


Abstract
Nociceptin is a novel neuropeptide of the opioid peptide family recently identified as the endogenous ligand of the opioid receptor-like “orphan” receptor. Unlike other opioids, nociceptin has hyperalgesic effects in vivo. In the present study, nociceptin was found to inhibit electrical field stimulation-induced tachykinergic contractions of the guinea pig isolated bronchus preparation. The threshold effect was about 1 nM, and at 0.1 μM, nociceptin inhibited contractions evoked by 5-Hz stimulation by more than 50%. This inhibitory effect was found to be mediated by a prejunctional mechanism involving none of the classical (μ, δ and κ) opioid receptors. Although the hypothesis that the effect of nociceptin was secondary to opioid receptor-like stimulation cannot be pharmacologically addressed, opioid receptor-like-receptor-mRNA was found to be expressed in the upper vagal sensory ganglion, where the cell bodies of the tachykinin-containing sensory neurons are located. Nociceptin immunoreactive nerve fibers in the airway wall, distinct from the tachykinin-containing fibers, were identified as an endogenous source of nociceptin. These data indicate that nociceptin may influence airway physiology by modulating tachykinergic neurotransmission.
Opioid peptides are a family of neuropeptides with modulatory functions on nociception and inflammation (see Olson et al., 1995). These effects are mediated through three known types of opioid receptors termed μ, δ and κ receptors. These receptors bind with high-affinity endorphins, enkephalins and dynorphin, respectively. By homology screening, an additional receptor with little affinity for the endogenous opioids has been identified in the murine and human brain (Mollereau et al., 1994) as well as in rat brain (Bunzowet al., 1994). This “orphan” receptor has been termed ORL1-receptor (Mollereau et al., 1994). Recently, the endogenous ligand of the ORL1-receptor has been discovered independently by two groups and termed orphanin FQ (Reinscheid et al., 1995) or, because of its hyperalgesic properties in vivo, nociceptin (Meunier et al., 1995). Pharmacological studies in vivo indicated that nociceptin exerts its effects predominantly in the CNS. The ORL1-receptor, accordingly, was found to be present in the neuropil of the neocortex and the hippocampus of the rat (Anton et al., 1996). However, Giuliani and Maggi (1996) showed that in the PNS, nociceptin affects mediator release from sensory nerves in the guinea pig renal pelvis.
In the airways, the relevance of the dual (afferent and efferent) functions of sensory nerves has been shown under both physiological and pathophysiological conditions. Local tachykinin release from airway sensory nerves produces symptoms of inflammation and thus has been related to the pathophysiology of the asthmatic reaction (Barnes, 1990). Allergic inflammation in the airways in turn changes the excitability and phenotype of afferent nerves in the airways, which then may contribute to airway disease (see Undem and Riccio 1997;Fischer et al., 1996).
Opioids inhibit EFS-induced tachykinergic contractions in the airway by a naloxone-sensitive mechanism (Bartho et al., 1987; Belvisiet al., 1990; Johansson et al., 1989; Matranet al., 1989). In the present study, we have investigated the effects of nociceptin on tachykinin release from these nerves and have attempted to identify the receptor involved. The guinea pig isolated bronchus preparation was used as a convenient experimental model to study tachykinergic transmission in the airway (see Ellis and Undem, 1994a). The endogenous source of nociceptin and its site of action were identified using immunohistochemistry and RT-PCR. The results support the hypothesis that, like other opioids, nociceptin inhibits tachykinergic transmission in the guinea pig airway. Unlike the effects of the classical opioids, however, the effect of nociceptin is not blocked by naloxone and is probably due to the stimulation of ORL receptors on jugular afferent neurons.
Materials and Methods
Functional studies.
Male Hartley guinea pigs (300–500 g) were asphyxiated in a CO2 chamber. After asphyxiation, the thorax was opened. The bronchi were isolated and placed in a dissection dish containing a modified Krebs’ buffer solution (composition in mM: 118 NaCl, 5.4 KCl, 1 NaH2PO4, 1.2 MgSO4, 1.9 CaCl2, 25 NaHCO3 and 11.1 glucose) that contained 1 μM atropine, 1 μM propranolol and 3 μM indomethacin (Sigma, St. Louis, MO) to block the effect of betaadrenergic receptor stimulation, muscarinic receptor stimulation and prostaglandins, respectively (Undem et al., 1990). The left or right main-stem bronchus was bisected into rostral and caudal rings, attached to a Grass FT03 force transducer and suspended under 1 g of tension in a 10-ml water-jacketed (37°C) organ chamber containing the aforementioned Krebs’ solution bubbled with 95% O2-5% CO2. The isometric tension was continuously recorded on a Grass (Warnick, RI) model 7 polygraph.
EFS.
The right or left bronchial rings were allowed to equilibrate for 1 h, the buffer being replaced with fresh solution at 15-min intervals. After the equilibration period, electrical field current was delivered to platinum electrodes, situated on opposite sides of the tissue, from a Grass S48 stimulator. The current was first passed through a StimuSplitter (Med Lab Instruments, Fort Collins, CO) for signal monitoring and amplification. The tissues were stimulated at 20 V, 1 ms, 5 Hz for 15 s. The stimulation parameters delivered approximately 200 to 400 mA of current to the electrodes. This is most suitable for inducing neural tachykinin-mediated contractions with this design. As expected (Ellis and Undem, 1994b), the EFS-induced contractions were abolished by 1 μM tetrodotoxin (n = 3) or by a combination of CP 9994 (1 μM) and SR 48968 (1 μM) (kindly provided by Zeneca Pharmaceuticals, Wilmington, DE). At the concentrations used, these compounds are selective antagonists of NK-1 and NK-2 receptors, respectively (Edmonds-Alt et al., 1992;McLean et al., 1993); therefore, these contractions are referred to as tachykinergic responses. The tissues were stimulated with EFS at 20-min intervals. The EFS-induced contraction always returned to the base-line level before the subsequent stimulation. The average response to the first two stimulations was taken as the control response.
Pharmacological treatments.
A concentration-response curve for nociceptin-induced inhibition of the EFS-induced response was obtained in a cumulative fashion (see the representative tracing in fig. 1). Five minutes after the second “control” response had reached base line, 1 nM nociceptin (Sigma) was added to the bath, and 15 min later the tissue was again subjected to EFS. This was repeated with 10 nM and 100 nM nociceptin in the same tissue. The response in the presence of nociceptin was then calculated as a percentage of the control response. Nociceptin inhibited the response in the left and right bronchi equivalently (P > .1), so the data for both bronchi were pooled. The inhibition by nociceptin (0.1 μM) in the left bronchus and the right bronchus averaged 64 ± 9% (n = 5) and 49 ± 8% (n = 6), respectively. In a separate series of experiments, vehicle (saline) was added to the tissues to serve as a vehicle and time control.
We also examined the ability of opioid receptor antagonists toreverse the nociceptin effect. After acquisition of the two control responses to EFS, a single concentration of nociceptin (0.1 μM) was added to the tissue, and 15 min later the EFS response was again obtained. In all experiments, nociceptin caused a concentration-related reduction in the EFS response (compared with the time control). The antagonist (naloxone, nor-binaltorphimine or naltrindole) was then added to the bath, and 20 min later the final (fourth) EFS response was obtained. If nociceptin were acting on a classical opioid receptor, then a large concentration of a competitive antagonist would reverse its effect. As noted in “Results,” we observed no difference in the amplitude of five consecutive EFS responses in vehicle and time-control studies. In some experiments, the opioid agonist DAMGO was given in place of nociceptin.
Data analysis.
At the end of each experiment, the bronchus was contracted with barium chloride (30 mM). The contraction evoked was taken as the maximum obtainable contraction. Nociceptin had no effect (P > .05) on the contraction to barium, which averaged 2.1 ± 0.2 g. With a 15-s train of stimuli, the EFS-induced contraction does not reach a steady state (fig. 1). The peak contraction to EFS or capsaicin was expressed as a percentage of this maximum barium chloride-induced contraction. When comparing the mean responses for two treatments we analyzed the data using Student’st test for paired comparisons. When comparing multiple mean values (antagonist study), we first analyzed the data using an ANOVA calculation. The data are expressed as mean ± S.E.M., and n refers to the number of experiments carried out on tissues obtained from separate animals.
RT-PCR.
Cervical dorsal root ganglia, jugular ganglia, cervical spinal cord, liver and kidney were removed and snap-frozen in liquid nitrogen (n = 5). Total RNA was isolated using an extraction kit (Rneasy mini kit; Qiagen, Santa Clarita, CA). In a separate control experiment, no RNA was included in the reaction. First strand cDNA was synthesized by incubation of 1.5 μl RNA (∼1.5 μg) with 1 μl of oligo dTs (Pharmacia Biotech, Freiburg, Germany). The oligo dTs (200 μg/ml) were incubated at 70°C for 10 min, followed by 3 min in an ice bath and subsequent addition of 4 μl RT buffer (5×), 1 μl reverse transcriptase (Gibco BRL, Eggenstein, Germany), 1 μl dNTPs (Invitrogen BV, NV Leck, Netherlands) and 1 μl RNA guard (Pharmacia), and were dissolved to a final volume of 20 μl by addition of 7 μl of autoclaved water. This was then incubated for 90 min at 37°C. The following primers were chosen from the cloned sequence of guinea pig ORL (GeneBank accession number U04369, NID g606788): 5′-TCAGCCTGCTAAGTCTCTGTGCTTTCGGTG, 3′-CTCTGGAGTATGGAACCTTTACCGTCCTGGA, amplification product 747 bp. Thirty cycles (94°C for 30 s, 67.3°C for 1 min and 72°C for 30 s) of PCR were performed using a thermocycler (Perkin-Elmer, Norwalk, CT; model 2400). The PCR products obtained were separated by a 1% (w/v) agarose gel and visualized with the aid of ethidium bromide.
Double-labeling immunohistochemistry.
Immunofluorescence was performed as described before (Fischer et al., 1996). Briefly, guinea pig bronchi were fixed in Zamboni’s fixative (2% paraformaldehyde/15% saturated picric acid in phosphate buffer, pH 7.4). Cryostat sections of each tissue were used for single-labeling (n = 5) or double-labeling (n = 3) immunohistochemistry. The cryostat sections of Zamboni-fixed bronchi and lungs, after blocking of nonspecific binding sites, were incubated with an anti-nociceptin antiserum from rabbit (BioTrend, Cologne, Germany; dilution 1:1500) alone or in combination with a monoclonal anti-substance P antibody from rat (Serva, Heidelberg, Germany, dilution 1:200) overnight at room temperature. For the subsequent detection of the monoclonal antibody, an anti-rat biotinylated immunoglobulin (Ig) from sheep (Amersham, Braunschweig, Germany; dilution 1:50) was applied. This was followed by a mixture of a streptavidin-Texas red conjugate (Amersham; dilution 1:50) and fluorescein isothiocyanate (FITC)-conjugated anti-rabbit immunoglobulin from goat (Organon-Teknika, Eppelheim, Germany; dilution 1:500) for detection of the rabbit antiserum. Sections were viewed with an Olympus epifluorescence microscope (BX 60F; Olympus, Hamburg, Germany). Controls were performed by preincubation of the primary antiserum with synthetic nociceptin, dynorphin A, leu-enkephalin or met-enkephalin (20 μg/ml diluted antiserum; overnight, +4°C). The preabsorbed antiserum was then applied as described above. Labeling was regarded as specific when it could be prevented by preabsorption with nociceptin but not with enkephalins or dynorphin A.
Results
Functional studies.
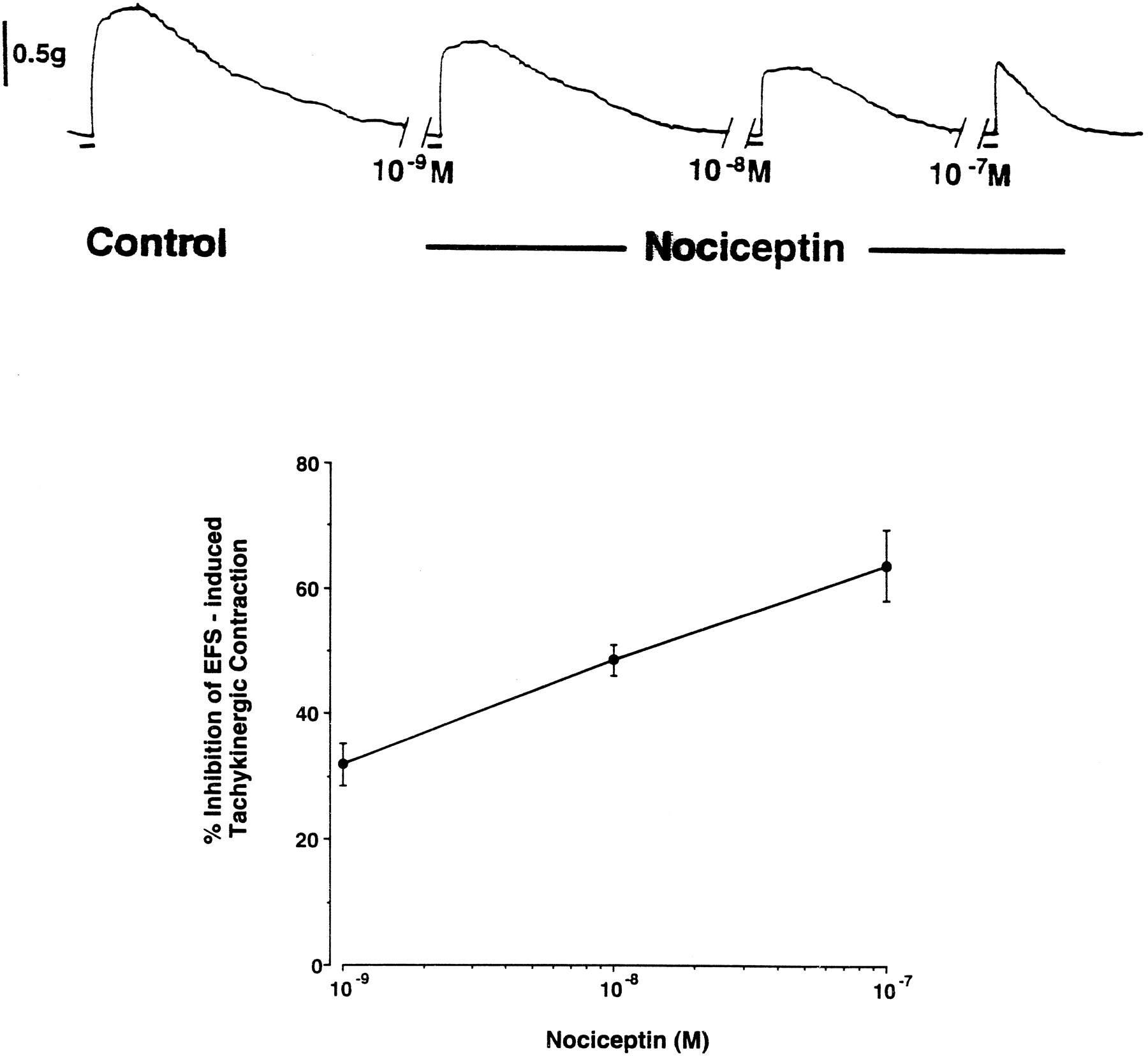
EFS with 1-ms pulses (12 V, 5 Hz, for 15 s) resulted in a contraction of the propranolol- and atropine-treated tissues that averaged 15.9 ± 2.1% of the maximum contraction (n = 11). The noncholinergic, nonadrenergic contractions of the guinea pig bronchus in response to EFS have been previously noted to be tachykinergic in nature (see Ellis and Undem, 1994a). It is consistent with this that in three of our experiments, the EFS-induced contractions were virtually abolished by a combination of CP 9994 (1 μM) and SR 48968 (1 μM). These compounds are selective antagonists of neurokinin NK-1 and NK-2 receptors, respectively (McLean et al., 1993; Edmonds-Alt et al., 1992). Nociceptin (1 nM–100 nM) effectively inhibited the EFS-induced tachykinergic contractions in a concentration-dependent fashion, beginning with 1 nM (fig. 1). Concentrations greater than 100 nM were not studied. At 100 nM the percentage inhibition averaged 56.2 ± 6.7% (n = 11). In a result consistent with our previous study (Undem et al., 1990), a time-control study revealed no differences between the control response and the subsequent responses obtained in the presence of only vehicle at 20-min intervals; in three experiments, the control response was 25.1 ± 3.2% of the maximum contraction, and the response to the fifth subsequent EFS averaged 23.1 ± 2.0%.
The inhibitory effect of nociceptin on the EFS-induced tachykinergic contractions was not reversed by the nonselective opioid receptor antagonist naloxone (10 μM) (table 1). In agreement with other reports (Belvisi et al., 1990), however, we found that the opioid-receptor agonist DAMGO (1 μM) inhibited the EFS-induced contractions by 62.7 ± 9.7%, and this effect was reversed by 48.7 ± 18% with 10 μM naloxone (n = 4, P < .05; data not shown). This suggests that nociceptin does not act via a classical opioid receptor to inhibit the tachykinergic contractions. Supporting this hypothesis, neither nor-binaltorphimine, a potent OP2 (κ) receptor antagonist, or naltrindole, a potent OP1 (δ) receptor antagonist (Dhawan et al., 1996), reversed the nociceptin effect (table 1). Note that the inhibition of the EFS response by the single application of 0.1 μM nociceptin in these experiments was not different from that obtained in the concentration-response study (fig. 1), which indicates that in the latter study, the previous addition of 1 nM and 10 nM nociceptin to the bronchus did not lead to significant desensitization.
Capsaicin (10 nM–1 μM) caused a concentration-dependent contraction of the bronchus. As with the response to EFS, and as is consistent with our previous findings (Ellis and Undem, 1994b), these contractions were abolished with the combination of CP 9994 (1 μM) and SR 48968 (1 μM), a result that indicates their tachykinergic nature (data not shown). By contrast to its effect on the electrically evoked tachykinergic contractions, however, nociceptin (0.1 μM) had no significant effect on the contractions elicited by capsaicin (fig. 2).
RT-PCR.
In tissues derived from each of five animals, a PCR product with a length of 747 bp was amplified from mRNA from jugular and dorsal root ganglia using guinea pig ORL1-specific primers (fig. 3). No product was amplified from mRNA obtained from kidney or liver tissue. This amplification product corresponds to the length of the sequence flanked by the primer pair. PCR products with the same length were amplified from RNA extracted from the spinal cord (fig. 3).
Double-labeling immunohistochemistry.
Although a detailed morphometric analysis was not performed, we noted specific nociceptin-positive fibers in guinea pig bronchi. Nociceptin-positive fibers were seen in bronchi of each of eight animals studied (fig. 4); however, not every section contained nociceptin-positive fibers. The specificity of the nociceptin-immunoreactivity was demonstrated by inhibition of the labeling when antinociceptin antiserum was preabsorbed with nociceptin (fig. 4D). Labeling was unaffected when the antiserum was preincubated with dynorphin A (fig. 4C), met-enkephalin or leu-enkephalin (data not shown). In bronchi obtained from three animals, we used double-labeling techniques to determine whether the nociceptin and substance P were colocalized to the same fiber population. In each section studied, the nociceptin-immunoreactive fibers were distinct from the substance P-immunoreactive fibers (fig. 4, A and B).
Discussion
These data demonstrate that nociceptin is a potent (threshold about 1 nM) and effective inhibitor of smooth muscle contractions evoked by electrical stimulation of tachykinin-containing nerve fibers in the guinea pig bronchus. This observation is consistent with the findings of Giuliani and Maggi (1996), who recently showed that nociceptin inhibits the tachykinergic responses in the guinea pig bladder evoked by afferent nerve stimulation.
Nociceptin is a heptadecapeptide with homology to dynorphin A. By contrast to dynorphin A, however, nociceptin has only very low affinity for opioid receptors. Our observations that neither the nonselective opioid antagonist naloxone nor potent and selective antagonists for the OP1 (δ) or OP2 (κ) receptors inhibited the effect of nociceptin support the hypothesis that the inhibitory effect of nociceptin on the tachykinergic contractions occurs independently of classically defined opioid receptors. It would seem more likely that nociceptin actsvia the ORL receptor. The ORL1 receptor has 50–60% homology with other opioid receptors but has low or no affinity for opioid ligands. Nociceptin, by contrast, binds to the ORL1 receptor with high affinity (Mollereau et al., 1994). The tachykinin-containing C-fiber afferents in the guinea pig large airways arise primarily from cell bodies in the jugular ganglion (Kummeret al., 1992; Riccio et al., 1996). Considering that nanomolar concentrations of nociceptin were capable of inhibiting tachykinergic responses, along with the evidence that cells in the jugular ganglion contain the ORL receptor-mRNA, provides circumstantial evidence that a nociceptin-ORL receptor interaction leads to inhibition of tachykinergic transmission in the airway.
The decrease in electrically evoked tachykinergic contractions of the bronchus could occur either by inhibiting tachykinin release (prejunctional mechanism) or by inhibiting the smooth muscle response to the released tachykinin (postjunctional mechanism). The fact that nociceptin had no effect on the contractions elicited by capsaicin supports the hypothesis that a prejunctional mechanism is involved. As with electrical stimulation, capsaicin causes contractions of the bronchus by releasing tachykinins from afferent nerves. Thus, pharmacologically antagonizing NK-1 and NK-2 receptors blocks the contractions to both EFS (in presence of atropine) and capsaicin (present study, Ellis and Undem, 1994a,b). By contrast to electrical stimulation, however, the response to capsaicin can occur independently of tetrodotoxin-sensitive sodium channels (Canning and Undem, 1994). If nociceptin inhibited the electrically evoked tachykinergic contractions by a postjunctional mechanism, then it should have the same effect on the response whether the tachykinins were released electrically with EFS or chemically with capsaicin. That nociceptin selectively inhibited the electrically evoked response indicates that it probably acts prejunctionally to inhibit selectively the action potential-driven release of tachykinins. The anatomical evidence that ORL receptors-mRNA is expressed in cells of in the jugular ganglion neurons also circumstantially supports a prejunctional mechanism of action.
The precise mechanism by which nociceptin leads to an inhibition of tachykinin release is not known, but the selective effect on EFSvs. capsaicin-induced responses suggests a voltage-dependent mechanism. Nociceptin amplifies an inward-rectifying potassium current in neurons of the locus ceruleus and raphe nucleus (Vaughan and Christie, 1996; Connor et al., 1996a), inhibits voltage-dependent calcium channels in neuroblastoma cells (Connoret al., 1996b) and inhibits forskolin-induced cAMP accumulation in cells transfected with ORL1 (Reinscheid et al., 1995). Each of these mechanisms could conceivably contribute to an inhibition of electrically evoked transmitter release in the bronchus. Morphine and other opioid receptor ligands have been shown to inhibit electrically evoked tachykinergic contractions of the guinea pig bronchus by a mechanism that is blocked by the opioid receptor antagonist naloxone (Bartho et al., 1987; Belvisi et al., 1990; Johansson et al., 1989; Martran et al., 1989). The inhibition of tachykinergic transmission in the airways by opioid receptor agonists can be blocked by the potassium channel blocker charybdotoxin (Stretton et al., 1992), which supports the hypothesis that the opioids inhibit tachykinin release by enhancing the activity of a charybdotoxin-sensitive potassium channel. We attempted to address this hypothesis as it relates to the mechanism of action of nociceptin, but unfortunately, in our hands charybdotoxin (10 nM) and iberiotoxin (10 nM) appeared to affect the bronchial smooth muscle directly, leading to contractions ranging from 19% to 66% of maximum (n = 4) and, in the case of iberiotoxin, rendered the EFS-induced contractions insensitive to tetrodotoxin (n = 2).
Using immunohistochemistry we localized endogenous nociceptin to nerve fibers within the bronchus. Whereas nerve fibers that are immunoreactive for dynorphin or met-enkephalin are frequently also immunoreactive for substance P (Kummer et al., 1992), this coexistence was not seen for nociceptin. These anatomical observations, coupled with the functional data, support the speculation that nociceptin may play a physiological role in regulating tachykinergic transmission in the airways.
Acknowledgments
We thank Sonya Meeker and Silke Wiegand for their expert technical assistance.
Footnotes
-
Send reprint requests to: Axel Fischer, Institut für Anatomie und Zellbiologie, Justus-Liebig-Universität, Aulweg 123, 35385 Giessen, Germany.
-
↵1 Part of this work has been previously presented in abstract form: Undem BJ, Meeker SA, and Fischer A (1997) Am. J. Respir. Crit. Care Med. 155:A484.
-
↵2 This study was funded in part by a joint program of the National Institutes of Health, Bethesda, Maryland, and the Bundesministerium fur Bildung und Forschung, Bonn, Germany.
- Abbreviations:
- ORL1
- opioid receptor-like
- EFS
- electrical field stimulation
- NK
- neurokinin
- PBS
- phosphate-buffered saline, PNS, peripheral nervous system, TTX, tetrodotoxin, RT-PCR, reverse transcription polymerase chain reaction
- dNTP
- desoxynucleotide triphosphate
- Received September 15, 1997.
- Accepted December 22, 1997.
- The American Society for Pharmacology and Experimental Therapeutics
References

Top) Photocopy of a polygraph tracing showing the inhibitory effect of nociceptin on EFS-induced tachykinergic contractions of the guinea pig isolated bronchus. The contractions were evoked by EFS (20 V, 1 ms, 5 Hz, 15-s train) given at 20-min intervals. The control response shown is the second of two control responses. Nociceptin, at the concentrations indicated, was added to the tissue bath 15 min before the EFS. The horizontal bars denote 15 s (the duration of the EFS). Bottom) Concentration-response curve of nociceptin-induced inhibition of tachykinergic contractions of the guinea pig isolated bronchus evoked by EFS. The response to a given concentration of nociceptin was compared to the control response and expressed as a percentage inhibition. As noted in “Results,” there was no difference among five consecutive EFS-induced contractions when given at 20-min intervals (data not shown). Each point represent the mean ± S.E.M. of six experiments. Each of the three concentrations of nociceptin caused a significant (P < .05) inhibition of the EFS-induced contraction.

Concentration-response curves for capsaicin-induced contractions of guinea pig isolated bronchus in the absence (•) and presence (○) of nociceptin (0.1 μM). The data are expressed as a percentage of the maximum contraction elicited by barium chloride added at the end of the experiment and are presented as the mean ± S.E.M. of five experiments.

RT-PCR for ORL1-mRNA. Amplification products and markers (M) were separated on an agarose gel (1%) and stained with ethidium bromide (lane 1). RT-PCR for ORL1-mRNA in guinea pig spinal cord (lane 2), jugular ganglion (lane 3) and dorsal root ganglion (lane 4). No PCR product was amplified when RNA was replaced by distilled water (lane 5) or by RNA extracted from kidney (lane 6) or liver (lane 7). These data are representative of five experiments.

Immunohistochemical localization of endogenous nociceptin in nerve fibers in the guinea pig bronchus and correlation with substance P by double-labeling immunohistochemistry. a) A nerve fiber immunoreactive for nociceptin (arrows). b) Nerve fibers immunoreactive for substance P (arrowheads). Note that in this double-labeling experiment, the fiber that is immunoreactive for nociceptin is not immunoreactive for substance P, and vice versa. c) The labeling of nociceptin-immunoreactive nerve fibers (arrows) is not affected when the nociceptin antiserum is preincubated with dynorphin A. d) The nociceptin labeling is blocked when the antiserum is preabsorbed with nociceptin. Bar represents 20 μm.
Inhibition of EFS-induced tachykinergic contractions by nociceptin in the absence and presence of opioid receptor antagonists1-a




{kind=link}
{kind=link}
{kind=link}
{kind=link}