions between the 16S rRNA and 23S rRNA was used, a technique shown in 1992 to be capable of separating epidemiologically unrelated isolates of B cepacia 18 and which has been successfully applied to B cepacia strains in other clinics.8 ,19Secondly, PFGE of genomic DNA digested with a rare cutting restriction endonuclease was used. This method has been shown to give different patterns for organisms from different sources,20 is more specific than ribotyping, and has become the gold standard technique in molecular epidemiology.21 Thirdly, specific PCR for the unique pilus form22 of the UK epidemic strain, labelled “cable” pilus due to its length (2 μm) and propensity to intertwine, was used. In all the patients reported here PFGE and ribotyping established the initial B cepacia strains to be of unique genotypes. The strain subsequently acquired in each case was identical to the prevalent strain in the clinic, and PCR for the cable pilus gene established this strain to be of the UK epidemic genotype. Recently, two other epidimicity factors have been described which show a high correlation with the presence of cable pili.23 ,24
This study shows for the first time that, even amongst segregated patients, cross colonisation with epidemic B cepaciastrains can occur. Indeed, all our B cepaciapatients are now colonised by the epidemic strain. Furthermore, in four cases this was associated with a fatal outcome, suggesting that the acquisition of two strains of B cepacia and, particularly, the secondary acquisition of the epidemic strain has a worse prognosis than single strain colonisation alone. In future it may be necessary to segregate cystic fibrosis patients colonised with the UK epidemic strain from all other cystic fibrosis patients, including those with other strains of B cepacia. In our clinic we have now adopted a policy of isolating new patients with B cepaciafrom all other patients until the genotype of their strain has been established, and the strain genotypes of existing cepacia patients are regularly reviewed.
Acknowledgments
We thank Fiona Clode and Mary Kaufmann from the Hospital Infection Laboratory, Central Public Health Laboratory, Colindale, London. The work of M J Gallagher was supported by a grant from the Cardiothoracic Centre.