Article Text

Abstract
Background: Intestinal epithelial cells secrete exosome-like vesicles. The aim of this study was to characterise murine intestinal epithelial exosomes and to analyse their capacity to inform the immune system in vivo in mice.
Methods: Epithelial exosomes were obtained from the murine epithelial cell line MODE K incubated in the presence or absence of interferon γ (IFN-γ) together with pepsin/trypsin ovalbumin hydrolysate (hOVA) to mimic luminal digestion. Exosomes isolated from MODE K conditioned media (EXO-hOVA and EXO-hOVA-IFN) were characterised by western blot, peptide mapping, and mass spectrometry. They were injected intraperitoneally to C3H/HeN mice to test their immunocompetence.
Results: MODE K epithelial exosomes displayed major histocompatibility complex (MHC) class I and class II (upregulated by IFN-γ) molecules and tetraspan proteins (CD9, CD81, CD82) potentially involved in the binding to target cells. A33 antigen, an Ig-like molecule highly specific for intestinal epithelial cells, was enriched in exosomes and was also found in mice mesenteric lymph nodes, suggesting exosome migration towards the gut associated lymphoid tissues. Intraperitoneal injection of EXO-hOVA or EXO-hOVA-IFN did not induce humoral or cellular tolerance to OVA in mice. In contrast, exosomes obtained after incubation with IFN-γ (EXO-hOVA-IFN), bearing abundant MHC class II/OVA complexes, induced a specific humoral immune response.
Conclusions: Epithelial exosomes are antigen presenting vesicles bearing MHC class II/peptide complexes that prime for an immunogenic rather than tolerogenic response in the context of a systemic challenge. In the intestine, both the mucosal microenvironment and local effector cells are probably key players in determining the outcome of the immune response to exosome derived epitopes.
- exosomes
- mucosal immunity
- food antigen
- intestinal epithelium
- antigen transport
- A33 antigen
- IFN-γ, interferon γ
- APC, antigen presenting cell
- IEC, intestinal epithelial cells
- OVA, ovalbumin
- hOVA, ovalbumin hydrolysate
- MHC, major histocompatibility complex
- MALDI-TOF-MS, matrix assisted laser desorption ionisation-time of flight-mass spectrometry
- DMEM, Dulbecco’s modified Eagle’s medium
- MLN, mesenteric lymph nodes
- SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis
- CFA, complete Freund’s adjuvant
- DTH, delayed type hypersensitivity
- PBS, phosphate buffered saline
Statistics from Altmetric.com
- IFN-γ, interferon γ
- APC, antigen presenting cell
- IEC, intestinal epithelial cells
- OVA, ovalbumin
- hOVA, ovalbumin hydrolysate
- MHC, major histocompatibility complex
- MALDI-TOF-MS, matrix assisted laser desorption ionisation-time of flight-mass spectrometry
- DMEM, Dulbecco’s modified Eagle’s medium
- MLN, mesenteric lymph nodes
- SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis
- CFA, complete Freund’s adjuvant
- DTH, delayed type hypersensitivity
- PBS, phosphate buffered saline
The high concentration of foreign antigens present in the gut lumen is separated from the local intestinal immune system by a single layer of polarised epithelial cells. Intestinal epithelial cells (IEC) can endocytose dietary proteins, process them into peptides,1 and in vitro can present these peptides to CD4+ T lymphocytes, particularly when major histocompatibility complex (MHC) class II molecules are upregulated,2–4 and in the presence of accessory molecules.5,6 However, in vivo, direct interactions between IEC and CD4+ T cells predominating in the lamina propria are limited due to the presence of the basement membrane. IEC have been shown to release exosome-like vesicles7 morphologically similar to the exosomes secreted by professional antigen presenting cells (APC) such as B cells8 or dendritic cells.9 Epithelial exosomes released from basolateral sides of IEC bear high amounts of MHC class I and class II molecules, suggesting that they may interact with the underlying immune system. They also contain A33 antigen,7 a molecule whose expression is restricted essentially to the intestinal epithelium.10,11 Exosomes derived from professional APC moderately activate CD48 or CD812 T lymphocytes, and it was recently shown that their antigen presenting capacity highly depended upon dendritic cells,14 suggesting that exosomes have to be taken up by professional APC to induce efficient T cell activation. They may therefore be involved in the rapid dissemination of immune information via APC.
In the present study, we tested the hypothesis that intestinal epithelial exosomes bearing MHC class II/ovalbumin (OVA) peptide complexes, generated in the presence of interferon γ (IFN-γ) and OVA that had been predigested with gastric and pancreatic enzymes, can transmit immune information and thereby act as sensors of the antigenic information present in the intestinal lumen. Murine IEC derived exosomes were characterised and their interaction with the immune system was tested in vivo in mice. We tested the possibility that intraperitoneal injection of OVA loaded epithelial exosomes could either: (1) mimic oral ingestion of OVA and inhibit specific humoral and/or cellular immune responses (oral tolerance), as previously suggested15 or (2) directly prime the humoral immune response to OVA (sensitisation).
METHODS
Antigens
We incubated epithelial cells in the presence of OVA that had been predigested with pepsin and trypsin to mimic the fate of this protein in the intestinal lumen. OVA 10 mg/ml was treated for four hours (37°C, pH 1.8) with pepsin (20 mg/g protein; Sigma, St Louis, Missouri, USA) and trypsin (20 mg/g protein) at pH 7.8. Enzymes were inactivated for 45 minutes at 85°C and eliminated by ultrafiltration on membranes retaining molecules greater than 10 000 Da (centriplus, YM10; Millipore, St Quentin en Yuelines, France). The filtrate, referred to as ovalbumin hydrolysate (hOVA), was sterilised and frozen until use.
Murine intestinal epithelial cell line (MODE K)
The murine intestinal epithelial cell line MODE K,16 kindly provided by D Kaiserlian, was used. This cell line is derived from C3H/He mice and expresses I-Ak MHC class II molecules. Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Life Technologies, Cergy-Pontoise, France) containing 10% heat inactivated fetal calf serum, 4 mM glutamine, and 50 µg/ml gentamicin, in a humidified atmosphere of 5% CO2 /95% air.
Exosome preparation from MODE K epithelial cell line conditioned media
MODE K cells were seeded (5×106 cells) into 75 cm2 Falcon flasks in DMEM containing 10% Prolifix (synthetic serum; Biomedia, France), 4 mM glutamine, 50 µg/ml gentamicin, and OVA (10 µM) or OVA predigested with pepsin and trypsin to mimic degradation in the intestinal lumen (hOVA corresponding to 10 µM OVA). Four days later, cell supernatants were collected to isolate exosomes under basal conditions (EXO-OVA or EXO-hOVA). To obtain exosomes from MHC class II enriched enterocytes, MODE K cells were further incubated for 48 hours with OVA or hOVA in the presence of murine IFN-γ (100 U/ml) before exosomes were collected (EXO-OVA-IFN or EXO-hOVA-IFN). Exosomes obtained under basal and IFN-γ conditions were isolated and quantified as previously described.7 Generally, 10 µg MODE K exosomes were collected from the 48 hour conditioned medium from a 75 cm2 culture flask.
In some experiments, exosomes were isolated from human or mice serum to analyse whether intestinal epithelial exosomes can access the peripheral blood stream. Serum exosomal protein content was estimated by western blot analysis using the A33 antigen, a specific marker of intestinal epithelial exosomes.
Identification of epithelial exosomes in mouse mesenteric lymph nodes
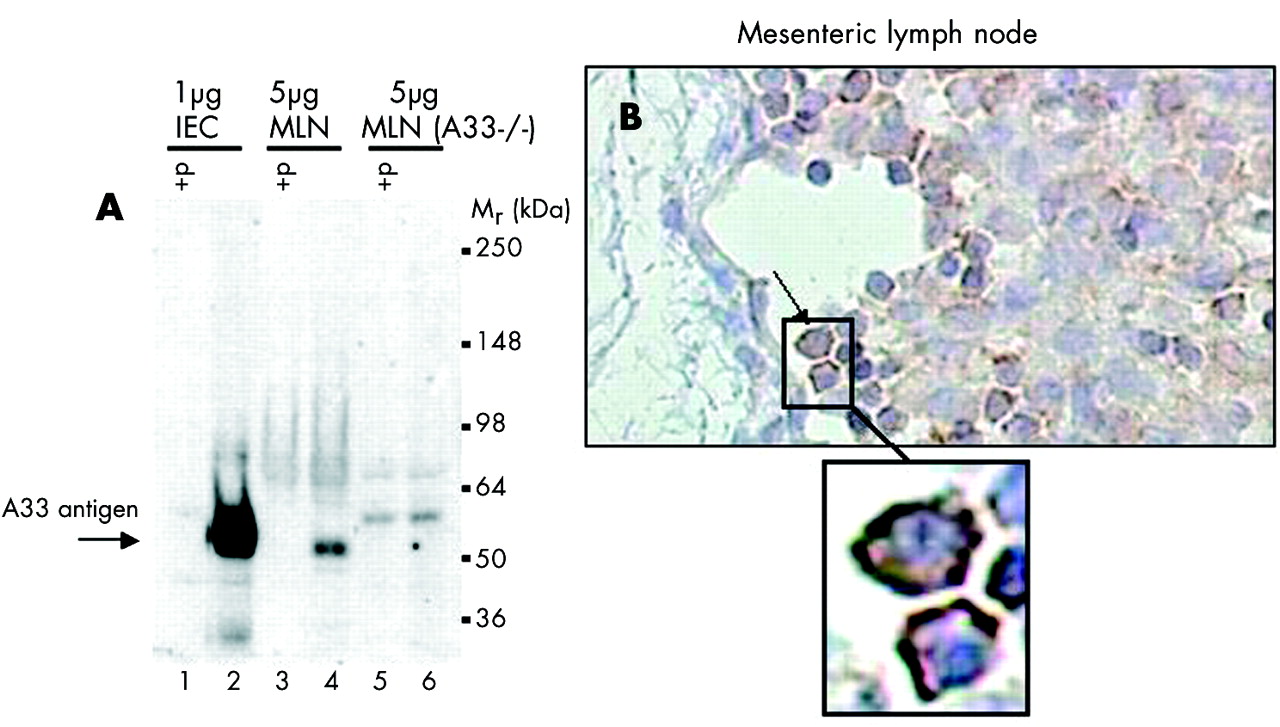
We looked for the presence of A33 antigen in mesenteric lymph nodes (MLN) to test the hypothesis that epithelial exosomes released at the basal pole of enterocytes can migrate (alone or bound to dendritic cells) towards these secondary lymphoid organs. As A33 antigen is highly expressed in IEC,11 a comparison between MLN and freshly isolated mouse IEC was performed. IEC or MLN were homogenised in 1% Triton X100 in 10 mM HEPES buffered saline, pH 7.4, containing a protease inhibitor cocktail. The lysates were incubated for 30 minutes at 4°C and ultracentrifuged at 100 000×g for one hour. Partially purified proteins were then prepared using anion exchange high pressure liquid chromatography by injecting the resultant supernatant onto a Mono-Q HR10/10 column at pH 7.8, and by eluting the proteins from the column using a linear 0–1 M NaCl gradient. The A33 antigen (molecular weight ∼50 kDa) in eluant fractions was detected by western blot analysis using polyclonal rabbit anti-mA33antigen IgG (1.2 mg/ml). Controls to confirm the specificity of the protein detection included MLN harvested from A33 antigen deficient mice (A33−/−; Tebbutt NC, Ernst M, and Heath JK, unpublished data) and preabsorption of the anti-mA33 antigen antibodies with the immunising A33 antigen C terminal peptide.11 Immunohistochemical detection of the murine A33 antigen was also performed on sections of paraffin embedded MLN from normal mice using the same rabbit polyclonal antibody.
Characterisation of MODE K exosomes by western blot analysis
MODE K cells were lysed over 30 minutes at 4°C in 50 mM Tris-HCl, pH 8, 150 mM NaCl, 1% EDTA, and 0.5% Triton X100, containing a protease inhibitor cocktail and centrifuged for 10 minutes at 700 g (4°C) to remove nuclei. The supernatants were ultracentrifuged for one hour at 100 000 g (4°C). Supernatants containing whole cell proteins were adjusted to 2 mg/ml in phosphate buffered saline (PBS), diluted (1/2) in sample buffer (2×), and stored at −20°C.
Exosomal proteins were separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) in 10% acrylamide gels. Gels were transferred onto nitrocellulose membranes (Amersham Pharmacia Biotech, Amersham Biosciences Europe, Saclay, France) in 10 mM Tris, 0.2 M glycine, and 30% methanol. Non-specific binding sites were blocked by incubation in Tris-HCl, pH 7.6, containing 5% bovine serum albumin and 0.1% Tween. Blots were incubated for one hour with mouse anti-I-A (mouse/rat) antibodies (clone MRC OX-6; Serotec Cergy-St Christophe, France) to detect MHC class II molecules and with peroxidase conjugated sheep antimouse Ig (Amersham) (1:20 000).
A33 antigen was detected with polyclonal rabbit antimouse A33 antigen antibodies in non-reducing conditions.11 These antibodies were detected using peroxidase conjugated goat antirabbit IgG (Biorad Ivry sur Seine, France) (1:15 000), followed by ECL (Amersham Pharmacia Biotech).
MODE K derived exosomes: peptide mapping analysis using matrix assisted laser desorption ionisation-time of flight-mass spectrometry (MALDI-TOF-MS)
Identification of proteins present in MODE K derived exosomes was analysed using MALDI-TOF-MS. MODE K derived exosomes (20 µg), prepared in sample buffer, were run on 10% SDS-PAGE and stained with Coomassie blue. The major bands were excised from the gel and in-gel digested with trypsin. The digest solution was analysed by MALDI-TOF-MS using a Bruker Biflex mass spectrometer (Bruker-Franzen Analytik Bremen, Germany), as previously described.17 Monoisotopic peptide masses were assigned and used in database searches. Typical search parameters, using the Microsoft FIT program, were as follows: maximum allowed peptide mass error of 100 ppm; consideration of one incomplete cleavage per peptide; no restriction was placed on the isoelectric point of the protein; and a protein mass range of 0–200 kDa was allowed.
In vivo studies in mice
We tested the capacity of MODE K epithelial exosomes to induce specific tolerance or sensitisation in mice.
Oral tolerance (humoral response)
Fifty two C3H/HeN mice (three week old females) were divided into six experimental groups comprising 5–10 mice each.
Standard oral tolerance protocol
The first two groups comprised tolerised and sensitised mice, obtained using standard protocols of oral tolerance. OVA tolerised mice (TOL) received 20 mg OVA in 200 µl 0.2 M sodium bicarbonate by intragastric gavage using a stainless steel blunt feeding needle. OVA sensitised mice (SENS) received bicarbonate alone by gastric gavage. One week later (day 8), all mice were immunised: they received a subcutaneous injection (100 µl), to the base of the tail, of 25 µg OVA/complete Freund’s adjuvant (CFA, v/v) and a second injection (100 µl) of 10 µg OVA in PBS two weeks later. One week after the second injection, blood was collected for anti-OVA IgG and IgE measurements.
Induction of tolerance by epithelial exosomes
To test for the tolerogenic capacity of exosomes, four additional groups were tested in parallel with the first two groups. In these groups, OVA loaded exosomes were administered intraperitoneally with the aim of mimicking oral administration of OVA. EXO-OVA mice received, instead of OVA gavage, an intraperitoneal injection of 10 µg exosomes (100 µl) derived from the MODE K cell line cultured in the presence of OVA under basal conditions. EXO-OVA-IFN mice received an intraperitoneal injection of 10 µg OVA-exosomes derived from the MODE K cell line cultured in the presence of OVA and 100 U/ml IFN-γ. In some experiments, hOVA was used instead of intact OVA to produce exosomes from MODE K cells incubated in the presence or absence of IFN-γ. Mice in these groups received an intraperitoneal injection of 10 µg exosomes and are referred to as EXO-hOVA and EXO-hOVA-IFN, respectively. After one week, all mice were immunised as stated above and blood was collected for anti-OVA IgG and IgE measurements.
Oral tolerance (DTH response)
Tolerised (TOL) and sensitised (SENS) mice were given OVA or bicarbonate buffer, respectively, by gastric gavage, as described above (see humoral response section above). The other mice received intraperitoneal OVA exosomes instead of OVA by gavage: EXO-hOVA mice received an intraperitoneal injection of 10 µg exosomes derived from MODE K cells fed hOVA under basal conditions and EXO-hOVA-IFN mice received an intraperitoneal injection of 10 µg exosomes derived from MODE K cells fed hOVA in the presence of IFN-γ. After one week, all mice were injected subcutaneously (base of the tail) with 100 µg OVA/CFA. Two weeks later, mice received a subcutaneous injection of 30 µl OVA (1 µg/µl) in the ear and ear swelling was measured as the difference in ear thickness before and 24 hours after OVA injection using a micrometre calliper (Oditest, Kröplin, Germany) to quantify the delayed type hypersensitivity (DTH) response.
Sensitisation protocol (humoral response)
To analyse whether intestinal epithelial exosomes could directly prime the immune system, mice received an intraperitoneal injection of exosomes. EXO-hOVA mice received an injection of 10 µg exosomes (100 µl) derived from MODE K cells fed hOVA in the absence of IFN-γ whereas EXO-hOVA-IFN mice received an injection of 10 µg exosomes derived from MODE K cells fed hOVA in the presence of IFN-γ. A control group received an intraperitoneal injection of 100 µl PBS instead of exosomes. After one week, all mice groups were injected subcutaneously with 25 µg OVA/CFA in 100 µl. One week later mice were bled and anti-OVA IgG and IgE antibody responses were measured. In another experiment, the capacity of OVA loaded epithelial exosomes and free OVA peptides to stimulate the immune system were compared: 0.2 µg of pepsin/trypsin ovalbumin peptides (hOVA), calculated as being equivalent to the amount loaded on 10 µg epithelial exosomes in IFN-γ conditions, were injected intraperitoneally in order to prime the mice before the subcutaneous OVA boost.
Measurement of OVA specific IgE and IgG antibodies (ELISA)
IgG and IgE titres in plasma were measured using an enzyme linked immunosorbent assay as follows: 96 well plates (Immulon II, Labsystems, France) were coated overnight at 4°C with 100 µl of 25 µg/ml OVA (Sigma) in PBS. After washes with PBS-0.05% Tween 20 (PBS-Tween), non-specific binding sites were blocked for one hour at 37°C with 100 µl/well of PBS-1% bovine serum albumin. The plates were then incubated for two hours at 37°C with 100 µl plasma samples serially diluted in PBS-Tween. Peroxidase conjugated rat antimouse IgG (Serotec) diluted 1:1000 in PBS-Tween were added (100 µl/well) and incubated for two hours at 37°C. Finally, 100 µl of citrate buffer, pH 4.0, containing 10 mg ortho-phenylenediamine (Sigma) and H2O2 (Sigma) were added to each well. The reaction was stopped with 50 µl/well 6N H2SO4 and plates were read at 450 nm using an automatic Multiskan microplate reader (Labsystems, Cergy-Pontoise, France).
Statistical analysis
Statistical analysis was performed using SAS software. Results are expressed as mean (SD). Comparison of means was performed using analysis of variance and non-parametric Wilcoxon tests. The general linear model procedure was also used for multiple group to group comparisons. Differences were considered significant when p<0.05.
RESULTS
Characterisation of MODE K intestinal epithelial exosomes
Two molecules found in human epithelial exosomes,7 namely MHC class II molecules and A33 antigen, were analysed by western blot. As shown in fig 1A, MHC class II molecules were not detectable in MODE K cell lysates, even in the presence of IFN-γ. MHC class II molecules were detectable in MODE K derived exosomes harvested under basal conditions and their expression was markedly upregulated in the presence of IFN-γ, indicating substantial enrichment in epithelial exosomes. The discrepancy between expression of MHC class II molecules in entire cells and exosomes may be due to the antibodies differentially recognising the mature peptide bound class II molecules possibly present on exosomes compared with nascent class II forms mainly present in epithelial cells. A33 antigen was detected in cell lysates and was highly enriched in exosomes although its expression in exosomes was not upregulated by IFN-γ (data not shown).

Expression of major histocompatibility complex (MHC) class II molecules and A33 antigen in MODE K cells incubated in the presence or absence of interferon γ (IFN-γ), and in MODE K cell derived exosomes. Although MODE K cell lysates did not express detectable levels of MHC class II molecules, even in the presence of IFN-γ, MODE K derived exosomes expressed detectable levels under basal conditions, which were markedly upregulated in the presence of IFN-γ. A33 antigen, a molecule expressed specifically by intestinal epithelial cells, was enriched in exosome preparations compared with whole cell lysates.
Peptide mapping by MALDI-TOF-MS analysis
The results of peptide mapping of MODE K derived exosomes are summarised in table 1. Several protein families were observed: (a) proteins playing a role in the biogenesis and release of exosomes, such as integrin precursors and cytoskeleton proteins under basal and inflammatory (+IFN-γ) conditions and Ras related protein (Rab-2, Rab-7); (b) proteins responsible for directing exosomes to their target cells, such as MFG-E8, a protein previously found in dendritic cell derived exosomes,18 and heat shock proteins hsp71 and hsp90 that may play a role in the transfer of antigenic peptides and binding to dendritic cells, and in their maturation, as recently described in mast cell derived exosomes19; (c) proteins potentially involved in antigen presentation (MHC class I molecules) or in exosome function (tetraspan molecules CD81, CD82, and undescribed transmembrane four superfamily protein); and (d) cytoplasmic proteins probably not related to exosome function which may have been non-specifically captured into the exosome lumen during their formation (histones, enzymes).
Peptide mapping of MODE K derived epithelial exosomes using matrix assisted laser desorption ionisation-time of flight-mass spectrometry (MALDI-TOF-MS) analysis
Epithelial exosomes do not induce tolerance in mice
Humoral tolerance
An earlier study suggested that epithelium derived exosomes might induce oral tolerance in rats.15 In contrast, in our murine model, MODE K derived exosomes bearing MHC class II/OVA molecules and injected intraperitoneally did not suppress IgE and IgG antibody responses (fig 2A, B). As expected, anti-OVA antibodies were significantly lower in tolerised (TOL) mice (60 and 3950 for IgE and IgG titres, respectively) than in sensitised (SENS) (480 and 192 000, for IgE and IgG titres; p<0.001) mice, but Ig titres observed in EXO-OVA or EXO-OVA-IFN mice were not significantly different from those observed in SENS mice, indicating that epithelial exosomes did not tolerise mice to OVA. Furthermore, anti-OVA antibody titres were significantly higher in EXO-hOVA-IFN mice than in SENS mice (IgE 6.15×103v 0.48×103 (p<0.0001); and IgG 900×103v 192×103 (p<0.0001)), suggesting that exosomes may sensitise rather than tolerise mice, at least in our experimental conditions.

Ovalbumin (OVA) loaded epithelial exosomes, injected intraperitoneally, did not abrogate humoral immune responses. Anti-OVA IgG and IgE titres obtained in C3H/HeN mice were orally tolerised to OVA (TOL), OVA sensitised (SENS), or injected intraperitoneally with exosomes derived from MODE K cells fed intact OVA (EXO-OVA) or pepsin-trypsin hydrolysed OVA (EXO-hOVA), or from MODE K cells treated with interferon γ (IFN-γ) (EXO-OVA-IFN and EXO-hOVA-IFN). Not only did exosomes not induce specific humoral tolerance, but on the contrary, EXO-OVAh-IFN and EXO-OVA increased sensitisation to OVA. *p<0.05 compared with tolerised mice; †p<0.03 compared with sensitised mice (n = 5–16 mice per group).
DTH tolerance
As shown in fig 3, the cellular response (DTH) was decreased in orally tolerised mice compared with sensitised mice (28 v 89 µm; p<0.05) as expected, but not in mice that received 10 µg EXO-hOVA or EXO-hOVA-IFN by intraperitoneal injection (89 and 98 µm, respectively), indicating that epithelial exosomes administered intraperitoneally did not tolerise the cellular immune response to OVA.

Ovalbumin (OVA) loaded epithelial exosomes injected intraperitoneally did not abrogate the cellular immune response (delayed type hypersensitivity), as measured by the difference in ear thickness before and after local injection of OVA or pepsin-trypsin hydrolysed OVA (EXO-hOVA), or from MODE K cells treated with interferon γ (IFN-γ) (EXO-hOVA-IFN). *p<0.05 compared with tolerised (TOL) mice (n = 5–8 mice per group). SENS, sensitised mice.
Epithelial exosomes can prime mouse humoral responses
IEC derived exosomes, under our experimental conditions, were unable to induce tolerisation of mice towards OVA and even seemed to enhance the sensitisation process. As shown in fig 4, exosomes derived from MODE K cells stimulated by IFN-γ in the presence of hOVA (EXO-hOVA-IFN) primed a specific anti-OVA humoral immune response. This response was not observed with exosomes prepared in the absence of IFN-γ (EXO-hOVA) that express fewer or non-functional MHC class II/OVA peptide complexes. These results suggest that IFN- promotes the release of functional epithelial exosomes bearing MHC II/OVA-peptide complexes able to stimulate the immune system. In addition, immunisation of mice with 0.2 µg of OVA peptides (calculated as being equivalent to the amount expressed on 10 µg epithelial exosomes under IFN-γ conditions), with or without CFA, did not induce a significant IgG response. This result highlighted the increased efficiency of peptide loaded epithelial exosomes in stimulating the immune system compared with free peptides.

Epithelial exosomes can prime the humoral immune response in mice. Anti- ovalbumin (OVA) IgG and IgE titres obtained in C3H/HeN mice injected intraperitoneally (ip) with phosphate buffered saline (PBS) or exosomes obtained from MODE K cells exposed to pepsin-trypsin OVA hydrolysate (hOVA) and incubated in the presence or absence of interferon γ (IFN-γ) (EXO-hOVA and EXO-hOVA-IFN) before subcutaneous (sc) injection of OVA/complete Freund’s adjuvant (CFA). Exosomes obtained in the presence of IFN-γ (thus bearing more MHC II/OVA peptide complexes) were capable of inducing specific IgG and IgE responses. The amount of hOVA peptides corresponding to that loaded on 10 µg EXO/hOVA/IFN (that is, 0.2 µg) did not induce an IgG response, even in the presence of CFA. IgE responses to 0.2 µg OVA peptides did not parallel IgG responses, suggesting differential susceptibilities to low dose antigens (n = 4–9 mice per group). *p<0.001 compared with the other groups; †compared with EXO-hOVA (p<0.02), PBS (p<0.08), and hOVA (p<0.02).
Mouse epithelial exosomes are present in mesenteric lymph nodes but not in serum
As IEC derived exosomes seem to act as vehicles of the antigenic information originating from the gut lumen, we searched for their presence in mice MLN. We used A33 antigen as a specific marker of epithelial exosomes to detect their presence in the lymph nodes draining the small intestine. As shown in fig 5A, MLN lysates contained A33 antigen, suggesting the presence of IEC derived exosomes in these secondary lymphoid organs. The amount of A33 antigen detected in MLN was much lower than that observed using lysates from purified IEC. The A33 antigen band was specific, as preabsorption of the immune serum with A33 antigen immunising peptide led to the disappearance of the band, and A33−/− mice did not display any specific bands. An immunohistochemical study of the localisation of A33 antigen in MLN (fig 5B) suggested that A33 antigen was enriched on the external membranes of immune cells entering the MLN in the subcapsular sinus. However, on a cautionary note, while the presence of A33 antigen in MLN might be a reliable indicator of the presence of epithelial exosomes, an alternative explanation would be that the A33 antigen was present as a result of a subpopulation of dendritic cells transporting apoptotic intestinal epithelial cells in phagocytic vacuoles to T cell areas of MLN.20 The apparent distribution of A33 antigen on the external membranes of the immune cells in the MLN (rather than in apoptotic bodies) would tend to argue against this possibility.

(A) Western blot analysis of A33 antigen in mice mesenteric lymph node (MLN) lysates showing significant expression of A33 antigen and suggesting that A33 antigen, a marker of intestinal epithelial cells (IEC), also expressed in exosomes, can reach the draining lymph nodes. The western blot was performed under reducing conditions using a rabbit polyclonal anti-mA33 antigen IgG (1.2 mg/ml). Proteins were extracted from mouse IEC (1 µg protein: lanes 1 and 2), MLN (5 µg protein: lanes 3 and 4), and MLN from A33 antigen deficient mice (A33−/−) (5 µg protein: lanes 5 and 6). In lanes 1, 3, and 5, the analysis was conducted after preabsorption of the anti-mA33 antigen IgG with 0.75 mg/ml (∼50×molar excess) of the immunising mA33 antigen peptide (+P). (B) Immunohistochemistal analysis of MLN from C3H/HeN mice, showing immune cells entering the subcapsular sinus and displaying the A33 antigen at their surface (see magnified inset), suggesting that epithelial exosomes may reach MLN via migration of antigen presenting cells. It cannot be excluded however that the A33 antigen immunoreactivity is due to the presence of intestinal epithelial apoptotic bodies, although enrichment of the staining around the periphery of the cells argues against this explanation.
We also searched for the presence of intestinal epithelial exosomes at the systemic level by analysing human and mice serum derived exosomes for their content in A33 antigen. We were unable to detect significant amounts of A33 antigen in serum derived exosomes (data not shown) suggesting that free intestinal epithelial exosomes do not achieve significant concentrations in the circulation.
DISCUSSION
The present results show that murine IEC stimulated with IFN-γ in vitro release exosomes bearing MHC class II molecules/peptide complexes that, when administered intraperitoneally to mice, induce a specific immune response.
The murine intestinal epithelial cell line MODE K16 produced exosomes displaying a pattern of molecule expression similar to that previously described in human intestinal epithelial exosomes, suggesting that these molecules play an important role in exosome biogenesis and function. The presence of the tetraspan proteins CD9, CD81, and CD82 confirms their endosomal origin and MHC class II molecules, upregulated by IFN-γ, underline their potential to carry dietary protein derived peptides. The heat shock proteins hsp71 and hsp90 could play a role in the transfer of peptides from IEC to professional APC. Indeed, hsps have been involved in peptide exchange with MHC class II molecules after internalisation of hsp-peptide complexes through the CD91 receptor present on APC, and their role in cross presentation by dendritic cells of antigen associated exosomes from mast cells has recently been described.19 Hsps may also induce maturation and migration of dendritic cells to the draining lymph nodes.21 Epithelial exosomes were also shown to bear A33 antigen, a molecule that is highly specific for intestinal epithelial cells, and MFG-E8,22,23 a molecule that binds to dendritic cell integrins that could play a role in adhesion of exosomes to their target cell. It is noteworthy that no costimulatory molecules were found on epithelial exosomes.
In the present study, we tried to reproduce the array of OVA peptides produced in the digestive tract in vivo by mimicking gastric and pancreatic hydrolysis by pepsin and trypsin. The resulting OVA peptides were placed in contact with the intestinal epithelial cell line, MODE K, allowing further uptake and processing of OVA peptides by IEC prior to their loading on MHC class II molecules, especially in the presence of IFN-γ. It is generally accepted that antigen presentation at the intestinal level is tolerogenic, at least under basal conditions.13,24 In the present study, OVA loaded MODE K exosomes were used in a tolerisation protocol, in order to check whether their intraperitoneal administration could mimic the tolerising effect induced by an oral dose of OVA. In contrast with results previously reported on epithelial tolerosomes,15 intraperitoneal injection of OVA loaded epithelial exosomes in mice did not induce a tolerogenic humoral or cellular immune response to OVA. The fact that EXO-OVA-IFN and EXO-hOVA-IFN generated different responses (hOVA having greater potential to sensitise mice) suggests that processing of food antigens by epithelial cells may lead to the formation of distinct peptide epitopes.
The discrepancy between our results and those previously reported on epithelial tolerosomes15 is difficult to explain although the two studies were conducted using different epithelial cell lines and different animal models.
As we did not observe any tolerogenic effect of epithelial exosomes delivered through the intraperitoneal route, we tested whether they might, per se, induce a specific immune response to OVA. We found that MODE K derived exosomes loaded with OVA peptides in the presence of IFN-γ were capable of inducing a strong OVA specific immune response when administered intraperitoneally. The absence of response with OVA exosomes obtained in the absence of IFN-γ suggested that a certain density of MHC class II molecules on the exosomal surface is mandatory to the immune response. Alternatively, MHC class II molecules present on exosomes in the absence of IFN-γ have been described as nascent rather than mature MHC class II forms25; these immature nascent forms have been suggested to induce tolerogenic rather than immunogenic responses.
Our results indicate that epithelial exosomes can induce a significant immune response to OVA despite their lack of costimulatory molecules. This would indicate that they would most probably have to interact with professional APC expressing these molecules in order to induce an immune response, as previously demonstrated with dendritic cell derived exosomes.14 Indeed, our results are in agreement with those recently reported in mice on dendritic cell derived exosomes which, after injection, can generate an antigen specific MHC class II restricted CD4+ T cell proliferation and upregulation of activation markers. This activation mainly depends on costimulatory molecules expressed by dendritic cells, and not those present on exosomes.14 In the same way, mast cell derived exosomes also need dendritic cells to efficiently activate T cells in vitro,26 suggesting that MHC class II-peptide complexes are transferred from exosomes to dendritic cells. The interactions between APC and exosomes was also demonstrated with B cell derived exosomes which were shown to bind specifically to follicular dendritic cells in germinal centres.27 In our study, intestinal epithelial exosomes also seem to play a role as MHC/peptide carriers. We artificially injected these epithelial exosomes intraperitoneally, which is not the site (intestinal mucosa) where they are released normally. We hypothesise (fig 6) that epithelial exosomes may allow the luminal antigens taken up and processed by epithelial cells to be transferred from the intestinal lumen, via epithelial exosomes, to the local intestinal dendritic cells “sensing” the content of the gut lumen, and to the MLN. Indeed, A33 antigen, a specific marker of epithelial cells11 highly enriched on epithelial exosomes, was detected in lysates from MLN. This antigen sampling pathway via exosomes could add to the previously described direct sampling, by dendritic cells, of bacteria present in the intestinal lumen, through the epithelial tight junctions.28 In the intestinal suppressive environment (transforming growth factor β), the immune recognition of exosome derived peptides by mucosal lymphocyte populations, either directly (without costimulation) or indirectly through professional APC, might drive a tolerogenic response in the gut, the “natural” environment for exosomes, at least under physiological conditions. Thus effector cells that interact with exosome derived epitopes, together with the microenvironment in which this occurs, are probably key factors in determining the outcome of the immune response, the same determinants triggering different responses within different sites. In infectious conditions, maturation of dendritic cells in the presence of “danger signals” (lipopolysaccharide, TNF-α),29 as well as the increase in MHC class II/peptide density on epithelial exosomes, may explain an abnormal immune response to luminal antigens. This hypothesis requires further investigation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Working hypothesis on the role of epithelial derived exosomes. Exosomes released at the basolateral surfaces of enterocytes and loaded with luminal antigen derived peptides cross the basement membrane to reach the lamina propria (1) where they may interact with dendritic cells (DCs) (2). After maturation of exosome loaded DCs, antigen presentation may take place locally (3), or epithelial exosomes/DCs may migrate to the draining lymph nodes (4). IEC, intestinal epithelial cells; MLN, mesenteric lymph nodes.
In summary, our results suggest that epithelial exosomes may serve as messengers sensing the antigenic content of the gut lumen. Transmission of immune information via epithelial exosomes to professional APC present in the intestinal mucosa may be one of the pathways involved in the immune response to intestinally delivered antigens.
Acknowledgments
G van Niel was a recipient of a grant from the Fondation pour la Recherche Médicale.
REFERENCES
Footnotes
-
↵* J Mallegol and C Bevilacqua contributed equally to this work