Article Text

Abstract
Background Sarcoidosis is a systemic disease of unknown aetiology characterised by granuloma formation and the presence of interferon γ (IFNγ)-producing T cells that cause inflammation and tissue damage in multiple organs, especially the lung. Exosomes are nano-sized immunomodulatory vesicles of endosomal origin released from a diverse range of cells and are also found in physiological fluids including bronchoalveolar lavage fluid (BALF) from healthy individuals.
Objective To investigate whether exosomes are enriched in the lungs of patients with sarcoidosis compared with healthy individuals and whether they could contribute to pathogenesis.
Design BALF exosomes from patients with sarcoidosis (n=36) and healthy controls (n=14) were compared by electron microscopy, flow cytometry, western blot analysis and mass spectrometry. BALF exosomes were incubated with autologous peripheral blood mononuclear cells (PBMCs) or the human bronchial epithelial cell line 16HBE14o-. Cytokines were measured by ELISPOT and ELISA.
Results BALF from patients with sarcoidosis showed increased levels of exosomes compared with healthy individuals. Exosomes from patients showed significantly higher expression of MHC class I and II, tetraspanins CD9, CD63 and CD81 as well as neuregulin-1, known to be associated with cancer progression. Furthermore, BALF exosomes from patients induced significantly higher IFNγ and interleukin (IL)-13 production in autologous PBMCs compared with healthy individuals and could also stimulate IL-8 production from epithelial cells.
Conclusion The results indicate for the first time a role for exosomes in human lung disease with possible contributions to the initiation and progression of inflammation in sarcoidosis. This suggests that exosomes may be a new potential target for the clinical treatment of lung diseases.
- Exosomes
- sarcoidosis
- bronchoalveolar lavage fluid
- neuregulin
- interleukin-8
- innate immunity
- lymphocyte biology
- systemic disease and lungs
Statistics from Altmetric.com
- Exosomes
- sarcoidosis
- bronchoalveolar lavage fluid
- neuregulin
- interleukin-8
- innate immunity
- lymphocyte biology
- systemic disease and lungs
Introduction
Sarcoidosis is a systemic granulomatous disease with diverse organ system manifestations that predominantly affects the lung. Common symptoms include dyspnoea, cough and fatigue. Spontaneous remission occurs in most patients but some develop chronic disease leading to death in 1–4% of patients. Although the aetiology of sarcoidosis is not known, many observations including the presence of oligoclonal Th1-like CD4+ T cells and macrophages in bronchoalveolar lavage fluid (BALF) and blood, as well as granuloma formation in the lungs, suggest an antigen-driven autoimmune disease.1 2 Autoantigens such as vimentin and ATP-synthase have been identified as targets for expanded T cell clones in the lung.3 Data suggest contributing roles of Mycobacterium tuberculosis and Proprionibacterium spp. in the pathology of sarcoidosis.4
Exosomes are nano-sized vesicles (30–100 nm in diameter) produced by inward budding of multivesicular bodies. Fusion of multivesicular bodies with the plasma membrane leads to the extracellular release of intraluminal vesicles now called exosomes.5 The term exosome was first coined in 1987 by Johnstone who isolated exosome vesicles from culture supernatants6 and were discovered as a waste product in erythrocyte development. However, a wide variety of other cell types have since been shown to release exosomes, including B cells, dendritic cells (DC), macrophages, mast cells, T cells, epithelial cells, platelets and tumour cells.5 7 The in vivo role of exosomes still remains elusive, but their presence in several body fluids such as breast milk8 and plasma9 suggest a role in vivo. The analysis of exosomes is complicated because of their small size and the lack of exosome-specific markers, hence several methods including electron microscopy are needed to identify them. Exosomes have received great attention lately owing to their potential to induce immune responses or tolerance,10 depending on their cellular origin. Exosomes loaded with tumour antigens are considered as promising vaccine candidates in cancer11 12 and infectious diseases such as toxoplasmosis.13 In 2003 our group isolated exosomes from BALF (BALF exosomes)14 which had phenotypic similarities to DC-derived exosomes. Studies by Prado et al showed that BALF exosomes from tolerised mice can be given intranasally to prevent allergic sensitisation.15 Furthermore, it was recently shown that BALF exosomes from Mycobacterium bovis BCG-infected mice have the capacity to stimulate tumour necrosis factor α (TNFα) production in naïve macrophages.16 Even though the role of exosomes in the lungs or other compartments in humans is not known, they could either participate in immune surveillance16 or in pathogenesis—for example, by potentiating inflammation. In this study we found major differences between BALF exosomes from patients with sarcoidosis and healthy individuals. Patients showed increased numbers of exosomes in their lung compared with healthy individuals and had higher levels of MHC class I and II, tetraspanins and heat shock protein 70 (HSP70). The novel finding that BALF exosomes contain neuregulin-1 (NRG1), previously connected to cancer progression,17 suggests a pathological role for exosomes in sarcoidosis. Furthermore, exosomes from patients could induce IFNγ and IL-13 production in autologous peripheral blood mononuclear cells (PBMCs) and IL-8 production in an epithelial cell line. Taken together, these data suggest that exosomes may contribute to the inflammatory state of sarcoidosis.
Methods
Study subjects and collection of BALF
All patients were recruited from Karolinska University Hospital, Sweden and subjected to bronchoscopy with BAL as part of the routine investigation for sarcoidosis. A diagnosis of sarcoidosis was established by several findings including clinical manifestations, radiological findings, BAL cell differential counts with elevated CD4/CD8 ratio and histological findings of granulomas. Disease activity was determined according to established criteria.18 BALF was obtained from 36 patients with sarcoidosis of median age 42 years (range 23–63); 24 were non-smokers (>2 years) and 12 were current smokers. An overview of the clinical characteristics of the participating subjects is given in table 1. BALF was collected from 14 healthy individuals of median age 25 years (range 20–38), all of whom were non-smokers. The healthy subjects had normal chest x-rays and had no signs of respiratory infection for at least 1 month before BAL. All healthy subjects underwent BAL voluntarily and provided informed consent under protocols approved by the local ethics committee.
Clinical characteristics of patients with sarcoidosis: age, gender, smoking habits, mode of onset, chest radiographic disease stage, smoking habits and treatment with inhaled steroids
Bronchoscopy with BAL (five aliquots of 50 ml phosphate buffered saline (PBS)) was performed as described previously.19 The BALF was strained through a double layer of Dacron nets (Millipore, Bedford, MA, USA) and centrifuged at 400g for 10 min at 4°C. Cell viability was determined by Trypan blue exclusion and was always >90%. For differential cell counts, cytospins were prepared at 20g for 3 min and stained in May–Grünwald Giemsa.
Exosome preparation
Exosomes were isolated by differential ultracentrifugations as described previously14 with some modifications. BALF was centrifuged at 300g followed by 3000g and 10 000g and filtered through 0.2 μm filters (Advantec MFS, Dublin, CA, USA) to remove cell debris and large protein aggregates. Exosomes were pelleted at 140 000g for 2 h 10 min at 4°C. The pellet was dissolved in PBS and the exosomal protein content was measured by DC protein assay (Bio-Rad Laboratories, Hercules, CA, USA) according to the manufacturer's instructions.
Sucrose gradient
BALF exosomes were layered on a continuous sucrose gradient (0.25–2 mM sucrose and 20 mM HEPES/NaOH, pH 7.4) (Sigma-Aldrich, St Louis, MO, USA) prepared manually and centrifuged for 20 h at 79 000g at 4°C. Fractions were collected and the density was determined by refraction index measurements. Each fraction was either coated directly on anti-MHC II beads for analysis by flow cytometry or ultracentrifuged at 200 000g for 35 min for western blot analysis.
Flow cytometry
Pelleted exosomes were adsorbed onto 4.5 μm Ø Dynabeads as described previously.14 1 μl beads were coated with an amount of BALF exosomes corresponding to 6 ml of the original volume of BALF. Exosomes were stained with antibodies to HLA-DR-FITC, MHC class I-FITC, CD40-FITC, CD54-PE, CD63-PE, CD80-FITC, CD81-PE, CD86-FITC, CD9-FITC (BD Bioscience, San Diego, CA, USA) and MUC-1-FITC (Abcam Ltd, Cambridge, MA, USA) and compared with isotype-matched controls (Becton Dickinson, Erembodegen, Belgium). Samples were analysed by a FACS Calibur flow cytometer (Becton Dickinson) and data were analysed using CellQuest Pro software (Becton Dickinson).
Transmission electron microscopy
BALF exosomes corresponding to 3 ml of the original volume of BALF was coated to 1 μl anti-MHC class II Dynabeads. The beads were dehydrated in ethanol followed by acetone and embedded in LX-112 (Ladd, Burlington, VT, USA) and sections were contrasted as previously described.8 Digital images were captured by a Mega View III digital camera (Soft Imaging System GmbH, Munster, Germany).
SDS-PAGE, silver staining and western blot analysis
The pellet from 140 000g centrifugations was denatured in Laemmli sample buffer (Bio-Rad Laboratories), separated on 8–16% SDS polyacrylamide gels (Bio-Rad Laboratories, Hercules CA, USA) and each lane was loaded with 10 μg of exosomes. Gels were either stained with silver staining kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's recommendations or used for immunoblotting. After transfer to PVDF membrane (Millipore, Bedford, MA, USA) and blocking overnight with ECL blocking agent (Amersham Biosciences, Pittsburgh, PA, USA), primary antibodies anti-HLA-DR, anti-HSP70 and β-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA) or anti-NRG1 (Abcam and Santa Cruz) were added and developed as described previously.14
Mass spectrometry analysis
Protein bands were excised manually from the silver-stained gels and subsequently processed and digested by trypsin using a robotic protein handling system (MassPREP, Waters, Milford, MA, USA). Gel pieces were washed with ammonium bicarbonate/acetonitrile at 40°C followed by reducing proteins using DTT/ammonium bicarbonate. The gels were shrunk in acetonitrile and the proteins were alkylated in iodoacetamide. Trypsin was added and the peptides were extracted with 5% formic acid/2% acetonitrile followed by 2.5% formic acid/50% acetonitrile. The tryptic peptide extracts from in-gel digestion were mixed at 1:1 with a saturated solution of α-cyano-4-hydroxy cinnamic acid and dried on a standard steel target plate followed by MALDI MS (Voyager DE Pro, Applied Biosystems, Foster City, CA, USA) in positive ion reflectron mode. For database searches and protein identification, the web site of ProteinProspector (http://prospector.ucsf.edu) was used.
IFNγ and IL-13 ELISPOT
PBMCs were isolated from freshly drawn whole blood by density centrifugation on Ficoll Paque (Amersham Pharmacia Biotech AB, Uppsala, Sweden). Pelleted exosomes corresponding to 6 ml of the original volume of BALF/ml culture medium were then added to autologous PBMCs (300 000 cells/well) and incubated for 48 h in the anti-IFNγ or anti-IL-13 precoated ELISPOT plate (Mabtech, Uppsala, Sweden). Experiments were set up in triplicate and developed according to the manufacturers' instructions. Spots were counted by an ELISPOT reader using AID ELISPOT software (AID, Strassberg, Germany).
16HBE14o- cell culture with BALF exosomes
16HBE14o- cells were kindly provided by Dr D Greunert, University of California, San Francisco, USA.20 Cells were cultured in fibronectin/collagen (BD Biosciences, bovine) coated flasks and grown in minimal essential medium supplemented with 10% fetal calf serum (exosome-depleted), glutamine (200 mM) and penicillin (50 mg/ml) and kept at 37°C with 5% CO2. After growth to confluence, cells were trypsinised using Trypsin-EDTA (R&D Systems, Minneapolis, MN, USA). Viability was controlled by trypan blue exclusion.
16HBE14o- cells (50 000 cells/well) were co-cultured in duplicates with BALF exosomes (1 μg/well or exosomes corresponding to 6 ml of the original volume of BALF/ml culture medium as indicated) from patients with sarcoidosis or healthy controls in a 24-well plate for 48 h. Supernatants were then collected and stored at −20°C until used for IL-8 ELISA according to the manufacturer's instructions (Biolegend, San Diego, CA, USA).
Endotoxin test
The endotoxin test was performed on BALF exosomes (9 patients with sarcoidosis, 9 healthy controls) using the Limulus Amebocyte Lysate endotoxin test (Charles River, Wilmington, MA, USA) according to the manufacturer's instructions. No difference in endotoxin levels was detected in exosome preparations between the two groups and no correlation was seen between the lipopolysaccharide content and cytokine production (data not shown). Lipopolysaccharide levels ranged between 8.1 and 223.2 pg/ml (median 53.4).
Statistical analyses
The results were expressed as individual data for each group. Data were tested for normality by the D'Agostino and Pearson omnibus normality test. Mean values are shown for normally distributed data, otherwise median values are shown. Unpaired t tests were performed on normally distributed data to identify significant differences between experimental groups. Where data were not normally distributed, the Mann–Whitney U test was used to compare groups. p Values were considered significant at <0.05. Correlations were assessed using the Pearson correlation coefficient for normally distributed data and the Spearman ranked correlation coefficient for non-normally distributed data, and associated p values were calculated. Statistical analyses were made with software Graph Pad Prism 5.
Results
Characterisation of isolated exosomes
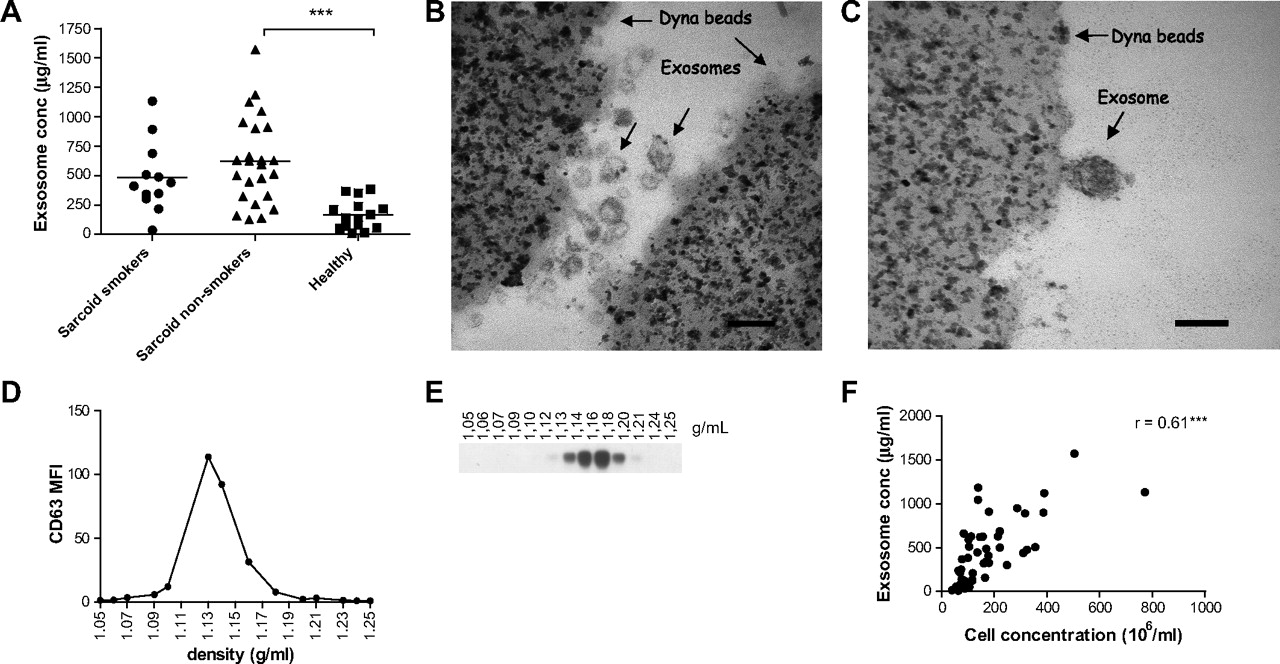
Exosomes were obtained from the BALF of patients with sarcoidosis (n=36) and healthy individuals (n=14) by differential ultracentrifugation. Patients with sarcoidosis were both smokers and non-smokers, but there was no significant difference in their protein content (figure 1A). The level of BALF exosomes of non-smokers with sarcoidosis was, however, significantly higher than that of healthy individuals (p<0.0001). Both electron microscopy (figure 1B,C) and sucrose gradient purification (figure 1D,E) of BAL exosomes revealed a distinct exosome population with typical exosome size (30–100 nm) and density (1.09–1.19 g/ml). They also confirmed that BALF of patients with sarcoidosis contained more exosomes (figure 1B) than BALF from healthy individuals (figure 1C). Immunolabelling showed increased expression of HLA-DR on BALF exosomes from patients with sarcoidosis compared with those from healthy individuals (data not shown). Furthermore, we found a strong correlation between total cell concentration and total exosome protein in BALF (figure 1F).
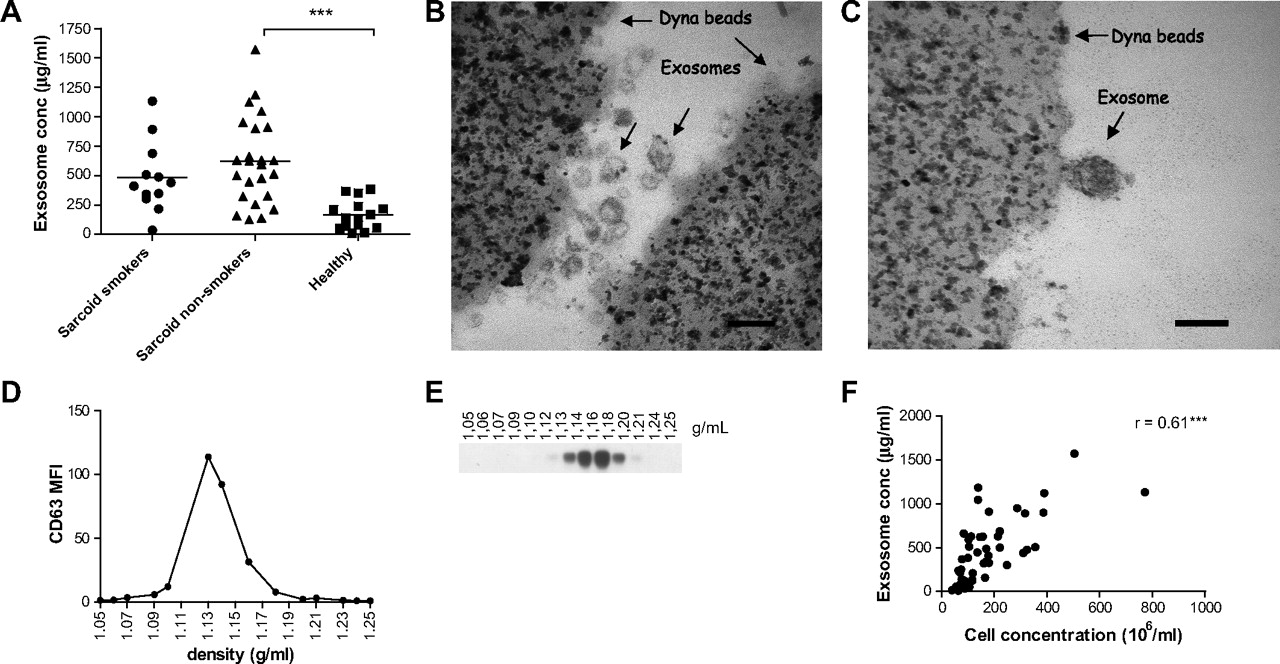
Enhanced levels of total exosomal protein in bronchoalveolar lavage fluid (BALF) from patients with sarcoidosis. (A) Total exosomal protein content in BALF from patients with sarcoidosis (smokers n=12, non-smokers n=24) and healthy individuals (n=14) was determined using the DC protein assay. Results are shown as μg BALF exosomes/ml BALF. Significant differences between groups were assessed using an unpaired t test, ***p<0.001. (B,C) BALF exosomes from one patient with sarcoidosis (B) and one healthy individual (C) were coated to anti-MHC class II beads. Samples were fixed and analysed by electron microscopy. Scale bar 100 nm. Pictures are from one representative individual out of two. (D) FACS analysis of CD63 expression on anti-MHC II bound exosomes from exosome-containing sucrose gradient fractions. (E) HLA-DR expression by western blotting on the different sucrose density fractions. (F) Correlation analysis between cell concentration in BALF and exosomal protein concentration from patients with sarcoidosis and healthy individuals (n=47) assessed using Spearman correlation coefficient (r) and associated p values (***p<0.001).
BALF exosomes were coated to anti-MHC class II beads and analysed by FACS. Exosomes from both smoking and non-smoking patients had significantly upregulated levels of HLA-DR, HLA-ABC, CD9, CD54, CD63 and CD81 compared with healthy individuals (figure 2B–I). There was no difference in CD86 and MUC1 expression (figure 2H,I). CD40 and CD80 were not detected (data not shown). Levels of surface molecules on BALF exosomes did not correlate with stage of disease or disease onset (acute/insidious) (data not shown). A positive correlation was seen between HLA-DR expression and CD9 (r=0.62, p<0.0001), CD63 (r=0.59, p<0.0001) (data not shown) and CD81 (r=0.79, p<0.0001) (figure 3A) on BALF exosomes from both patients and healthy individuals. Interestingly, we found a negative correlation between HLA-DR and MUC1 on BALF exosomes from patients (figure 3B).

Increased levels of certain exosome-associated proteins from bronchoalveolar lavage fluid (BALF) of patients with sarcoidosis compared with healthy controls. (A) Representative FACS graphs on BALF exosomes from one healthy individual. BALF exosomes from patients with sarcoidosis (smokers n=12, non-smokers n=24) and healthy individuals (n=14) coated to anti-MHC class II beads and stained with antibodies against (B) HLA-DR, (C) HLA-ABC, (D) CD9, (E) CD54, (F) CD63 (G) CD81, (H) CD86 (p=0.390) and (I) MUC-1 (p=0.840). Values are presented as Mean fluorescence intensity (MFI) values normalised to isotype controls. Significant differences between groups were assessed using unpaired t test (B, D, H) or Mann–Whitney test (C, E, G, I). *p<0.05, **p<0.01, ***p<0.0001.

Significant correlation between HLA-DR expression and tetraspanins on bronchoalveolar lavage fluid (BALF) exosomes. (A) Correlation analysis of the expression of HLA-DR and CD81 (n=50) on BALF exosomes from healthy individuals and patients with sarcoidosis. (B) HLA-DR and MUC1 (n=35) on BALF exosomes from patients with sarcoidosis. Correlation analysis was assessed by Spearman ranked correlation coefficient (r) and associated p values (**p<0.01, ***p<0.0001). Exosomes were bound to MHC class II beads. Values are presented as MFI values normalised to isotype controls.
Protein composition and identification
SDS-PAGE was performed with exosomes from 14 patients and 7 healthy controls. Silver staining showed several protein bands ranging from 250 kDa to 10 kDa (figure 4A). Three bands around 50, 20 and 10 kDa (figure 4A, marked with arrows) were stronger for patients with sarcoidosis than for healthy controls in most exosome preparations and were therefore excised and analysed by mass spectrometry (identified proteins are shown in table 2). Among the identified proteins we verified the presence of NRG1 on BALF exosomes by sucrose gradient analysis (figure 4B). Western blotting was performed on NRG1 in randomly selected individuals. Three different antibodies directed against full-length NRG1 and different isoforms ((ab2994 (Abcam), SC-348 and SC-80226 (Santa Cruz)) were used. Multiple bands (19–180 kDa) were detected on exosomes when using the SC-348 antibody (9 patients with sarcoidosis, 7 healthy controls, data not shown). However, several high and low molecular weight bands indicative of intracellular cleaved products21 were prominently stronger in exosomes from patients with sarcoidosis but were hardly detectable in those from healthy controls. Based on the molecular weight previously reported for NRG1, we speculate that the 80–140 kDa immunoreactive bands represent full-length NRG121 22 while smaller bands correspond to cleaved NRG1 intracellular fragments or different isoforms.23 The aberrant upregulation of NRG-1 on exosomes from patients was again proved using the SC-80226 antibody, specifically recognising the NRG1 isoform with a molecular weight of 40 kDa (figure 4C). The intensity of the bands when normalised to loading control showed significantly higher levels of the 40 kDa NRG1 in patients with sarcoidosis than in healthy controls (figure 4D). No significant difference, but the same tendency, was seen for the 44 kDa NRG1 β isoform, with a higher level in BALF exosomes from patients compared with healthy controls (figure 4C,F). BALF exosomes from patients with sarcoidosis also showed strong signals for HLA-DR and HSP70 (figure 4C), with significantly higher levels on BALF exosomes from patients with sarcoidosis compared with healthy controls (figure 4E,G).

Bronchoalveolar lavage fluid (BALF) exosomes from patients with sarcoidosis contain neuregulin-1 (NRG1). (A) Silver staining analysis of BALF exosomes (healthy controls n=7, sarcoidosis n=14). (B) Western blot analysis of NRG1 44 kDa isoform and HLA-DR expression on exosomes purified on sucrose gradient. (C) Western blot analysis of the expression of NRG1 40 kDa isoform, HLA-DR (healthy, n=7, sarcoidosis, n=7), NRG1 44 kDa isoform, HSP70 and β-actin (healthy, n=5, sarcoidosis, n=7) on BALF exosomes. Densitometric quantification of (D) 40 kDa NRG1, (E) HLA-DR, (F) 44 kDa NRG1 (p=0.268) and (G) HSP70 on western blot analysis was measured and normalised to loading control (β-actin). Significant differences between groups were assessed using the Mann–Whitney test: *p<0.05, ***p<0.0001. AU, arbitrary units.
Protein identifications in bronchoalveolar lavage fluid exosomes by mass spectrometry of bands (10, 20 and 50 kDa) appearing stronger in patients with sarcoidosis by silver staining
Induction of cytokine production from PBMC by BALF exosomes
Sarcoidosis is known as a Th1-mediated disease with IFNγ production in the lungs24 and increased levels of IL-13 have also been reported.25 We therefore investigated whether BALF exosomes could stimulate autologous PBMC to secrete IFNγ as well as IL-13 in vitro measured by ELISPOT. BALF exosomes from patients with sarcoidosis stimulated significantly higher levels of IFNγ and IL-13 production in PBMC compared with BALF exosomes from healthy individuals (figure 5A,B). In addition, IFNγ spots from exosome stimulation in patients with sarcoidosis were much larger than those obtained with BALF exosomes from healthy individuals. Neither stage of disease (figure 5A–D) nor disease onset (data not shown) correlated with cytokine production.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Bronchoalveolar lavage fluid (BALF) exosomes from patients induce interferon-γ (IFNγ) and interleukin (IL)-13 in autologous peripheral blood mononuclear cells (PBMCs) and IL-8 in epithelial cells. Incubation of exosomes for 48 h corresponding to 6 ml of original volume of BALF per 500 000 autologous PBMCs. Co-incubations were performed on (A) anti-IFNγ (sarcoidois n=15, healthy n=9) or (B) anti-IL-13 (sarcoidois n=13, healthy n=9) coated ELISPOT plates. Assays were performed in triplicates and results are expressed as mean spots per 3×105 PBMCs. (C) IL-8 production by 16HBE14o- cells was measured by ELISA (pg/ml) after 48 h of co-incubation with exosomes from 6 ml original BALF volume or (D) 4 μg/ml of exosomal protein from patients with sarcoidosis (n=9) or healthy individuals (n=9). Results are expressed as the mean of duplicates. Production of IL-8 by unstimulated epithelial cells is indicated by the dotted line. Different symbols represent different stages of disease as indicated. Significant differences between groups were assessed using unpaired t test (B–D) or the Mann–Whitney test (A). **p <0.001, ***p<0.0001.
BAL exosomes induce the production of proinflammatory IL-8 in lung epithelial cells (16HBE14o-) and autologous PBMC
We further measured levels of IL-8 production by the lung epithelial cell line 16HB14o- and autologous PBMCs after co-incubation with exosomes from healthy individuals or patients with sarcoidosis. Preliminary results showed that BALF exosomes could trigger significant production of IL-8 in autologous PBMCs compared with unstimulated PBMCs, with no significant difference between the groups (sarcoidosis patients, n=7; healthy individuals, n=4; data not shown). Epithelial cells also produced IL-8 in response to BALF exosomes. Interestingly, exosomes from patients with sarcoidosis were more potent inducers of IL-8 release than those from healthy individuals, both when adding exosomes from the same original BALF volume (figure 5C) and the same exosomal protein (figure 5D).
Discussion
This study shows for the first time that exosomes might be of clinical relevance in a human respiratory disease by potentiating inflammation. We show that BALF exosomes from patients with sarcoidosis are quantitatively, phenotypically and functionally different from exosomes from healthy individuals. Electron microscopy data showed increased numbers of exosomes in the BALF from patients with sarcoidosis as well as an enrichment of MHC, tetraspanins, HSP70 and NRG1.
Exosomes enriched in HSP70 have previously been shown to be immunostimulatory in vivo26 and might potentiate inflammation. The strong correlation seen between HLA-DR and the tetraspanins (CD9, CD63 and CD81) is explained by the fact that MHC class II is an interaction partner to several members of the tetraspanin family.27 The negative correlation between HLA-DR and MUC1 in patients with sarcoidosis could be explained by the increase in exosomes from infiltrating cells and thereby a dilution of MUC1-expressing exosomes, likely to derive from epithelial cells.
One major question is whether or not the production of exosomes is beneficial for the patient with sarcoidosis. Exosomes could play a role in the clearance of an underlying infection prior to the onset of sarcoidosis by potentiating the anti-pathogen immune response. Alternatively, exosomes could contribute to the onset of sarcoidosis by exposing self-antigens on their MHC molecules, thus activating autoimmune T cells. We have previously shown in the mouse model that B cell activation is needed for an exosome-mediated T cell response,28 and that exosomes can carry B cell epitopes. B cell aggregates have been found in sarcoidosis pulmonary lesions.29 Thus, a role for exosomes in the activation of B cell responses in sarcoidosis is a possibility and could in turn lower the threshold for T cell activation. We propose that the increase in exosome production seen in patients is primarily a result of increased infiltration of cells in the lung. This is supported by the strong correlation seen between the concentration of cells and exosomal protein (figure 1F). However, other factors such as the state of cellular activation or a primary defect in patients could also contribute to the exosomal profile in sarcoidosis. Comparison with an infectious disease with similar levels of cell infiltration such as tuberculosis would resolve some of these questions. Either way, due to the capacity of these exosomes to stimulate cytokine responses, it is likely that they contribute to the inflammatory response in sarcoidosis.
Interestingly, we found NRG1 to be highly upregulated on exosomes from patients with sarcoidosis. NRG1 has not previously been associated with sarcoidosis but has frequently been implicated in cancer.17 30 It also serves as a pro-survival and pro-differentiation signal for epithelial cells and induces branching of the developing lung.31 To our knowledge, this is the first time NRG1 has been detected in sarcoidosis, and an overexpression of NRG1 could have a role in the pathogenesis of sarcoidosis—for example, by promoting cell survival and proliferation. It is noteworthy that sarcoidosis has been associated with an increased incidence of cancer in affected organs.32 However, a true connection between sarcoidosis and cancer development remains uncertain.
The T cell population in patients with sarcoidosis is skewed1 2 and only certain clones are activated, suggesting an antigen-specific reaction. Since exosomes from patients induce Th1-like cytokines, such as IFNγ, we suggest that exosomes also contribute to inflammation. However, they could also induce IL-13, previously shown to be elevated in BALF cells and PBMCs from patients with sarcoidosis.25 Whether IL-13 has pro- or anti-inflammatory properties in the lung in patients with sarcoidosis remains an unsettled issue.
We cannot rule out the possibility that exosomes have an immune regulatory role in the lung in sarcoidosis. The modest increase in CD86 compared with MHC class II in exosomes from patients with sarcoidosis suggests a reduced co-stimulatory ability in patients. Even though we could stimulate PBMCs from patients in vitro by these exosomes, it is possible that they act in a different way in vivo. The composition of leucocyte subsets in the lung differs from PBMC preparations, and BALF exosomes might be targeting other cells in vivo such as other DC populations. This interaction might therefore result in a different immunological effect such as induction of Th or T regulatory cells. In fact, sarcoidosis has been linked to clinical anergy to common environmental antigens such as Candida, possibly due to a reduced antigen presentation function of myeloid DC (mDC).33
The mechanism by which BALF exosomes induce IL-8 production in epithelial cells is probably different from that in PBMCs. Mycobacteria have been suggested as causative agents of sarcoidosis and have also been shown to be involved in the activation of epithelial cells.34 One might speculate that exosomes carrying pathogen-derived molecules such as proteins, RNA or DNA might induce activation through pattern recognition receptors. Alternatively, exosomes from patients might carry pro-inflammatory cytokines. Recent data show that exovesicles from human alveolar epithelial cells can carry TNFα with the ability to induce IL-8 production in epithelial cells.35
In view of the number of statistical tests carried out, results which were significant at the 5% level should be interpreted cautiously. However, we feel it is unlikely that these results are due to chance alone because, in the case of HLA-DR expression, we detected significant differences between the groups both by FACS and western blotting analysis (figure 2B and 4E). Furthermore, because HLA-DR is known to be associated with members of the tetraspanin family which is supported by our correlation data (figure 3A), it is likely that the difference in tetraspanin expression seen between groups (figure 2D,G) is truly significant.
In conclusion, this study shows that patients with sarcoidosis have increased production of exosomes in their lungs with the ability to stimulate cytokine production from autologous PBMCs and epithelial cells, suggesting both an adaptive and an innate mode of activation. These results suggest that exosomes from the BALF of patients with sarcoidosis might have a role in the pathogenesis of the disease and could be of relevance for the development of novel treatment strategies for sarcoidosis and other lung diseases.
What is already known on this topic
Exosomes are nano-sized immunomodulatory vesicles of endosomal origin previously found in the BALF of healthy individuals. These exosomes contain both major histocompatibility complex and co-stimulatory molecules and might have a regulatory role in the immune defence in the lung. Sarcoidosis is a Th1-driven inflammatory disease mainly affecting the lungs.
What this study adds
The results have implications for the use of exosomes as disease markers or targets for clinical treatment of lung diseases. We show that the BALF of patients with sarcoidosis is enriched in exosomes and that these exosomes have increased levels of immunostimulatory molecules such as MHC I and II, CD54 and tetraspanins, as well as NRG1 previously implicated in cancer progression. Exosomes from patients can induce IFNγ, IL-13 and IL-8 production, suggesting exosome-mediated activation of both the innate and adaptive immune system. These findings show the clinical relevance of exosomes as inducers of inflammation and suggest their importance in the pathogenesis of sarcoidosis.
Acknowledgments
We thank the Proteomics and Mass Spectrometry Unit at the Protein Analysis Center at the Karolinska Institutet for helping with mass spectrometry analysis. We are also grateful to Kjell Hultenby for helping with the electron microscopy.
References
Footnotes
Linked articles 138438.
KRQ and PTP share first authorship of the paper.
Funding This study was supported by grants from the Karolinska Institutet, the Swedish Medical Research Council (no 15242-05-2), the Torsten and Ragnar Söderbergs Foundation, the Stockholm County Council, the Mats Kleberg Foundation, the King Oscar II Jubilee Foundation, the Swedish Cancer Foundation (no. 08 0299), the Swedish Heart-Lung Foundation, the Swedish Society of Medicine, the Centre for Allergy Research Karolinska Institutet, the Swedish Cancer and Allergy Foundation and the Swedish Asthma and Allergy Association's Research Foundation.
Competing interests None.
Ethics approval This study was conducted with the approval of the local ethics committee of Northern Stockholm.
Provenance and peer review Not commissioned; externally peer reviewed.