





Abstract
T-helper (Th) 2 cytokines are thought to mediate most features of allergic inflammation in atopic asthma. However, it remains unclear whether chemokine pathways direct selective recruitment of Th2 cells to the airways during human allergic responses.
Bronchoalveolar lavage (BAL) was performed in 15 nonsmoking mild atopic asthmatics before and 24 h after a fibreoptic segmental allergen challenge, and chemokines related to T-cell recruitment were assayed by ELISA.
The Th2-related C-C chemokine (CCR)4 ligands, macrophage-derived chemokine/C-C chemokine ligand (CCL)22 and thymus and activation-regulated chemokine/CCL17, were increased in BAL after challenge. These chemokines correlated significantly with lymphocyte numbers and with interleukin (IL)‐5 and IL‐13 in post-challenge BAL. In contrast, two out of three putative Th1-related chemokines did not change. There were no alterations in monokine induced by interferon (IFN)‐γ/CXC chemokine ligand (CXCL)9 or macrophage inflammatory protein‐1α/CCL3; whereas a significant increase in IFN-induced protein-10kDa/CXCL10 was observed, which did not correlate with the T-cell influx. In peripheral mononuclear cells from atopic donors, CCL22 and CCL17 were induced by IL‐4 and IL‐13, further supporting the relationship between CCL22/CCL17 and Th2 cytokines. Finally, CCL22 was able to trigger actin polymerisation in peripheral CD4+ T-cells expressing CCR4.
Thus, C-C chemokine receptor 4 ligands are up-regulated in the airways of atopic asthmatics following allergen exposure, contribute to the T-cell influx to the airways and are closely related to the Th2-cytokine response.
C. Pilette is supported by a research fellowship from the European Respiratory Society, Lausanne, Switzerland (ERS; Grant No. LTRF2002-037). This work was supported by a grant from the National Asthma Campaign UK and financial support from GlaxoSmithKline (for the salary of J.N. Francis). *Authors contributed equally to this study.
Tissue infiltration by T-cells that are polarised towards a T-helper (Th) 2 phenotype has emerged as an important feature of asthma and related allergic disorders 1. Thus, T-cells producing specific cytokines (interleukin (IL)‐4, IL‐5, IL‐9 and IL‐13) are crucial for immunoglobulin (Ig)E synthesis, mucus production, the recruitment and activation of eosinophils and mast cells, and for the development of airway hyperresponsiveness (AHR) 2. Although induction of the expression of Th2 cytokines during late asthmatic responses has been documented in several studies 3–6, the profile of the chemokine response to allergen challenge has been only partly defined in atopic asthma.
It has been shown that eotaxin is rapidly induced after allergen challenge in seasonal asthmatics, and correlates with the number of airway eosinophils 7. The eotaxin receptor, C-C chemokine receptor (CCR)3, is expressed by eosinophils and basophils, but at a very low level on activated human T-cells in vivo 8, 9. In addition to CCR3, CCR4 and CCR8 have been associated with the Th2 phenotype 10. More specifically, the CCR4 pathway has been proposed to mediate the allergen-induced recruitment of T-lymphocytes to the airway in mice 11, 12 and humans 9, 13. However, the expression profile of Th2 versus Th1 chemokines, with regard to influx of T-lymphocytes to the airway, has not yet been evaluated in the same study during human late asthmatic responses. It therefore remains unclear whether the chemokine response of the airways from atopic asthmatics to allergen exposure is polarised towards a Th2 type, and to what extent this response is associated with the Th2-cytokine response.
The current authors have previously shown that gene expression of CCR4, assessed by in situ hybridisation, is up-regulated in bronchial tissue from challenged atopic asthmatics 13. Moreover, CCR4 expression correlated with the T-cell infiltration induced after allergen challenge. The aim of the present study was to evaluate, in atopic asthma, the allergen-induced changes in the bronchial expression of ligands for chemokine receptors expressed by T-cells and, more specifically, those associated with Th2 versus Th1 responses. Therefore, the current authors assessed Th2- and Th1-related chemokines in the bronchoalveolar lavage (BAL) fluid recovered before and after local endobronchial allergen challenge of atopic asthmatics. The authors also evaluated the chemokine response to allergen challenge in healthy, nonatopic control subjects. Macrophage-derived chemokine /C-C chemokine ligand (CCL)22, thymus and activation-regulated chemokine/CCL17 (CCR4 ligands) and I-309/CCL1 (CCR8 ligand) represented Th2-related chemokines, while putative Th1 chemokines consisted of macrophage inflammatory protein‐1α/CCL3 (ligand for CCR1 and CCR5), monokine induced by interferon (IFN)‐γ/CXC chemokine ligand (CXCL)9 and IFN-inducible protein‐10 kDa (IP‐10)/CXCL10 (CXC chemokine receptor (CXCR)3 ligands). Chemokine pathways related to T-cells, irrespective of their Th2/Th1 phenotype (CCR6-7, CXCR1-2-4-6, CX3CR1) 14, were not assessed. The relationship between the chemokine response and Th2 cytokines (IL‐4, IL‐5, IL‐13) was investigated in vivo. In order to further evaluate the association between the chemokine and cytokine responses, the current authors assessed, in vitro, the regulation of chemokine production by cytokines in peripheral blood mononuclear cells (PBMC) from atopic donors. Finally, the authors evaluated the responsiveness of circulating, blood T-cells to CCR4 ligands using actin polymerisation as functional readout 15, 16.
Materials and methods
Patient characterisation
Fifteen nonsmoking atopic subjects with a clinical history of house dust mite- or grass pollen-sensitive asthma, as defined by the American Thoracic Society criteria, took part in the study. All patients, aged 18–55 yrs, were required to have baseline forced expiratory volume in one second (FEV1) >80% predicted and methacholine provocative concentration causing a 20% fall in FEV1 <32 mg·mL−1 but >0.5 mg·mL−1. All subjects were also required to have >5‐mm diameter weal at 15 min after skin-prick testing with house dust mite (Dermatophagoides pteronyssinus) or timothy grass pollen (Phleum pratense) extracts (Soluprick; ALK Abello, Horsholm, Denmark) in the presence of negative diluent and positive histamine controls, and serum specific IgE antibodies (detected by radioallergosorbent test (RAST)) to the same allergen. Subjects were excluded if they had received systemic glucocorticosteroids within the past 6 months or allergen immunotherapy within the previous 5 yrs. In addition, inhaled corticosteroids were discontinued for ≥2 weeks before the allergen challenge. Patient characteristics (allergen sensitivity, late-phase response) are shown in table 1⇓. In addition, four healthy, nonatopic and nonsmoking volunteers (aged 18–55 yrs, with negative skin and RAST tests to common aeroallergens) were included as a control group. The study was performed with the approval of the Ethics Committee of the Royal Brompton and Harefield Hospitals NHS Trust and the written informed consent of all the participants.
Clinical and cellular late-phase responses to allergen challenge
Fibreoptic endobronchial allergen challenge
The procedure for bronchoscopy, BAL and segmental allergen provocation was as previously described 6. During an initial endoscopy, BAL was taken from the right upper lobe and 500 biological units (100 units first and, after 5 min, 400 units in the absence of excessive local bronchoconstriction) of the relevant allergen (D. pteronyssinus or P. pratense as respective major allergens for house dust mite and grass pollen, according to the clinical history and skin-prick tests), diluted in sterile saline, were instilled in the middle lobe. All controls were challenged with P. pratense. BAL was obtained 24 h later from the challenged middle lobe segment. FEV1 was carefully monitored for at least 8 h and 22–24 h after the allergen challenge procedure. Inhaled β2‐agonists were administered as required. The maximal fall in FEV1 >4 h after the allergen challenge was used to define the clinical late-phase response.
Processing of bronchoalveolar lavage fluid
BAL samples were filtered through sterile gauze and centrifuged. Cytospins were prepared from resuspended cells and BAL supernatants were divided into 1‐mL aliquots stored at −80°C until analysis. Total and differential cell counts were performed on methanol-fixed cytospins of BAL cells after May-Grünwald and Giemsa staining.
Peripheral blood mononuclear cell cultures
PBMC were isolated from heparinised blood of atopic donors by centrifugation over Histopaque. Mononuclear cells were washed twice in RPMI‐1640 medium and resuspended in RPMI‐1640 supplemented with 2 mM l-glutamine, 5% human serum, 100 U·mL−1 penicillin and 100 µg·mL−1 streptomycin. PBMC were then incubated at 1×106 cells·well−1 for 72 h in humidified atmosphere (37°C, 5% CO2) with IL‐2 (10 ng·mL−1) and one of the following cytokines: IL‐4, IL‐5, IL‐13 or IFN‐γ (20 ng·mL−1 for each). Phytohemagglutinin (1 µg·mL−1) was used as a positive control. The supernatants were frozen at −20°C until chemokine assessments.
Chemokine and cytokine ELISA
Chemokines and cytokines were measured in unconcentrated BAL and cell culture supernatants by ELISA, using specific pairs (capture and biotinylated-detection) of antibodies. Paired antibodies to CCL1, CCL3, CXCL9 and CXCL10 were from PeproTech (London, UK), as well as detecting antibodies to CCL17 and CCL22. Capture antibodies to CCL17 and CCL22 were from R&D Systems (Abingdon, UK) and paired antibodies to IL‐4, IL‐5 and IL‐13 from BD Pharmingen (Oxford, UK). IL‐12 was detected using paired antibodies (PeproTech) to all subunits of the cytokine (p35, p40, p70).
Nunc microplates (96 well) (Life Technologies, Paisley, UK) were coated with the appropriate capture antibody and blocked with 1% weight/volume (w/v) bovine serum albumin (BSA)-phosphate buffered saline (PBS). BAL samples in duplicate and serial dilutions of recombinant chemokines/cytokines (from Peprotech, except IL‐5, which was from BD Pharmingen) were then incubated, before the appropriate biotinylated-detection antibody. The reaction was revealed by incubation with peroxidase-conjugated streptavidine, and developed with tetramethylbenzidine and hydrogen peroxide. Optical density was recorded at 450 nm and concentrations expressed in pg·mL−1. Detection limits were 5–10 pg·mL−1 for CCL22, CCL1, CCL3, IL‐4, IL‐5, IL‐13, IL‐12 and IFN‐γ, and 2 pg·mL−1 for CCL17, CXCL9 and CXCL10. General laboratory reagents were obtained from Sigma (Poole, UK).
BAL chemokine data were corrected for protein plasma leakage 17 by measuring albumin concentration in BAL before/after challenge by a chemoluminescent immunoassay using the DPC Immunolyte analyser (EuroDPC Ltd, Llanberis, Wales, UK). For each chemokine, the ratio to albumin was calculated in BAL before/after segmental challenge.
Actin polymerisation assay
Freshly isolated PBMC were washed with staining buffer (0.1% w/v BSA and 0.09% w/v sodium azide in PBS), resuspended at 5×106 cells·mL−1 and incubated with 10 µg·mL−1 of anti-CCR4-phycoerythrin (BD Pharmingen) and anti-CD4-phycoerythrin-Cy5 (DakoCytomation, Ely, UK) for 30 min. After washing for 10 min at 37°C in RPMI, either CCL22 or CCL17 at 10 ng·mL−1 was added to the cell suspension; every 15 s, 0.5×106 cells were removed and mixed with 400 µL of 10−7 M FITC-labelled phalloidin, 0.125 mg·mL−1 l‐α‐lysophosphatidylcholine and 4% paraformaldehyde (Sigma) in PBS. Fixed cells were analysed by flow cytometry using a dual laser FACScan (Becton-Dickinson, Mountainview, CA, USA) and WinMDI software (J. Trotter, The Scripps Research Institute, La Jolla, CA, USA).
Statistical analysis
Data obtained in BAL before and after allergen challenge, presented as median (interquartile range), were compared by the nonparametric Wilcoxon's match-paired test, and correlations were assessed by the Spearman's rank correlation test. Results of the cell-culture experiments presented as mean±sd were analysed by the paired t-test, and Bonferroni's correction was applied when multiple comparisons were performed with a same control condition.
Results
Lymphocyte recruitment to the airway after allergen challenge of atopic asthmatics
Endobronchial exposure to allergen was associated with highly significant elevations of the total number of cells, and of the number of lymphocytes (3.28 (0.08–11.43)×106, median (range), versus 0.36 (0.06–1.12)×106, p<0.001), recovered 24 h later by BAL (table 1⇑). Among the 10 asthmatics who developed a clinically significant late-phase response after challenge (as defined by ≥20% fall in FEV1), seven patients displayed a ≥10-fold increase in BAL lymphocytes. In contrast, none of the five nonresponders showed such an increase in the number of BAL lymphocytes. No significant increase in BAL lymphocytes was observed after challenge in nonatopic controls (not shown), along with the absence of a significant clinical late-phase response in these healthy subjects.
CCL22 and CCL17 are induced in bronchoalveolar lavage fluid after allergen challenge
CCL22, which was consistently detected in BAL fluid supernatants from atopic asthmatics at baseline (fig. 1⇓), was strongly increased in BAL 24 h after allergen challenge (p<0.001). CCL17, the other ligand for CCR4, was also consistently induced after challenge (p<0.001). In contrast, CCL1 did not change in BAL after local exposure to the allergen.
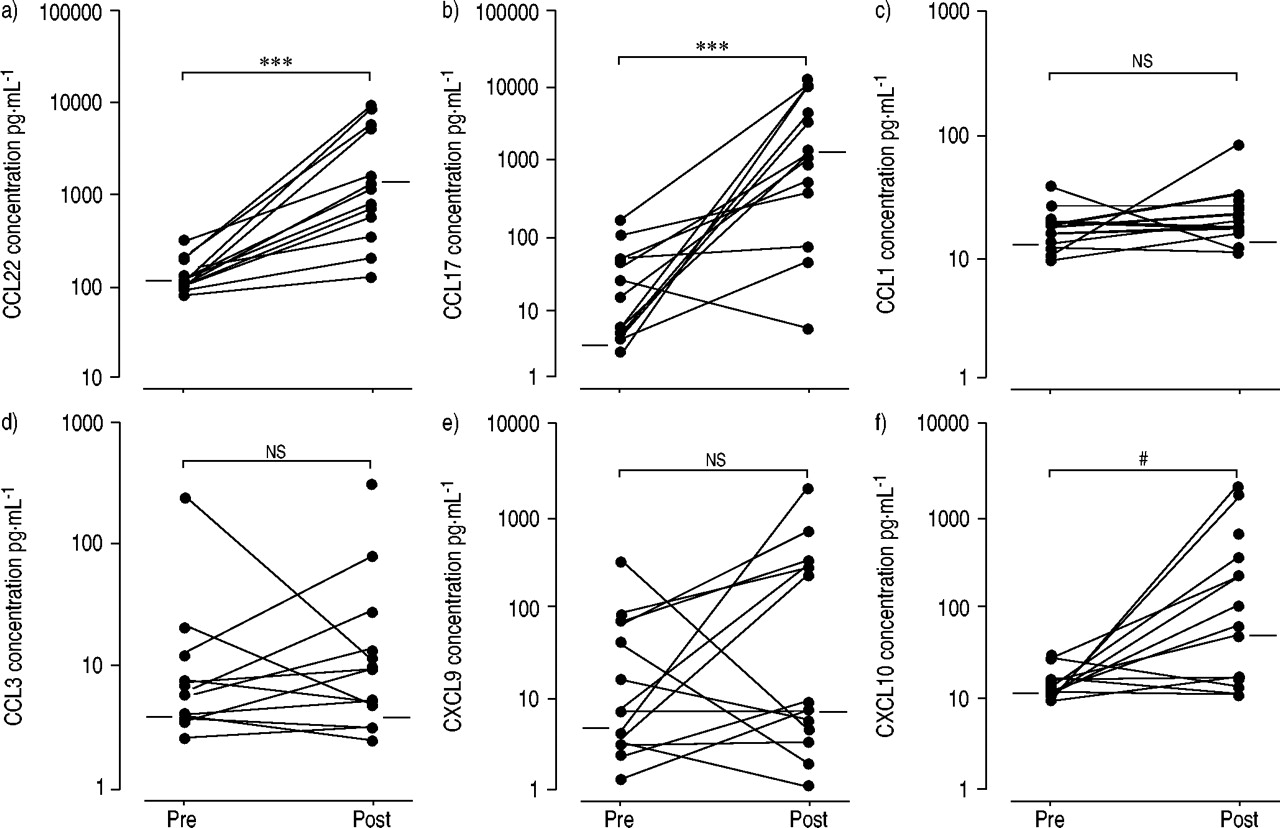
Chemokines in bronchoalveolar lavage (BAL) from atopic asthmatics (n=15) during the late response to allergen challenge. T-helper (Th)2- (a) C-C chemokine ligand (CCL)22, b) CCL17, c) CCL1) and Th1-related chemokines (d) CCL3, e) CXC chemokine ligand (CXCL)9, f) CXCL10) were assessed by ELISA in BAL recovered before (pre) and 24 h after (post) segmental challenge with the relevant allergen. Paired data were compared using the Wilcoxon's match-paired test. ***: p<0.001; ns: nonsignificant; #: p=0.004.
Among the Th1-related chemokines CCL3, CXCL9 and CXCL10, only CXCL10 was significantly up-regulated in the lung from challenged asthmatics (p=0.004).
CCL22 and CCL17 are locally produced after airway allergen challenge
To confirm that chemokines induced in BAL are locally produced within the challenged airway, the current authors corrected the concentrations of chemokines for that of albumin or urea. BAL concentration of albumin was significantly increased after allergen challenge (0.1 (0.06–0.15) versus 0.04 (0.03–0.06) mg·mL−1, p<0.001), relating to nonspecific leakage of plasma proteins into the bronchoalveolar space. BAL urea also tended to increase after challenge, but without reaching statistical significance (0.4 (0.4–0.5) versus 0.3 (0.2–0.3), p=0.07). The CCL22/albumin, CCL17/albumin and CXCL10/albumin ratios were also significantly increased after challenge, suggesting a local production within the airways, as supported by the absence of significant changes in the serum levels of these chemokines (data not shown).
CCL22 and CCL17 correlate with the allergen-induced lymphocyte influx to the airway
The level of CCL22 significantly correlated with the number of lymphocytes recovered in BAL after endobronchial allergen challenge (fig. 2⇓). A similar correlation was observed for CCL17 (in responders: r=0.62, p=0.02). Additionally, in the 10 responders, CCL22 correlated with the lymphocyte influx to the airway in terms of fold increase. No significant correlation was observed between CCL22 or CCL17 and other cell types such as eosinophils (not shown), or between CXCL10 and the allergen-induced cellular influx to the airway. However, CXCL10 correlated with the clinical late-phase response (r=0.55, p=0.04), especially in responders (r=0.65, p=0.03).

Correlation between C-C chemokine ligand (CCL)22 and lymphocytes during the late asthmatic response. CCL22 was plotted against the number of lymphocytes in a) post-challenge bronchoalveolar lavage (n=15, r=0.53, p=0.02) and b) the fold increase in lymphocytes (ratio of post-challenge/pre-challenge lymphocytes, table 1⇑) in asthmatics exhibiting a significant clinical late-phase response (n=10, r=0.71, p=0.01). Statistical significance was assessed using the Spearman's rank test.
Th2 cytokines are up-regulated after allergen challenge
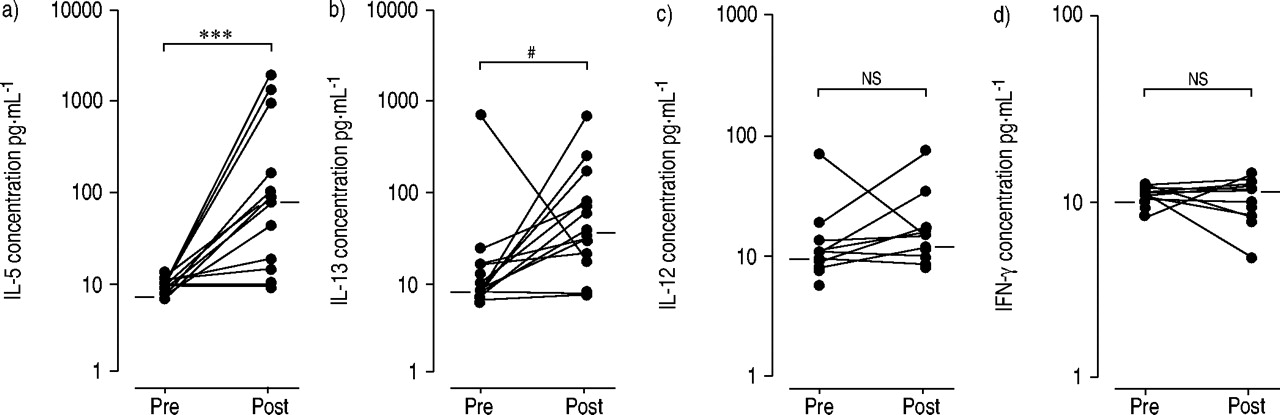
In order to assess the relationship between the chemokine and cytokine responses after bronchial segmental allergen challenge, Th2 and Th1 cytokines were also measured. IL‐5 and IL‐13 were markedly increased in BAL from atopic asthmatics 24 h after allergen challenge, as compared to baseline (fig. 3⇓; p<0.001 and p=0.002, respectively). IL‐4 remained just above the detection limit and did not change after challenge (data not shown). In contrast to Th2 cytokines, the levels of IFN‐γ and IL‐12 remained unchanged after challenge.

Cytokines in bronchoalveolar lavage (BAL) from atopic asthmatics during the late-phase response. T-helper (Th)2 (a) interleukin (IL)‐5, b) IL‐13) and Th1 cytokines (c) IL‐12, d) interferon (IFN)‐γ) were assessed by ELISA in BAL before (pre) and after (post) allergen challenge, and results were compared using the Wilcoxon's match-paired test. ***: p<0.001; #: p=0.002; ns: nonsignificant.
CCL22 and CCL17 levels correlate with Th2 cytokines
In post-challenge BAL, the level of CCL22 strongly correlated with CCL17 (fig. 4⇓; r=0.89, p<0.001). CCL22 also strongly correlated with IL‐5 and IL‐13 (r=0.84 and 0.77, respectively, p<0.001). CCL17 also correlated with the Th2 cytokines IL‐5 and IL‐13.

Relationship between T-helper (Th)2-related chemokines and Th2 cytokines during the late asthmatic response. a) C-C chemokine ligand (CCL)22 was plotted against CCL17 (r=0.89, p<0.001), and b) interleukin (IL)‐5 against IL‐13 (r=0.91, p<0.001), in bronchoalveolar lavage from challenged atopic asthmatics. c) IL‐5 (r=0.84, p<0.001) and d) IL‐13 (r=0.77, p<0.001) were also plotted against CCL22. Correlations were assessed using the Spearman's rank test.
CCL22 and CCL17 are not increased after allergen challenge in healthy controls
In healthy, nonatopic controls no significant increase was observed in CCL22 or CCL17 after segmental allergen challenge (table 2⇓). CXCL10 remained below the detection limit including after challenge. In fact, none of the measured chemokines was significantly increased in post-challenge BAL of the nonatopic, healthy controls.
Bronchoalveolar lavage (BAL) chemokines before/after allergen challenge in control subjects
CCL22 and CCL17 are up-regulated in vitro by Th2 cytokines
The current authors hypothesised that the correlation between CCL22/CCL17 and Th2 cytokines could be due to the up-regulation of their production by mononuclear cells, upon activation by Th2 cytokines. To address this hypothesis, the production of chemokines by PBMC cultured in the presence of Th2 cytokines was assessed, as compared to IFN‐γ. Production of CCL22 and CCL17 by cultured mononuclear cells was consistently induced in the presence of IL‐4 or IL‐13 (fig. 5⇓), and IFN‐γ down-regulated the constitutive production of CCL22. In contrast, CCL1 did not display this Th2-type pattern of regulation.
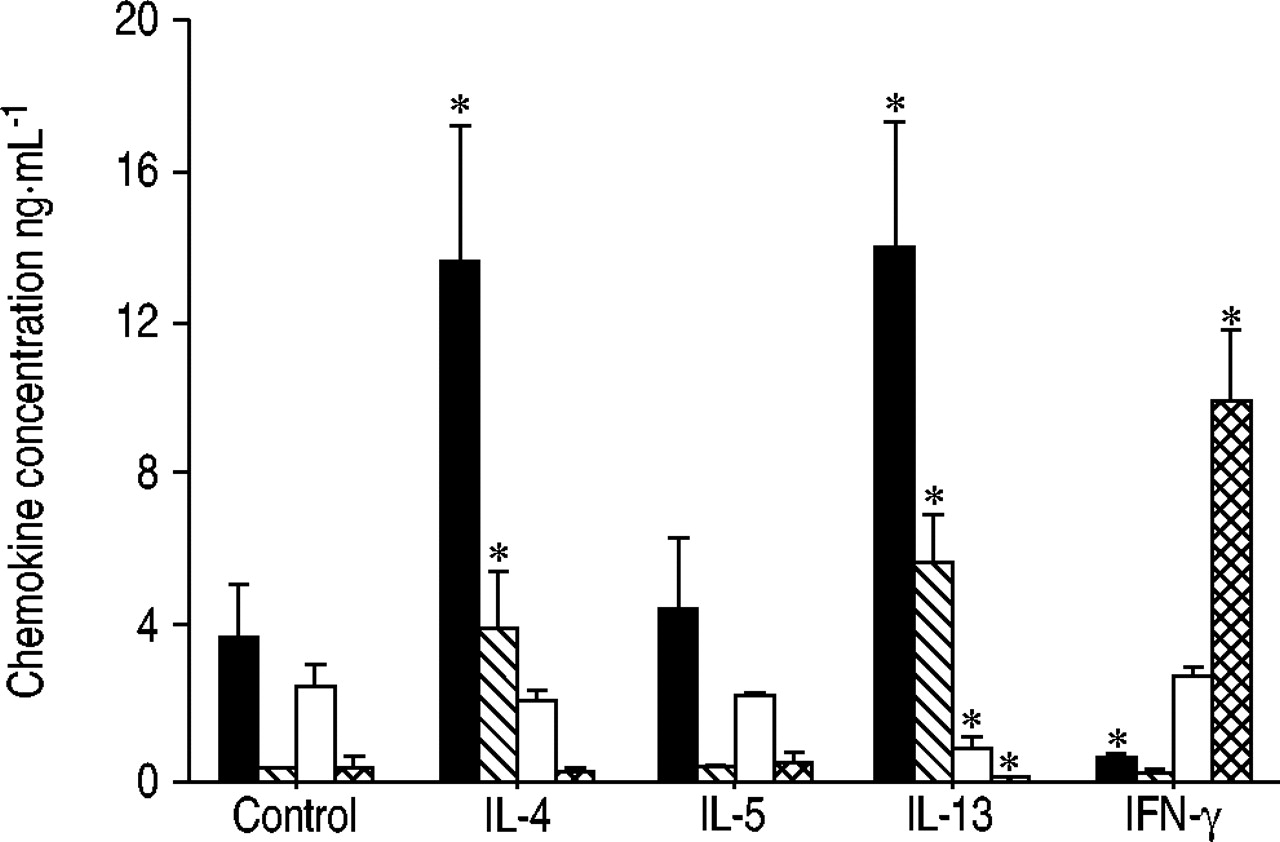
Regulation of chemokine production by T-helper (Th)2 cytokines and interferon (IFN)‐γ in peripheral blood mononuclear cells (PBMC) from atopic donors (n=3). Th2- (C-C chemokine ligand (CCL)22, CCL17 and CCL1) and Th1-related chemokines (CXC chemokine ligand (CXCL)10) were measured by ELISA in supernatants from PBMC cultured for 72 h in the presence of interleukin (IL)‐4, IL‐5, IL‐13 or IFN‐γ (each at 20 ng·mL−1). Results (mean±sd) were compared with cells incubated with medium using the paired t-test and are representative of three independent experiments. Values for CCL1 are expressed 10-fold. ▪: CCL22; ┘: CCL17; □: CCL1;  : CXCL10. *: p<0.05, as compared with control.
: CXCL10. *: p<0.05, as compared with control.
As expected, production of CXCL10 by PBMC was induced by IFN‐γ (fig. 5⇑), and was down-regulated by IL‐13 (and to a lesser extent by IL‐4). CXCL9 exhibited the same pattern of regulation as CXCL10, whereas CCL3 was not significantly increased by IFN‐γ .
CCL22 and CCL17 induce actin polymerisation in T-lymphocytes
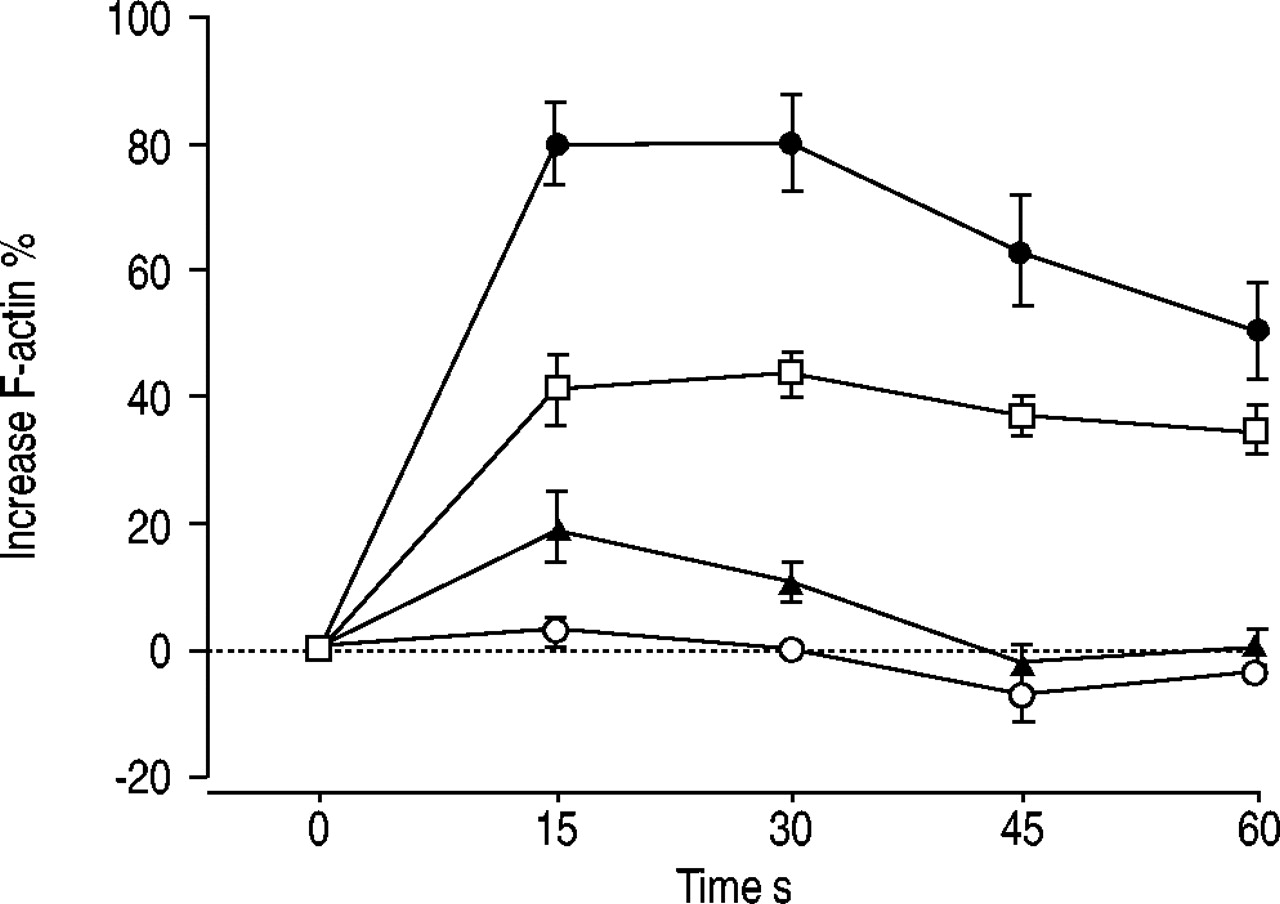
The current authors performed experiments to determine whether the CCL22/CCR4 and CCL17/CCR4 pathways are active in circulating T-cells. The functional readout was actin polymerisation in response to stimulation with CCL22 and CCL17. CCR4 expression was observed on 32.05±1.54% (mean±sem) of peripheral blood CD4+ T-cells from atopic donors (n=5). CCR4-expressing blood CD4+ T-cells, incubated with 10 ng·mL−1 CCL22, exhibited ∼2-fold increase in polymeric filamentous F-actin immediately after 15 s of stimulation (fig. 6⇓). A significant change in the cytoskeletal organisation of T-cells was also observed with CCL17, although this signal was to a lower extent than that observed with CCL22.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Induction of actin polymerisation in peripheral CD4+ T-cells by C-C chemokine receptor (CCR)4 ligands. Blood T-lymphocytes from atopic subjects (n=5) were incubated with C-C chemokine ligand (CCL)22 or CCL17 (10 ng·mL−1) for the indicated times, and actin polymerisation was assessed after labelling with fluorescein isothiocyanate-phalloidin by flow cytometry. Results are expressed as the increase (as a percentage from baseline fluorescence) in mean fluorescence intensity of CD4+ T-cells. •: 10 nM macrophage-derived chemokine (CCL2) CCR4+; □: 10 nM thymus and activation-regulated chemokine (CCL17) CCR4+; ▴: 10 nM CCL2 CCR4-; ○: 10 nM CCL17 CCR4-.
Discussion
The current authors and others have previously documented that CCR4 is up-regulated in the asthmatic airway after allergen challenge 9, 13. This study shows that the ligands for CCR4 are also strongly up-regulated during the late response in atopic asthma. CCL22 and CCL17 levels were increased in the BAL fluid recovered 24 h after endobronchial allergen challenge of atopic asthmatics, and significantly correlated with BAL lymphocytes to some extent, more particularly in patients developing a significant clinical late-phase response. Moreover, the current authors observed, for the first time in vivo, that CCL22 and CCL17 strongly correlated with IL‐5 and IL‐13, which were also induced in BAL upon challenge. CCL22 and CCL17 were not induced upon segmental allergen challenge in nonatopic, healthy controls. In addition, expression of CCL22 and CCL17 in PBMC from atopic subjects was induced by IL‐4 and IL‐13, further supporting the idea that CCR4 ligands are closely related to the Th2-cytokine response. Among Th1-related chemokines, only CXCL10 was significantly increased in post-challenge BAL of asthmatics, and did not correlate with the T-cell influx in to the airway. Finally, the current authors observed that CCR4 is functionally active on circulating T-cells in response to both chemokine ligands.
In asthma, once T-cells have been activated by cognate interactions with antigen-presenting cells, they re-circulate in the bloodstream and may be recruited to the airways upon further allergen exposure/challenge. Although it remains controversial whether the Th2 differentiation occurs systemically (in the lymph nodes draining the airways) or locally, it has been suggested that circulating T-cells from atopic subjects are type 2-biased 18. Since CCR4 appears selectively expressed on Th2 lymphocytes in vitro 10, the observation that CCL22 and CCL17 are strongly up-regulated in the airway after allergen challenge of atopic asthmatics, and correlate with BAL lymphocytes, is consistent with a selective recruitment of circulating Th2 cells. In contrast, CCL1 did not increase after challenge, in line with previous studies showing that CCR8 is up-regulated after challenge of asthmatics to a much lower extent than CCR4 9, and does not play a significant role in the long-term recruitment of T-cells upon repeated allergen exposures in mice 12.
It has been proposed that the cytokine microenvironment, especially that provided by dendritic cells, plays an important role during sensitisation to determine the T-cell profile 19. Similarly, the cytokine milieu of the inflamed asthmatic airway might represent an important factor to drive the tissue recruitment of blood Th2 cells upon further allergen exposure. This hypothesis is supported in the current study, which shows a strong correlation between airway production of CCR4-binding chemokines and Th2 cytokines during the late asthmatic response for the first time in vivo. In addition, the current authors show that in mononuclear cells from atopic subjects, the production of CCL22 and CCL17, in contrast with CCL1, is induced by IL‐4 and IL‐13. It is therefore tempting to speculate that, after allergen sensitisation, further allergen exposure stimulates the mucosal production of Th2 cytokines (by infiltrating T-cells and IgE-bearing leukocytes) that trigger the release of Th2-dependent chemokines, namely CCL22 and CCL17. These chemokines may, in turn, mediate a selective recruitment to the airway of circulating Th2 lymphocytes expressing CCR4. Both the induction of CCL22 and CCL17 by IL‐4 or IL‐13, and the selective attraction of Th2 cytokine-producing cells by these chemokines (as an amplification circuit), may account for the correlation observed in this study between the Th2-cytokine response and CCR4 ligands.
This study was not designed to identify the cellular source(s) of the chemokines released in the airways upon allergen challenge. The current authors' in vitro system was based on the fact that mononuclear cells (consisting of monocytes and lymphocytes) represent a major source of CCR4 ligands. However, alternative sources have been described, such as airway epithelial cells or smooth muscle cells. These findings using PBMC are in line with those of Andrew et al. 20, showing that CCL22 is inducible in monocytes upon IL‐4 or IL‐13 stimulation, as observed in T-cells; while macrophages constitutively produce CCL22, which is further increased by IL‐4 and reduced by IFN‐γ. It was shown that CCL17 may also be produced by bronchial epithelial cells stimulated by tumour necrosis factor (TNF)‐α and IFN‐γ 21. Dendritic cells activated by TNF‐α can produce CCL22 and CCL17, whilst producing CCL1 and CXCL10 when activated through Toll-like receptors 22. CXCR3-binding chemokines including CXCL10 can be released by mononuclear cells and/or by activated bronchial epithelial cells 23, although less information is available regarding the regulation of production of CXCR3 ligands. Preliminary data, including information in the present study, confirm that production of CXCL10 is mainly dependent on IFN‐γ, at least in peripheral mononuclear cells.
The current authors' finding that CXCL10 is also significantly up-regulated in atopic asthmatics upon airway allergen challenge may appear intriguing. CXCL10 has been mainly implicated in Th1-mediated diseases, such as sarcoidosis 24 or chronic obstructive pulmonary disease 25, which is consistent with the observation that CXCR3 (CXCL10 receptor) is predominantly expressed by Th1 cells 26. However, recent studies have also indicated a role for CXCL10 in asthma. First, it has been reported that airway expression of CXCL10 is increased in human asthma 27 and in mice sensitised to ovalbumin 28. Secondly, in murine models of asthma, a dual effect of CXCL10 has been identified on AHR and tissue eosinophilia 28, 29. Thirdly, it was shown that CXCL10 is induced in keratinocytes by both Th1- and Th2-derived supernatants 30. In blood mononuclear cells from atopics, the authors observed in this study that CXCL10 expression is only dependent on IFN‐γ (and inhibited by IL‐13), as observed for CXCL9. However, CXCL10 was not significantly correlated with IFN‐γ in the BAL from challenged asthmatics (not shown), as previously observed in mice 28, suggesting that other factors could trigger CXCL10 expression in vivo. Interestingly, in adoptive transfer experiments, Stephens et al. 31 showed that viral activation of Th1 cells can enhance subsequent recruitment of allergen-specific Th2 cells upon lung viral challenge. Moreover, it has been recently suggested that, in lymph nodes, Th2 cells can somehow influence T-cells located in the same microenvironment, including Th1 cells 32, indicating that the T-cell programme is flexible during developing immune responses and depends on signals provided locally. Taken together, these data suggest that CXCL10/CXCR3 might represent another chemokine pathway involved in asthma, but its role and relevance in humans remain to be determined. The fact that CCR4 ligands only modestly correlated with T-lymphocytes further suggests that other chemokine pathways are involved. Moreover, the CXCR3 axis might also play a role in asthma through actions on other cell types, such as γδ T-cells 33 or mast cells 34, which express CXCR3 and respond to its ligands.
An alternative explanation regarding IP‐10 up-regulation in asthma relates to the selective recruitment of Th1 cells. Thus, IFN‐γ-producing Th1-type cells detected in peripheral blood of atopic patients have been suggested to also play a role in asthma 35, 36, especially during the symptomatic phases of the disease. It could be hypothesised that a Th2 orientation in asthma mainly characterises the immune response during the sensitisation process and the acute reaction to allergen exposure; whereas, chronic inflammation could be underlined by a less specific T-cell profile. This view is supported by the clinical observation that Th2- (e.g. allergens) and Th1- (e.g. viruses) driving factors may both represent triggers of asthma exacerbations.
Finally, it was observed that a consistent proportion (∼30%) of circulating CD4+ T-lymphocytes express CCR4, in line with previous studies 37, 38. Evaluation of actin polymerisation in circulating blood T-cells expressing CCR4 confirmed that these cells are functionally responsive to CCL22 and CCL17, and could migrate to the airways upon stimulation by these chemokines released locally after allergen exposure in atopic asthmatics.
In summary, this study further highlights the C-C chemokine receptor 4 pathway as mediating the recruitment of T-lymphocytes to the inflamed airway in atopic asthma. The ligands for C-C chemokine receptor 4: 1) are strongly up-regulated within the airways from atopic asthmatics after allergen challenge; 2) contribute to the T-cell influx to the airways; and 3) are closely related to the T-helper 2-cytokine response. The C-C chemokine receptor 4 pathway could therefore participate in the mechanisms underlying a selective recruitment of circulating T-helper 2 cells to the airways. This study also shows that various chemokine pathways could be implicated and supports the idea that targeting several chemokine receptors may be appropriate 14 to dampen T-cell infiltration in chronic asthma.
- Received September 10, 2003.
- Accepted January 6, 2004.
- © ERS Journals Ltd
References









