





Abstract
Several cellular and molecular alterations have been described in skeletal and respiratory muscles of patients with chronic obstructive pulmonary disease (COPD), but information on potential abnormalities of mitochondrial function is scarce. The aim of the present study was to investigate mitochondrial function in the vastus lateralis (VL) and external intercostalis (EI) of COPD patients.
Biopsies from VL and EI were obtained during surgery for lung cancer in 13 patients with mild to moderate COPD (age 68±6 yrs, forced expiratory volume in one second (FEV1) 66±15% predicted) and 19 control subjects (age 67±9 yrs, FEV1 95±18% pred). State 3 and 4 mitochondrial oxygen consumption (V′O2,m), ATP synthesis, citrate synthase, cytochrome oxidase (COX) and complex I–III activities, as well as reactive oxygen species (ROS) production, were determined.
In COPD patients, in both muscles, COX activity (VL: COPD 3.0±0.8 versus control 2.0±0.8; EI: 3.7±1.6 versus 2.4±0.9 μmol·min−1·mg−1) and ROS production (VL: 1,643±290 versus 1,285±468; EI: 1,033±210 versus 848±288 arbitrary units) were increased, whereas state 3 V′O2,m was reduced (VL: 2.9±0.3 versus 3.6±0.4; EI: 3.6±0.3 versus 4.1±0.4 mmol·min−1·kg−1).
Skeletal muscle mitochondria of patients with chronic obstructive pulmonary disease show electron transport chain blockade and excessive production of reactive oxygen species. The concurrent involvement of both vastus lateralis and external intercostalis suggests a systemic (rather than a local) mechanism(s) already occurring in relatively early stages (Global Initiative for Chronic Obstructive Lung Disease stage II) of the disease.
- Chronic obstructive pulmonary disease pathophysiology
- muscle disorders
- oxidative stress
- quadriceps muscle
- reactive oxygen species
- respiratory muscles
Chronic obstructive pulmonary disease (COPD) is frequently associated with several extra-pulmonary manifestations that significantly influence the course and prognosis of the disease 1. Skeletal muscle dysfunction (SMD), one of the first systemic effects of COPD identified, contributes to limit exercise capacity and to jeopardise health status in these patients 2. Furthermore, it heralds poor prognosis, independently of the degree of airflow limitation 3. Thus, a better understanding of the mechanisms leading to SMD in COPD is of great clinical interest.
Previous studies have described a number of cellular and molecular abnormalities in the skeletal muscle of patients with COPD that can contribute to SMD. These include, among others, a shift in the fibre type distribution, the presence of fibre atrophy, evidence of oxidative and nitrosative stress, protein nitrosylation and enhanced apoptosis 2, 4–6. Despite the fact that mitochondria are involved in energy production and oxidative metabolism, as well as in the control of apoptosis, direct measurements of mitochondrial function in COPD muscles have seldom been obtained. Sauleda et al. 7 were the first to report that the activity of cytochrome oxidase (COX), the terminal enzyme of the mitochondrial electron transport chain, was upregulated in skeletal muscle of COPD patients in direct proportion to arterial hypoxaemia. More recently, Gosker et al. 8 found that the content of uncoupling protein 3 (a protein channel that facilitates proton leak, thus uncoupling oxidative phosphorylation from ATP synthesis, with energy dissipated as heat) was decreased in skeletal muscle of patients with COPD. Finally Ribera et al. 9 reported increased mitochondrial electron transport in saponine-skinned isolated fibres of respiratory muscles of severe emphysematous patients. Collectively, these observations suggest that mitochondrial function in skeletal muscle of COPD patients may be altered.
The present study sought to extend these previous and partial observations by comprehensively characterising mitochondrial function in skeletal muscles of patients with COPD. To this end, state 3 and 4 respiration, ATP production, the activities of COX, respiratory chain complex I+III and citrate synthase (CS) and the production of reactive oxygen species (ROS) were determined in mitochondria isolated from the vastus lateralis (VL) of patients with moderate COPD and control subjects with normal lung function, matched for age and degree of regular physical activity. Furthermore, because respiratory muscles seem to behave differently from skeletal muscle in patients with COPD 7, mitochondria isolated from the external intercostalis (EI) were also studied in these same patients.
METHODS
Study design
Biopsies from VL and EI were obtained during thoracic surgery for lung cancer (stage I or II) in 13 patients with moderate COPD 1 and 19 patients with normal lung function and no significant comorbidities or locomotor dysfunction. None of them had received chemotherapy before biopsies were obtained. All participants signed an informed consent after being fully aware of the objectives and nature of the study. The Institutional Committee for Ethics in Human Research of the Hospital General Universitario Gregorio Marañón (Madrid, Spain) had approved the study. All the aspects of the present study comply with the declaration of Helsinki.
Lung function and physical activity
Spirometry was performed using a Neumoscreen II (Jaeger, Hochberg, Germany) spirometer following international guidelines 10. Regular physical activity was evaluated by the International Physical Activity Questionnaire 11. To further characterise the exercise capacity of participants, all of them performed an incremental exercise test on an electromagnetically braked cycle-ergometer (ER-900; Jaeger) using a ramp protocol at 20 W·min−1 to a symptom-limited maximum. Ventilation and pulmonary gas exchange were measured breath-by-breath by a Quark b2 cardiopulmonary exercise system (Cosmed, Rome, Italy).
Investigations on isolated mitochondria
Surgical biopsies of VL and EI were obtained while the patient was anaesthetised, by performing a small incision about 25 cm proximal to the patella (VL) and from the incised muscles during thorax opening at the level of the 5th or 6th intercostal spaces (EI). Biopsies were transferred within 1 min to ice-cold isolation buffer (300 mM manitol, 1 mM EGTA, 10 mM Trizma-HCl (Sigma-Aldrich Co., St Louis, MO, USA), 1 mM KH2PO4, 1.74 mg·mL−1 phenylmethyl sulphonyl fluoride, 0.2% bovine serum albumin, 10 mg·L−1 amoxicillin, pH 7.4). The muscle samples were then placed on a Petri plaque surrounded by ice slurry. Mitochondria were isolated from these biopsies following previously described methodology 12. Briefly, after dissecting the fat and fibrous tissue, samples were weighed, washed with EGTA and chopped up into small pieces with a blade and suspended in isolation buffer (1:2 wt:vol). This was then homogenised with a Potter–Eveljehm homogeniser (B. Braun Medical AG, Emmenbrücke, Germany) avoiding heating of the sample. The homogenate was centrifuged at 4°C in a Sorvall RC-5, rotor SS-34, once for 10 min at 1,075×g, followed by three steps of 10 min at 8,635×g at 4°C. The final mitochondrial pellet was re-suspended in an Eppendorf tube with 150 µL of measurement buffer (300 mM Manitol, 10 mM Trizma-HCl, 1 mM KH2PO4, pH 7.4) and stored on ice.
Mitochondrial oxygen consumption (V′O2,m) was measured with a Clark-type electrode (YSI Inc., Yellow Springs, OH, USA) in a metacrilate chamber at 37°C equipped with magnetic stirring in the presence of 10 μL of succinate 5 mM with (state 3) or without (state 4) ADP 5 mM. V′O2,m was expressed in mmol·min−1·kg−1 of muscle tissue. The respiratory control index (RCI) was calculated as the ratio of state 3 to state 4 rates of respiration. ATP synthesis was indirectly assessed by measuring spectrophotometrically (Uvikon 930 spectrofluorometer; Kontron, Milan, Italy) the increase in reduced nicotinamide adenine dinucleotide phosphate (NADPH) concentration coupled to glucose phosphorylation by hexoquinase and later oxidation of glucose-6-phosphate to 6-phosphogluconate by glucose-6-phosphate dehydrogenase 13. To account for potential differences in mitochondrial density, measured values of state 3 and 4 V′O2,m and rate of ATP synthesis were expressed as tissue activities 14 in mmol·min−1·kg−1 of muscle, i.e. taking into account the dilution factor, the yield of mitochondria (an indirect validated marker of mitochondrial volume density in skeletal muscle) and the biopsy mass. The activities of COX (EC 1.9.3.1, complex IV), CS (EC 2.3.3.1) and complex I+III (rotenone-sensitive NADH cytochrome-c reductase; EC 1.6.2.1) were determined according to previously described spectrophotometric methods 12, 15. Aliquots of the homogenate and final mitochondrial suspension were assayed for total protein content 16. Mitochondrial yield was estimated as CS in the mitochondrial suspension relative to that of the actual biopsy. All reagents were obtained from Sigma-Aldrich Co.
ROS production and superoxide dismutase (SOD; EC 1.15.1.1) activity were measured in nine patients with COPD and 10 control subjects. Fixed volumes (10 μL) of resuspended mitochondria were incubated with 50 μM 5-(and-6)-chloromethyl-2′7′-dichlorodihydrofluorescein diacetate acetyl ester (CM-H2DCFDA) at 37°C for 60 min. ROS production is directly proportional to fluorescence emission (between 480 and 520 nm) measured using a multidetection microplate reader (SBL/AMINCO, Rochester, NY, USA). To assess ROS production during states 3 and 4 respiration, 70 mM of ADP and 10 mM of glutamate were added immediately before the addition of CM-H2DCFDA. The activity of the SOD was measured by the blue tetrazolium reduction method (Sigma-Aldrich Co.) 17.
Statistical analysis
Results are shown as mean±sd, unless otherwise specified. Comparisons between groups were performed by paired t-tests. Correlations between variables of interest were explored with the Pearson correlation test. A p-value <0.05 was considered significant.
RESULTS
Clinical data
Patients and control subjects were well matched in terms of age, weight, body mass index (BMI) and level of usual physical activity, which was relatively low in both groups (table 1⇓). Patients with COPD had moderate airflow obstruction, whereas spirometry was normal in control subjects (table 1⇓). Exercise tolerance was preserved in both groups, albeit a tendency to lower peak oxygen uptake (V′O2,peak) was seen in the COPD group. At peak exercise, patients with COPD had less ventilatory reserve (p = 0.02). While dyspnoea scores were not different between groups, leg fatigue scores tended to be higher in COPD patients (table 1⇓). Specifically questioned, all the patients declared having stopped smoking at the time of admission to the hospital.
Demographic and physiological characteristics of the two groups of subjects studied
Mitochondrial function
Table 2⇓ shows mitochondrial function measurements in VL and EI in the two groups of subjects studied. Biopsy weight, mitochondrial yield and mitochondrial protein content were similar in both groups and both muscles, supporting the reproducibility of the methodology used. Compared with control subjects, patients with COPD showed slightly but significantly reduced V′O2,m (state 3) values and ATP production, both in VL and EI (table 2⇓). The RCI was significantly reduced in the VL of patients with COPD, whereas in the EI, differences just failed to reach statistical significance (table 2⇓). The P:O ratio (i.e. the efficiency of ATP synthesis coupled with cell respiration) was similar (∼1.5) in both groups and both muscles (table 2⇓). Compared with control subjects, the activity of CS (a mitochondrial matrix enzyme) was also reduced in the VL of subjects with COPD (table 2⇓). In contrast, the activity of mitochondrial membrane enzymes (COX and complex I–III) was significantly higher in COPD patients than in control subjects in both muscles (table 2⇓). There were modest but significant (p<0.01) correlations between the V′O2,peak of the incremental exercise test and CS (r = 0.58) and state 3 respiration (r = 0.65). Interestingly, COX activity was negatively related to arterial oxygen tension (Pa,O2), at rest and during exercise (fig. 1⇓). A similar relationship was observed with the activities of the other respiratory chain enzymes (data not shown).

Correlation between arterial oxygen tension (Pa,O2) and cytochrome oxidase (COX) activity in a and b) vastus lateralis (VL) and c and d) external intercostalis (EI) muscles; a and c) at rest and b and d) during exercise. •: control subjects; ○: chronic obstructive pulmonary disease patients. a) r = -0.60, p<0.01; b) r = -0.66, p<0.01; c) r = -0.55, p<0.01; d) r = -0.59, p<0.01.
Mitochondrial function measurements in vastus lateralis and external intercostalis muscles
ROS production and SOD activity
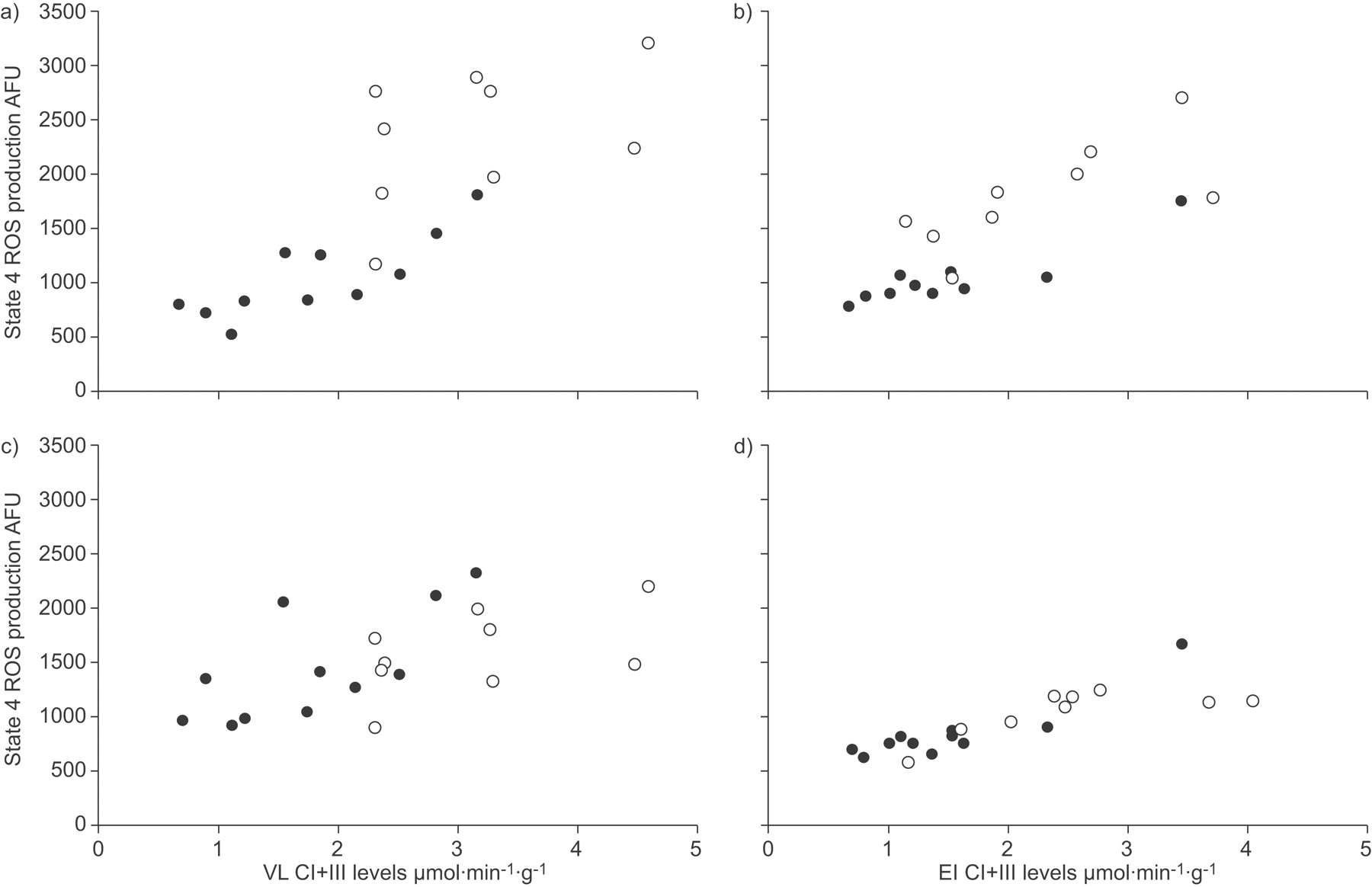
During state 3 respiration (ADP-stimulated; fig. 2a⇓) the production of ROS was significantly higher (p<0.01) in patients with COPD than in control subjects, both in VL (2.4-fold increase) and EI (1.7-fold increase). Similar results were observed during state 4 (glutamate-stimulated) respiration (VL 2.9-fold increase, EI 1.8-fold increase; p<0.05; fig. 2b⇓). Figure 2⇓ also corroborates the expectation that ROS production is highest in the absence of ADP (state 4 respiration), when the mitochondrial membrane potential is highest 18. ROS production was significantly (p<0.001) related to complex I+III activity in both state 4 and state 3, as shown in figure 3⇓, indicating, as anticipated, that the mitochondrial electron transport chain is a major source of ROS production in skeletal muscle 19.

Mean±sd reactive oxygen species (ROS) production (in arbitrary fluorescence units (AFU)) in isolated mitochondria from the vastus lateralis (VL) and external intercostalis (EI) muscles of chronic obstructive pulmonary disease patients (▓) and control subjects (□) during a) state 3 (ADP-stimulated) and b) state 4 (glutamate-stimulated) respiration. c) Mean±sd Mn2+ superoxide dismutase (SOD) activity. *: p<0.05; **: p<0.01.
{kind=link}
{kind=link}
{kind=link}
Relationship between complex (C) I+III (rotenone-sensitive reduced nicotinamide adenine dinucleotide cytochrome-c reductase) activity and reactive oxygen species (ROS) production during a and b) state 4 and c and d) state 3 respiration, in a and c) vastus lateralis (VL) and b and d) external intercostalis (EI) muscles. AFU: arbitrary fluorescents units. •: control subjects; ○: chronic obstructive pulmonary disease patients. a) r = 0.74; b) r = 0.78; c) r = 0.81; d) r = 0.84.
The activity of the antioxidant enzyme SOD was increased in mitochondria isolated from the VL in patients with COPD (p<0.05), but differences failed to reach statistical significance in EI samples (fig. 2c⇑).
DISCUSSION
The present study shows that the mitochondria isolated from the VL and EI of COPD patients present increased ROS production and a set of abnormalities consistent with mitochondrial membrane blockade 20, 21 (table 2⇑ and figs 1⇑ and 2⇑).
Previous studies
Several studies have previously investigated aspects of mitochondrial function in muscle biopsies of patients with COPD. To the current authors’ knowledge, the present study is the first describing mitochondrial function comprehensively in both peripheral and respiratory muscles in these patients. Sauleda et al. 7 reported increased COX activity in the VL of patients with COPD. The current results are in keeping with this observation (table 2⇑ and fig. 1⇑) and extend it by showing that the activities of other membrane-bound respiratory chain enzymes are also upregulated in patients with COPD (table 2⇑). Also in keeping with some former studies 22–24, the present study found decreased CS activity in the VL (table 2⇑). CS is a citric acid cycle enzyme of the mitochondrial matrix. Thus, the heterogeneous alteration of the mitochondrial oxidative metabolism enzymes found in patients with COPD suggests that the activities of the citric acid cycle and the electron transport chain are unmatched in these patients 14, 25 and points to different regulatory mechanism for both types of mitochondrial enzymes. In fact, COX and complex III are partially coded by mitochondrial DNA, while citric acid enzymes are regulated by nucleic DNA 26.
Increased ROS production by COPD striated muscle has been previously suspected from indirect findings 27; however, the present study actually measured it (figs 2⇑ and 3⇑). Superoxide anions are produced by the electron transport chain on the inner mitochondrial membrane 19 and their generation is strongly dependent upon the proton potential across the mitochondrial membrane 18. This potential was probably increased in the COPD mitochondria as a consequence of the increased respiratory enzyme activity together with the mitochondrial respiratory membrane blockade suggested by the lack of translation into more oxygen uptake 20, 21. Finally, Rabinovich et al. 28 reported reduced RCI in patients with COPD and low BMI. The present results extend these findings to the VL of subjects with normal BMI (table 1⇑).
Mechanisms
The present study is a descriptive study that does not address potential mechanisms directly. However, the observations allow some speculation. That mitochondrial dysfunction occurred in COPD both in skeletal and respiratory muscles (table 2⇑) suggests a systemic, rather than a local factor(s). Sedentarism, a frequently quoted mechanism in these patients, can be excluded, because both groups had similar activity history and both had a reasonably preserved exercise capacity (table 1⇑). Smoking may have influenced mitochondrial function 21; however, as shown in table 1⇑, there were no apparent differences in cumulative smoking exposure, carboxyhaemoglobin levels and/or percentage of current smokers between the groups. Sauleda et al. 7 reported increased COX activity in the VL of patients with COPD in proportion to arterial hypoxaemia. The current results also found an inverse relationship between respiratory chain activity and Pa,O2 (fig. 1⇑). This suggests that tissue hypoxia may upregulate respiratory chain enzymes, thus contributing to mitochondrial dysfunction. The absence of resting arterial hypoxaemia does not exclude this possibility because it may occur during exercise (fig. 1⇑) or sleep. An abnormal microcirculatory control that may eventually result in tissue hypoxia can not be excluded either 29. Finally, low-grade chronic systemic inflammation in COPD 30 may be another potential mechanism because inflammatory cytokines are related to basal energy metabolism and are known to trigger the production of ROS by striated muscle fibres 27.
Implications
Mitochondrial capacity is a well known limiting factor for exercise performance 14. The current observations of a correlation between state 3 respiration, CS activity and exercise capacity suggests that the functional abnormalities identified can contribute to limit exercise in patients with COPD. Conversely, it is worth noting that the enhanced ROS production observed in patients with COPD (fig. 2⇑) can produce further skeletal muscle damage through several non-mutually exclusive pathways. First, oxidative stress can alter the structure of several components of the respiratory chain 31 and cause proton leak and mitochondrial uncoupling 20, 32, particularly during exercise 14, 33. As a consequence, the potential for physical activity in these patients and/or their resistance to fatigue under conditions of increased respiratory load, such as episodes of exacerbation of COPD, may be impaired. Secondly, oxidative stress can be involved in muscle atrophy, a well recognised poor prognostic factor in COPD 3, since it alters the structure of muscle proteins, including myosin, facilitating their degradation by the ubiquitin-proteosome system 6, 34. It can also trigger apoptosis, an event described in the skeletal muscle of COPD patients 35. Finally, excessive ROS production deteriorates mitochondrial DNA and contributes to perpetuating muscle damage 36.
Potential limitations
Several potential limitations of the current study deserve comment. First, mitochondrial function was measured in vitro. Although this was done under strictly controlled experimental conditions following the same standard methodology in both groups 19, the results cannot be readily extrapolated to in vivo conditions. The fact that a high yield (∼35%) of functionally intact mitochondria 12 was obtained and values of RCI, P:O ratio and V′O2,m (table 2⇑) were similar to previously published results 33, 37, 38 indicates good preservation of the organelle’s membrane and supports the adequacy of the current study methodology. Moreover, previous studies using similar isolation procedures showed a good relationship between mitochondrial respiration in vitro and muscle maximum oxygen consumption in vivo 14. Secondly, using succinate, as in the present study, will provide electron input starting from complex II that typically renders lower state 3 V′O2,m (about 85% of the maximal state 3 respiration measured with other substrates) 33, 38. However, this fact will not substantially alter the current conclusions, since COPD results were compared with non-COPD control subjects studied in exactly the same way. Thirdly, smoking may have influenced the current results, but declared smoking history and carboxyhaemoglobin levels were similar between patients and control subjects. Furthermore, because smoking is forbidden in the hospital, it is likely that the majority (if not all) current smokers had not smoked during the 24 h they were hospitalised before surgery, when muscle samples were obtained. Fourthly, the subjects studied had localised lung cancer. The present approach has been used in other studies of respiratory muscles in COPD 9 and the current authors do not believe that it influenced the results because the same happened in both patients and control subjects and previous work has found no differences in the structural characteristics and expression of inflammatory cytokines and growth factors in samples from EI of patients with localised lung cancer 39. Finally, patients with mild-to-moderate COPD and normal BMI (table 1⇑) were studied, so results may be different in more advanced stages of the disease.
Conclusions
Patients with mild-to-moderate chronic obstructive pulmonary disease show evidence of mitochondrial blockade in both skeletal and respiratory muscles, suggesting that functional mitochondrial abnormalities occur in relation to systemic (as opposed to local) factors, and that they are already present at moderate stages of the disease (Global Initiative for Chronic Obstructive Lung Disease stage II).
Support statement
CIBERES is an initiative of the Instituto de Salud Carlos III (Ministerio de Ciencia e Innovación, Madrid, Spain). The present study was supported by the Fondo de Investigaciones Sanitarias (PI052563) and Neumomadrid (both in Madrid).
Statement of interest
None declared.
Acknowledgments
The authors thank the patients participating in this study for their willingness to contribute to the advancement of science. They are also grateful to A.L. Andreu and H. García-Arumí (Centre d’Investigació en Bioquímica i Biologia Molecular, University Hospital Vall d’Hebron, Barcelona, Spain) for their help in the determination of the complex I–III of the respiratory chain.
Footnotes
-
Earn CME accreditation by answering questions about this article. You will find these at the back of the printed copy of this issue or online at www.erj.ersjournals.com/current.dtl
- Received July 22, 2008.
- Accepted December 5, 2008.
- © ERS Journals Ltd
References










