Article Text

Abstract
Background: Repeated daily exposure of healthy human subjects to NO2 induces an acute airway inflammatory response characterised by neutrophil influx in the bronchial mucosa
Aims: To assess the expression of NF-κB, cytokines, and ICAM-1 in the bronchial epithelium.
Methods: Twelve healthy, young non-smoking volunteers were exposed to 2 ppm of NO2/filtered air (four hours/day) for four successive days on separate occasions. Fibreoptic bronchoscopy was performed one hour after air and final NO2 exposures. Bronchial biopsy specimens were immunostained for NF-κB, TNF-α, eotaxin, Gro-α, GM-CSF, IL-5, -6, -8, -10, -13, and ICAM-1 and their expression was quantified using computerised image analysis.
Results: Expression of IL-5, IL-10, IL-13, and ICAM-1 increased following NO2 exposure.
Conclusion: Upregulation of the Th2 cytokines suggests that repeated exposure to NO2 has the potential to exert a “pro-allergic” effect on the bronchial epithelium. Upregulation of ICAM-1 highlights an underlying mechanism for leucocyte influx, and could also explain the predisposition to respiratory tract viral infections following NO2 exposure since ICAM-1 is a major receptor for rhino and respiratory syncytial viruses.
- nitrogen dioxide
- bronchial epithelium
- cytokines
- ICAM-1
- Th2 response
- BHR, bronchial hyperresponsiveness
- CV, coefficient of variation
- FEV, forced expiratory volume
- GMA, glycol methacrylate
- ICAM, intercellular adhesion molecule
- IL, interleukin
- NF, nuclear factor
- TNF, tumour necrosis factor
- ROI, reactive oxygen intermediates
Statistics from Altmetric.com
- BHR, bronchial hyperresponsiveness
- CV, coefficient of variation
- FEV, forced expiratory volume
- GMA, glycol methacrylate
- ICAM, intercellular adhesion molecule
- IL, interleukin
- NF, nuclear factor
- TNF, tumour necrosis factor
- ROI, reactive oxygen intermediates
Nitrogen dioxide (NO2) is formed as a result of a chemical reaction between nitrogen and oxygen during high temperature combustion and constitutes an important air pollutant. It poses a major problem to human health both in the indoor as well as in the outdoor atmosphere. Peak levels of up to 0.4 parts per million (ppm) are encountered in the outdoors, particularly along kerbsides in downtown areas with heavy motor vehicular traffic.1 Peak levels in the indoor environment can reach up to 4 ppm in garages, ferries, skating ice rinks, and kitchens with gas cookers.1
Several epidemiological studies have shown strong links between exposure to high levels of NO2 and morbidity and mortality of asthma.1 In addition, several studies have also shown a link between personal exposure to NO2 and predisposition to childhood respiratory tract viral infections.1–3 The precise mechanisms underlying this latter biological link are not well understood but studies in animal models have shown that NO2 exposure could impair some local defence mechanisms in the airways.4
Short term exposure to 0.4–2 ppm NO2 can also induce modest bronchoconstriction and non-specific bronchial hyperresponsiveness (BHR) in healthy and asthmatic airways.1 We have shown previously that acute exposure to 2 ppm NO2 can induce an acute inflammatory response in healthy human airways.5,6 We have also shown that repeated daily exposure to NO2 (2 ppm for four hours/day for four successive days) can induce a decrement in FEV1 that returns to baseline by day 4, but the acute inflammatory response characterised by PMN influx and activation still persists after the final exposure.6 The precise mechanisms contributing to the acute inflammatory response are not well understood. Nitrogen dioxide is an oxidant pollutant and it has been suggested that it induces an oxidative damage to cell membranes resulting in the generation of reactive oxygen intermediates (ROIs).7 Bronchial epithelium is a metabolically active barrier and is the first structure in the human airways that encounters any noxious agent. Devalia and coworkers have reported that NO2 exposure could damage the bronchial epithelium, decrease ciliary beat frequency, and increase cell permeability.8 In addition, in vitro studies have shown that NO2 exposure induces secretion of several NF-κB mediated cytokines (GM-CSF, IL-8, TNF-α) and ICAM-1 in the bronchial epithelium.9,10
In the present study we examined the expression of NF-κB, ICAM-1, cytokines including GM-CSF, IL-5, -6, -10, -13, eotaxin, and TNF-α in the bronchial epithelium of healthy human airways following repeated daily exposure to 2 ppm NO2. We hypothesised that repeated daily exposure to peak concentration of NO2 would upregulate the expression of NF-κB, ICAM-1, and the cytokines in the bronchial epithelium and contribute to the acute inflammatory response. Using archived bronchial biopsy specimens from our previous study6 we examined the expression of these biomarkers in the bronchial epithelium using immunohistochemistry and computerised image analysis.
METHODS
Subjects
Twelve healthy volunteers were recruited (eight men, four women; mean age 26 years, range 21–32). The subjects had negative skin prick tests and normal lung function, with no history of asthma or other respiratory disease. All were non-smokers and had had no airway infection for six weeks prior to or during the study. Subjects were not allowed any non-steroidal anti-inflammatory drugs or vitamin supplements.
Nitrogen dioxide and air exposure
Exposures were performed in an aluminium exposure chamber in the university hospital of North Sweden in Umeå. During each four hour exposure, light exercise (75 W) on a bicycle ergometer was alternated with rest, in 15 minute intervals. The air and NO2 exposures (four hours/day for four successive days) were performed in random order three weeks apart.
The Ethics Committee of the University of Umeå approved the study and informed consent was obtained.
Bronchoscopy
Fibreoptic bronchoscopy was performed as previously described.5
Immunohistochemistry
Biopsy specimens were processed into glycol methacrylate (GMA) as previously described11 and stored at −20°C until stained.
Sequential sections (2 μm) were cut and stained with toluidine blue to assess the morphology and select the best specimen for immunostaining.
Sections were stained using monoclonal antibodies (table 1) as previously described11 and developed with streptavidin biotin-peroxidase complex (Dako, High Wycombe, UK), yielding a brown reaction product. Sections were counterstained with Mayer’s haematoxylin.
Names and sources of monoclonal antibodies used for immunohistochemical analysis
Image analysis
Sections were examined by computerised image analysis with colour vision 1.7.6 software (Improvision, Birmingham, UK) on an Apple Macintosh computer. The area of epithelium, containing the staining, was defined interactively. The amount of staining was then determined as a fraction of total epithelial area. This procedure was repeated for all the epithelium within the section and the percentage of positive staining calculated.
RESULTS
Intra-observer variability in quantification of biomarkers in the bronchial epithelium using computerised image analysis
One slide showing a positively stained epithelium for each biomarker was selected randomly and computerised image analysis was performed on four separate days. In order to eliminate bias due to tiredness or fatigue of the observer, the different biomarkers were studied according to a randomisation code obtained from the website Research Randomizer (http://www.randomizer.org). The mean of the four observations, together with the standard deviation (SD) was calculated and the coefficient of variation (CV) was expressed as [SD/mean] × 100 (<10% for biomarkers studied, data not shown).
Immunohistochemistry
A total of 19 biopsy specimens were available for this study; toluidine blue staining revealed that 17 (eight pairs) were technically suitable for immunostaining, based on morphology of the epithelium. Positive staining for IL-5, IL-6, IL-8, IL-10, IL-13, Gro-α, NF-κB, and ICAM-1 was detected. There was also positive staining detectable for TNF-α, GM-CSF, and eotaxin, but the majority of specimens stained negatively for these biomarkers.
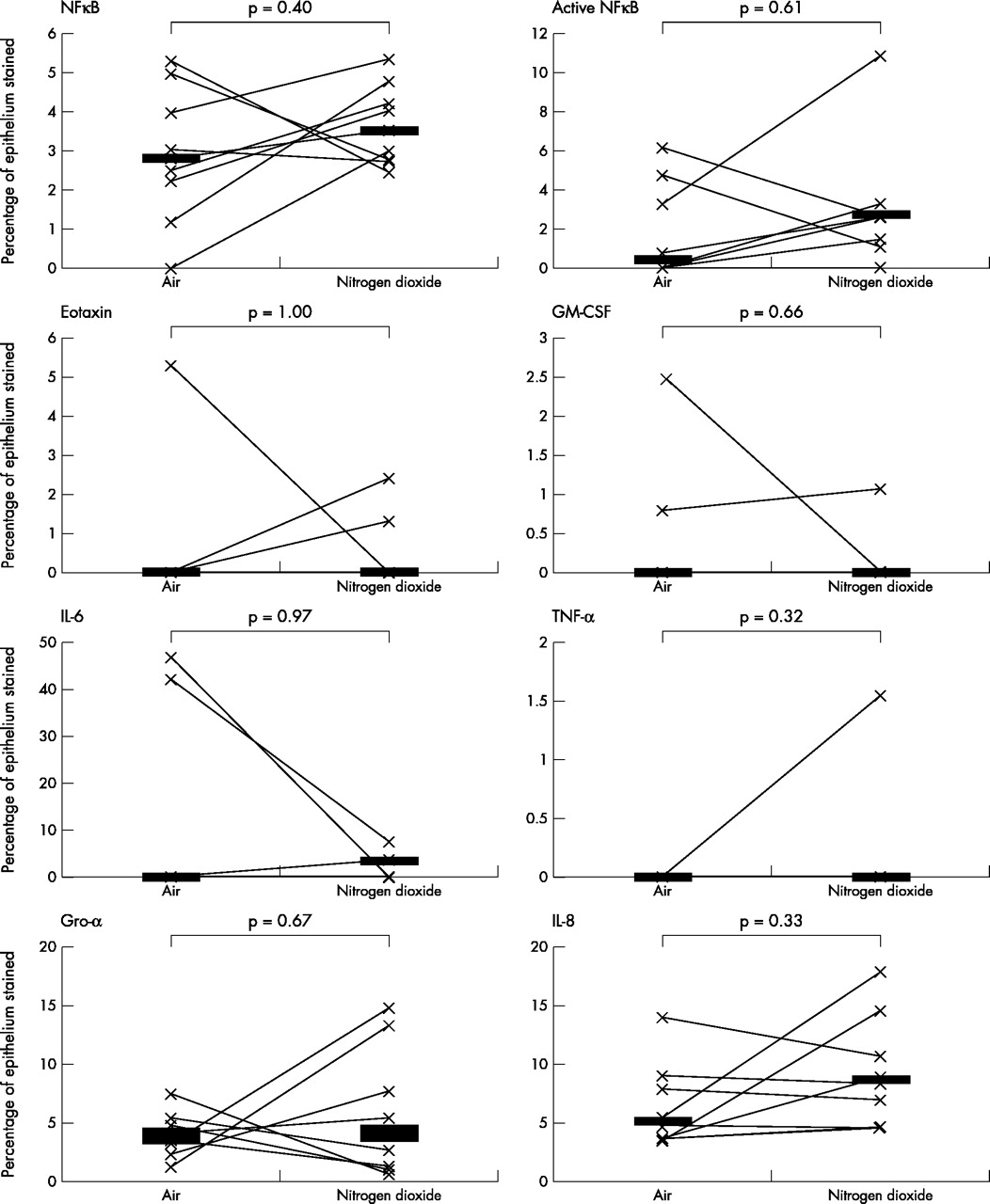
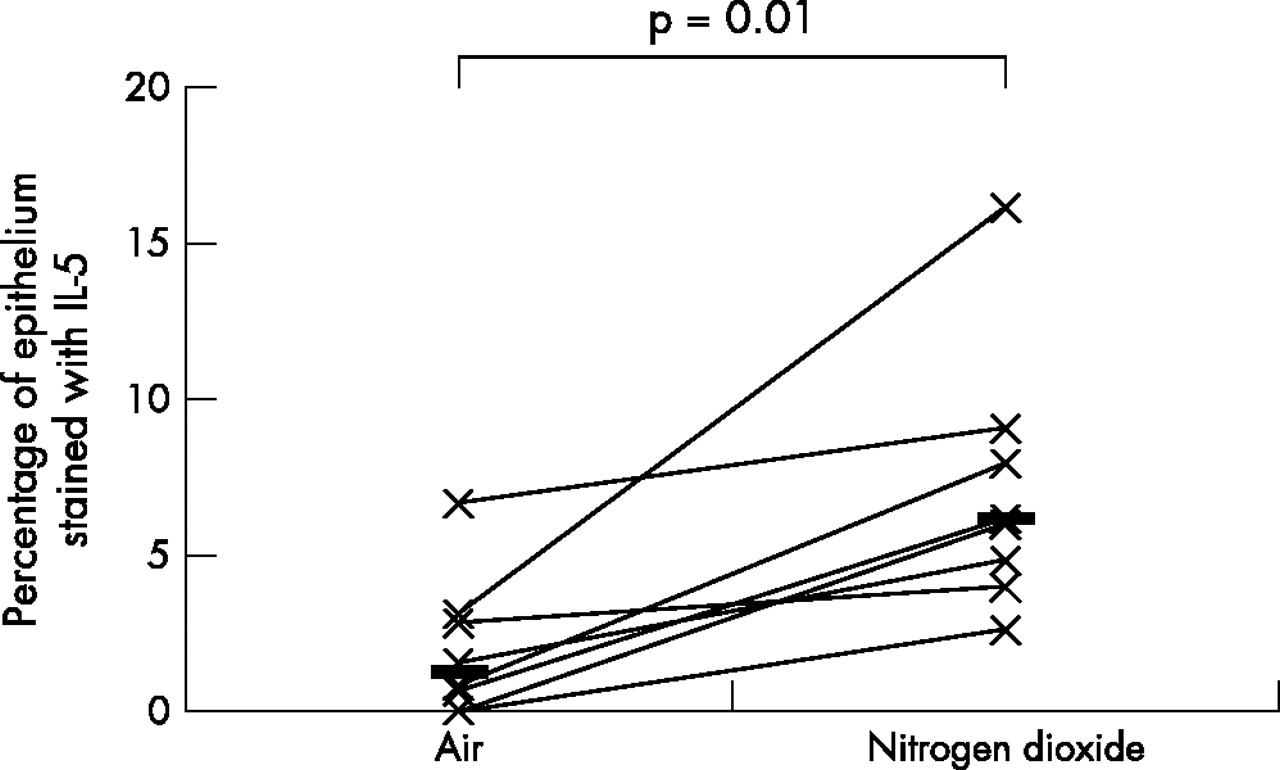
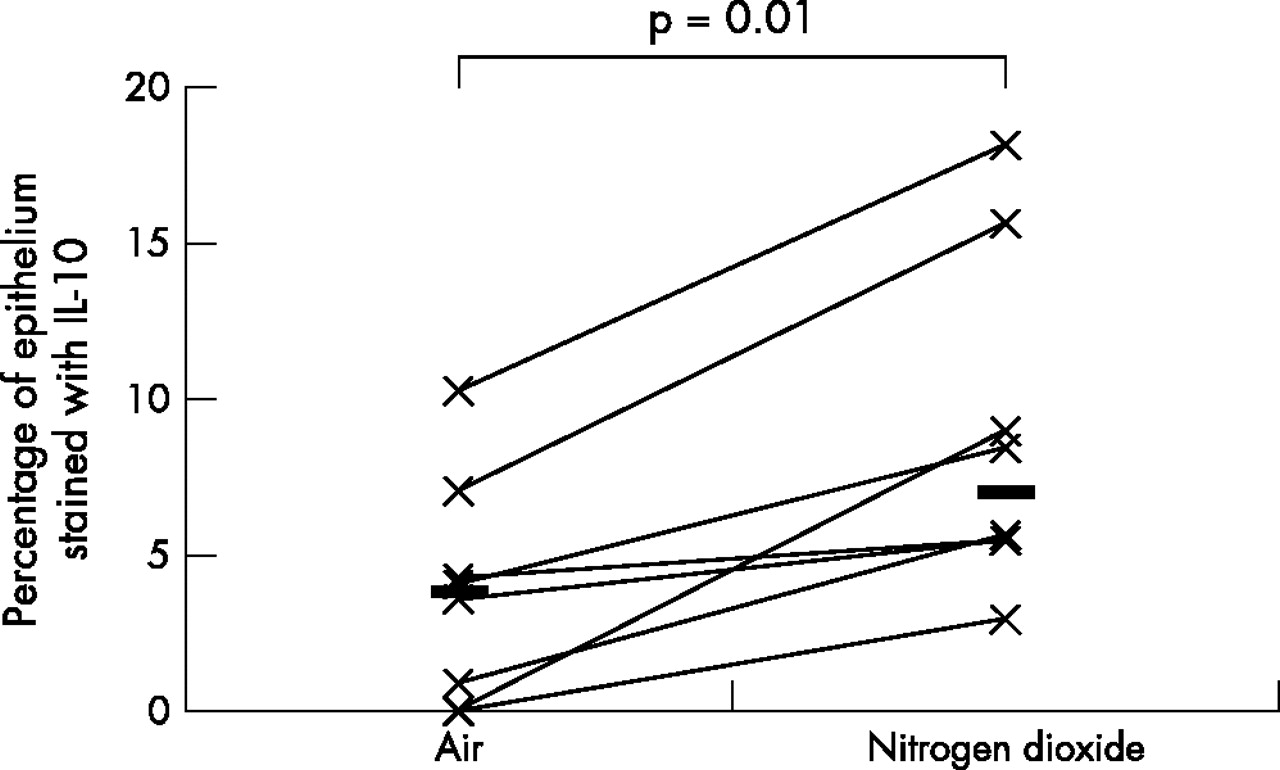
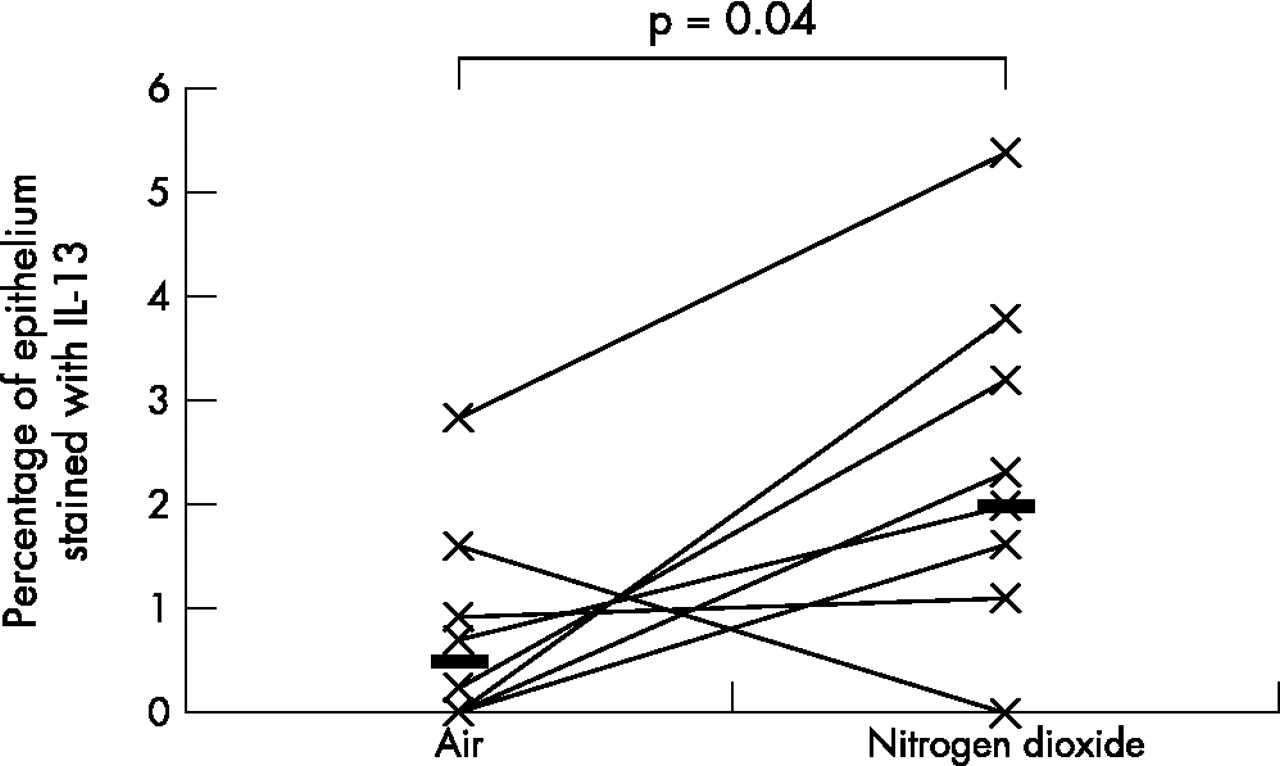
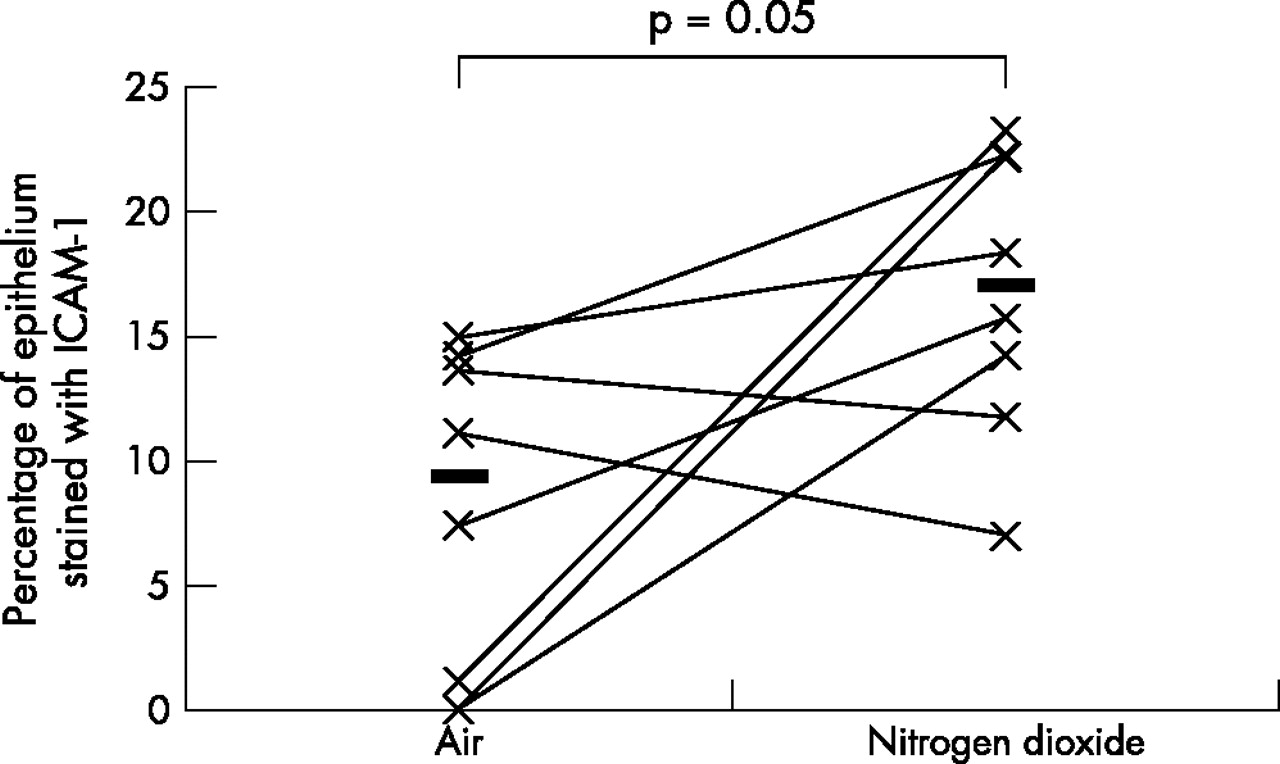
As the data was not normally distributed, Wilcoxon’s matched paired sign rank test was used to compare the differences between various biomarkers on the two exposure days; a p value of <0.05 was considered statistically significant. A fivefold median increase in the expression of IL-5 (p = 0.01) and IL-13 (p = 0.04), twofold median increase in the expression of ICAM-1 (p = 0.05), and a twofold median increase in the expression of IL-10 (p = 0.01) was observed in the bronchial epithelium following repeated daily exposure to NO2 (table 2, figs 1–4). No significant changes were seen in the other biomarkers (table 2, fig 5).
Quantification of biomarkers in the bronchial epithelium
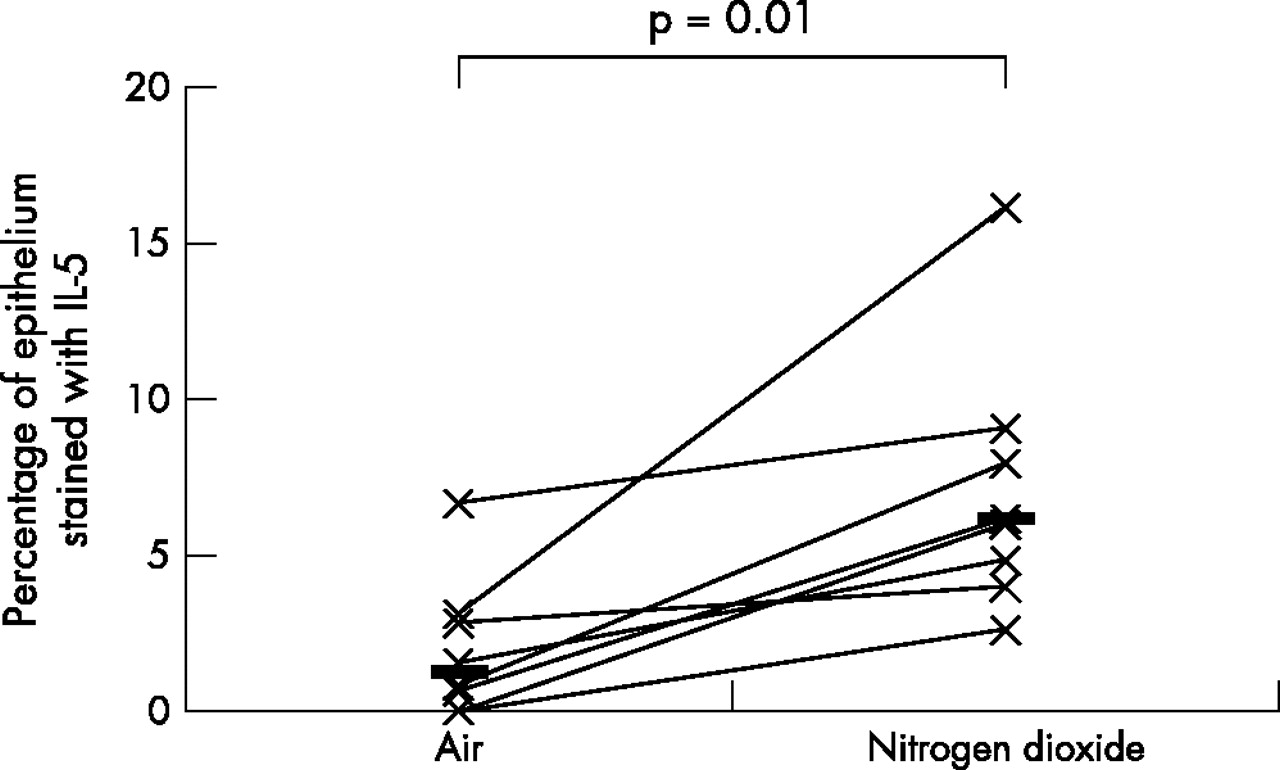
Comparison of bronchial epithelial expression of IL-5 between the two exposures. Horizontal lines represent medians.
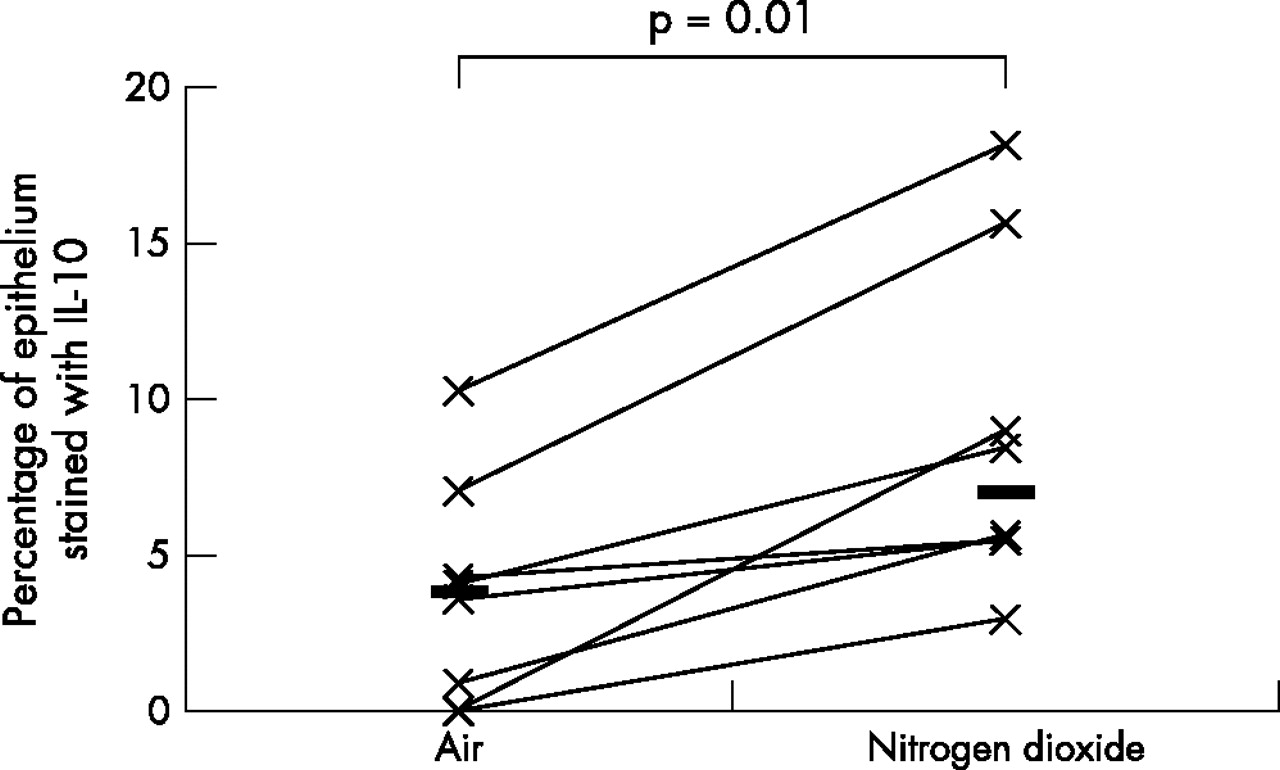
Comparison of bronchial epithelial expression of IL-10 between the two exposures. Horizontal lines represent medians.

Comparison of bronchial epithelial expression of IL-13 between the two exposures. Horizontal lines represent medians.
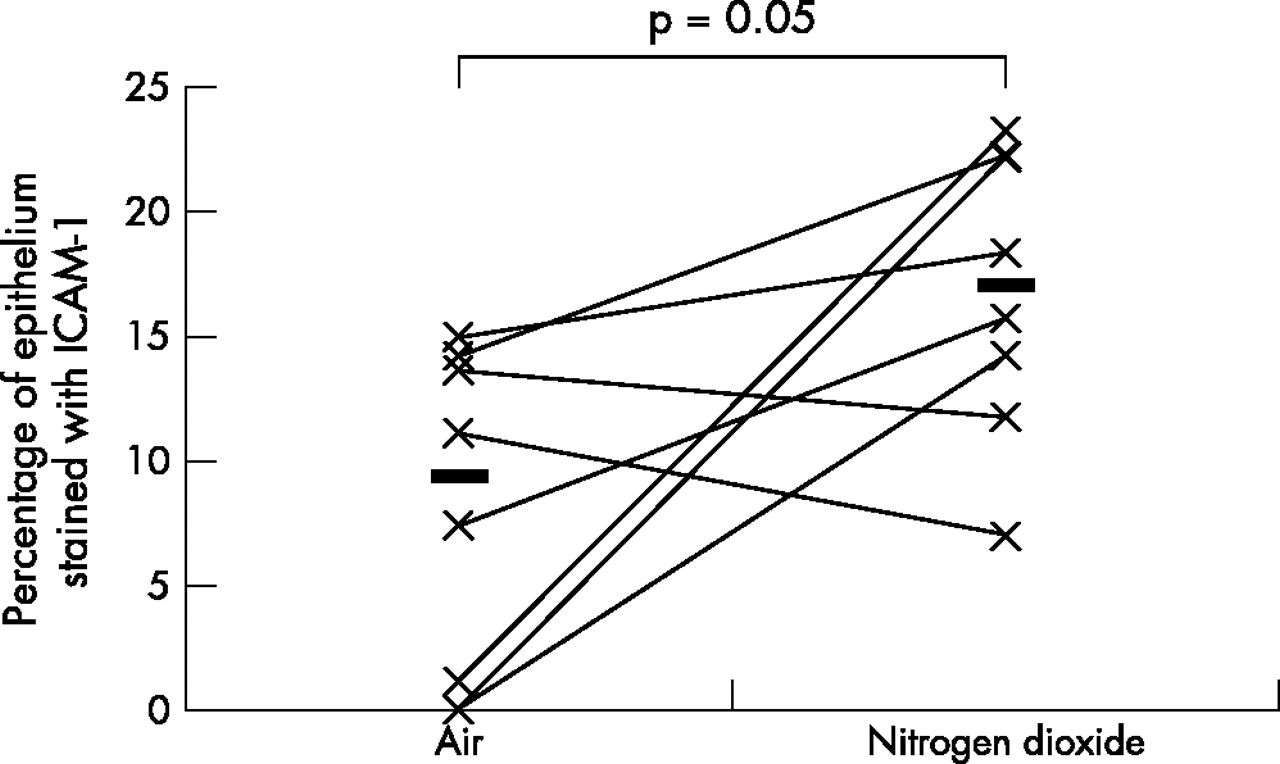
Comparison of bronchial epithelial expression of ICAM-1 between the two exposures. Horizontal lines represent medians.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comparison of bronchial epithelial expressions of other biomarkers between the two exposures. Horizontal lines represent medians.
DISCUSSION
This is the first in vivo study examining the effects of repeated NO2 exposure on the bronchial epithelium of healthy human airways. This study has shown that repeated exposure of healthy human airways to 2 ppm of NO2 induces an upregulation of IL-5, -10, -13, and ICAM-1 in the bronchial epithelium. Upregulation of these cytokines suggests that repeated exposure to peak indoor levels of NO2 induces a bias of the epithelial cytokine expression towards an “allergic” or Th2 phenotype.
We fixed our biopsy specimens in acetone and processed the tissue in GMA. This method has been shown to preserve the tissue morphology and immunoreactive epitopes and has been used successfully to study the expression of inflammatory cells, cytokines, and leucocyte-endothelial adhesion molecules in the bronchial mucosa.11 We were able to detect clear albeit weak constitutive expression of ICAM-1, NF-κB, IL-5, -6, -10, -13, Gro-α, eotaxin, and IL-8 in the bronchial epithelium. Using computerised image analysis we were able to objectively quantify the expression of these biomarkers. Intra-observer variability in quantification showed a CV of less than 10% for biomarkers supporting the reliability and reproducibility of this method of quantification.
The upregulation of IL-5, -10, and -13 suggests a Th2 epithelial response to repeated daily exposure to NO2. IL-5 is a pleiotropic cytokine with a major role in eosinophil chemoattraction, proliferation, differentiation, survival, and activation.12 IL-5 induces peripheral blood eosinophilia and activation when given by a nebuliser to patients with asthma,13 and it seems to play an important role in the pathogenesis of BHR in murine models of asthma.14,15
Several studies have highlighted an important role for IL-13 in the immunopathogenesis of asthma. Targeted pulmonary expression of IL-13 in a model of IL-13 knock out mice, led to several characteristic features of asthma, including influx of eosinophils, mononuclear cells, mucus cell metaplasia, airway fibrosis, eotaxin production, BHR, and bronchoconstriction.16 In addition, IL-13 can activate eosinophils and promote their survival. Another important property of IL-13 is to induce isotype switching of B cells for IgE synthesis.17 Bronchoscopy studies have shown significantly increased expression of IL-13 mRNA in the bronchial mucosa of atopic asthmatics as opposed to atopic non-asthmatics and healthy controls.18
IL-10 has been shown to possess immunoregulatory properties on airway inflammation and bronchial reactivity. In a mouse model of allergen sensitisation, IL-10 knock out mice showed profound inflammatory response to ovalbumin but failed to develop BHR.19 When the IL-10 gene was reinstated, development of BHR in response to ovalbumin was restored. IL-10 also downregulates the release of a broad range of cytokines from inflammatory cells.20,21 The bronchial epithelium constitutively expresses IL-10 and its expression has been shown to be downregulated in the bronchial epithelium of patients with cystic fibrosis.22 In support for a potential anti-inflammatory role for IL-10, reduced levels have been reported in the airways of smokers, and patients with chronic obstructive pulmonary disease and asthma as opposed to healthy controls.23 Thus, it is likely that increased expression of IL-10 in our study could represent either a physiological response of the epithelium to NO2 or a proinflammatory response in the context of BHR.
One of the limitations of our study is that we did not measure bronchial reactivity; clearly in the context of the present results this would have been important. The present study was carried out on archived bronchial biopsy specimens embedded in GMA and therefore it was not possible to study the mRNA expression or levels of the relevant cytokines in the specimens or bronchoalveolar lavage fluid respectively to further confirm our observations. Future studies should address these issues.
Adhesion molecules play an important role in acute and chronic inflammatory processes. ICAM-1 plays a key role in leucocyte trafficking in inflammatory responses and helps in transendothelial migration.24 In our previous study,6 we showed neutrophil influx in response to NO2; the upregulation of ICAM-1 seen in this study provides a plausible mechanism.
ICAM-1 is an important surface receptor for rhinovirus, a pathogen commonly implicated in upper respiratory tract infections.25,26 Recent studies have shown that ICAM-1 is also a receptor for human picorna27 and respiratory syncytial viruses.28 These viruses are major causes of respiratory tract infections in childhood; the upregulation of ICAM-1 in response to repeated NO2 exposure, highlights a plausible role for this adhesion molecule in explaining the link between personal exposure to NO2 and predisposition to respiratory tract virus infections in children. In a recent study in a cohort of 114 asthmatic children we have shown that personal exposure to NO2 increases the risk of PCR proven upper respiratory tract infection with picorna and respiratory syncytial virus infection.29 It follows that NO2, by upregulating the major receptor for these viruses (that is, ICAM-1), could make the airways more susceptible to infections. In support of this, Bianco and coworkers have reported that Th2 cytokines exert a dominant role on epithelial cell expression of ICAM-1.30 They have shown that co-incubation of H292 epithelial cells with Th2 cytokines increased ICAM-1 expression by fivefold; a further twofold increase in the expression was seen when the cells were previously infected with human rhino virus-14.30
No significant changes in NF-κB expression were seen. This does not necessarily mean that this transcription factor does not play any role in the inflammatory response to NO2. It is likely that upregulation of this transcription factor occurs early during the inflammatory process but does not persist.
In conclusion, repeated exposure of healthy human airways to 2 ppm NO2 induces an upregulation of Th2 cytokines including IL-5, -10, -13, and ICAM-1 in the bronchial epithelium. Upregulation of Th2 cytokines suggests that NO2 could exert a “pro-allergic” effect on the airways. Upregulation of ICAM-1 provides a plausible mechanism for neutrophil influx during the acute inflammatory response and predisposition of respiratory tract virus infections, following repeated NO2 exposure. Further studies are required to investigate these putative mechanisms, especially in asthmatics, so that a better understanding could pave the way for potentially new interventive therapeutic strategies.
Acknowledgments
The authors are grateful to Mr James Cameron for technical assistance.