
Abstract
Objective
To investigate the effect of mechanical ventilation on alveolar fibrinolytic capacity.
Design and setting
Randomized controlled animal study in 66 Sprague-Dawley rats.
Subjects and interventions
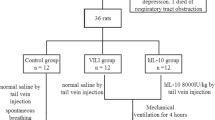
Test animals received intratracheal fibrinogen and thrombin instillations; six were killed immediately (fibrin controls), and the others were allocated to three ventilation groups (ventilation period: 225 min) differing in positive inspiratory pressure and positive end-expiratory pressure, respectively: group 1, 16 cmH2O and 5 cmH2O (n=17); group 2, 26 cmH2O and 5 cmH2O (n=16); group 3, 35 cmH2O and of 5 cmH2O (n=17). Ten animals that had not been ventilated served as healthy controls.
Measurements and results
After animals were killed, we measured D-dimers, plasminogen activator inhibitor (PAI) 1, and tumor necrosis factor α in the bronchoalveolar lavage fluid and calculated lung weight and pressure/volume (P/V) plots. The median D-dimer concentration (mg/l) decreased with increasing pressure amplitude (192 in group 1, IQR 119; 66 in group 2, IQR 107; 29 in group 3, IQR 30) while median PAI-1 (U/ml) increased (undetectable in group 1; 0.55 in group 2, IQR 4.55; 3.05 in group 3, IQR 4.85). PAI-1 level was correlated with increased lung weight per bodyweight (Spearman’s rank correlation 0.708). Tumor necrosis factor α concentration was not correlated with PAI-1 level.
Conclusions
Alveolar fibrinolytic capacity is suppressed during mechanical ventilation with high pressure amplitudes due to local production of PAI-1.
Similar content being viewed by others

Introduction
Intra-alveolar fibrin-rich hyaline membranes are the distinguishing microscopic feature of numerous acute diffuse lung diseases including acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) [1, 2]. In ALI acute insults (e.g., sepsis and pneumonia) trigger an inflammatory response causing increased capilloalveolar permeability for, for example, fibrinogen. Alveolar cells (i.e., macrophages and epithelial cells) are stimulated directly by bacterial endotoxins or by proinflammatory mediators, with a central role played by tumor necrosis factor (TNF) α in producing procoagulant (i.e., factor VII and tissue factor) and antifibrinolytic proteins resulting in alveolar fibrin depositions. The degree and persistence of alveolar fibrin depends predominantly on local production of plasminogen activator inhibitor (PAI) 1, the direct antagonist of the fibrinolytic system [3, 4, 5, 6, 7, 8].
Alveolar fibrin membranes inactivate and incorporate surfactant leading to disturbance of gas exchange, impairment of lung mechanics, and aggravation of capilloalveolar leakage [9]. Furthermore, they may contribute to the inflammatory response observed in ALI and also initiate a fibroproliferate state with de novo inflammation, fibrosis, and long-term compromised pulmonary function in infants, children, and adults after acute respiratory failure [10]. Under normal circumstances fibrin membranes are resolved within minutes by plasmin [11, 12]. However, during ALI additional insults (e.g., hemorrhagic shock, infections) may prolong disturbed alveolar fibrin turnover [13]. In patients with ALI mechanical ventilation, although mandatory, is considered a secondary insult for lung damage. Mechanical ventilation with high tidal volume and/or high pressure amplitude without sufficient positive end-expiratory pressures (PEEP) causes shear stress on lung tissue which stimulates de novo production of proinflammatory cytokines in, for example, alveolar cells [14, 15, 16]. This so-called mechanotransduction-related lung inflammation is believed to be the major determinant of ventilation-induced lung injury (VILI) and its associated morbidity and mortality [17, 18, 19, 20, 21]. Despite this understanding, there is still a tremendous lack of knowledge about the mechanisms involved [22]. For example, it has not yet been investigated whether mechanical ventilation interferes with alveolar fibrin turnover or with the natural capacity of alveolar fibrin resolution [23, 24].
Therefore we investigated the effects of mechanical ventilation with various high pressure amplitudes on alveolar fibrinolytic capacity in a rat model of iatrogenic intra-alveolar fibrin formation.
Materials and methods
The study was approved by the Animal Committee of the Erasmus University, Rotterdam. Care and handling of the animals were in accordance with European Union guidelines. The experiments were performed at the Department of Anesthesiology, Erasmus Medical Centre Faculty in male Sprague-Dawley rats (n=66) with a bodyweight (BW) of 287±3 g (IFFA Credo, The Netherlands).
Experimental protocol
Rats were anesthetized with nitrous oxide, oxygen and isoflurane (65/33/2%), tracheotomized, and had a catheter inserted into a carotid artery. Anesthesia was maintained with hourly intraperitoneal injections of pentobarbital sodium (60 mg/kg; Nembutal, Algin, Maassluis, The Netherlands). Muscles were relaxed with 2 mg/kg pancuronium bromide (Pavulon; Organon Technika, Boxtel, The Netherlands) intramuscularly hourly. After muscle relaxation all animals were connected to a ventilator (Servo Ventilator 300; Siemens-Elema, Solna, Sweden) in a pressure-controlled mode with positive inspiratory pressure (PIP) of 12 cmH2O and PEEP of 2 cmH2O, frequency of 30 breaths/min, inspiratory to expiratory ratio of 1:2, and fractional inspired oxygen tension of 1.0. Body temperature was kept within normal range by means of a heating pad.
After 15-min stabilization intra-alveolar fibrin formation was generated by intratracheal instillation of human fibrinogen (40 mg/kg, Sanquin, Amsterdam, The Netherlands) followed 1 min thereafter by human thrombin (10 µg/kg, a generous gift from Dr. W. Kisiel, University of New Mexico, Albuquerque, N.M., USA). Both fibrin and thrombin were diluted to a maximum volume of 1 ml (with normal saline). Doses of fibrinogen and thrombin were derived from the literature [25] and our own pilot experiments. For optimal fibrin deposition PIP was increased to 26 cmH2O just before, and intratracheal air (10 ml/kg BW) was instilled three times just after instillation of fibrin. After instillation PIP was adjusted immediately according to the allocation of the animals to the different treatment groups. Arterial blood gases were taken from a carotid artery; just before fibrinogen/thrombin instillation and additional samples were taken at 15 min and every 30 min thereafter. Blood gases were measured using conventional methods (ABL555, Radiometer, Copenhagen, Denmark). Blood pressure was monitored through the carotid artery before instillation and at 15 and every 30 min thereafter for 225 min after fibrin instillation. Before measurements the rats were killed after 225 min of mechanical ventilation with an intra-arterial overdose of pentobarbital sodium (600 mg/kg BW pentobarbital sodium).
Study groups
Ten rats were killed immediately after the surgical procedure and served as healthy control rats. Animals which received instillations of fibrinogen/thrombin were randomly allocated to four groups. In the group designated as “fibrin controls” six animals were killed 5 min after the instillation procedure; these served as controls for the ventilation groups with respect to the D-dimer and PAI-1 measurements. In the three other groups animals were ventilated with various high pressure amplitudes: group 1, PIP 16 cmH2O and PEEP 5 cmH2O (n=17); group 2, PIP 26 cmH2O and PEEP 5 cmH2O (n=16); and group 3, PIP 35 cmH2O and PEEP 5 cmH2O (n=17). The ranges of tidal volumes (ml/kg) for each group were: group 1 13–16, group 2 22–26, and group 3 28–32.
Bronchoalveolar lavage and lung mechanics
The time at which bronchoalveolar lavage was performed was defined at the moment when rats died prior to or at the end of the study period at 225 min. After the animals were killed, the thorax and diaphragm were opened (to eliminate the effect of chest wall compliance and intra-abdominal pressure) and a static pressure-volume (P/V) plot from the lung was calculated using conventional techniques [26]. Maximal compliance (Cmax) was defined as the steepest part of the P/V deflation curve and was determined separately for each animal. Measurements were not completed in 14 rats due to air leakage during the test. After the P/V plot bronchoalveolar lavage was performed with normal saline (30 ml/kg heated to 37°C) and reaspirated three times, and the recovered supernatant fluid was stored at −80°C until further processing. No bronchoalveolar lavage was performed in the animals in which histology was performed (one per group).
Fibrinolysis measurements
D-dimer levels in the bronchoalveolar lavage fluid (BALF) were quantitated by a sandwich-type enzyme-linked immunosorbent assay (Asserachrom D-dimer, Diagnostica Stago, Asnières-sur-Seine, France). PAI-1 activity in BALF was determined on an automated coagulation analyzer (Behring Coagulation System, Marburg, Germany) with reagents and protocols from the manufacturer. This assay determines the urokinase-inhibiting activity of the sample. The remaining urokinase is then assayed by activating plasminogen to plasmin and subsequent determination of plasmin’s chromogenic activity. The assay is independent of variable concentrations of plasminogen, α2-antiplasmin, and fibrinogen in the sample. We cannot exclude that other urokinase inhibitors were determined; however, PAI-1 is the major inhibitor of urokinase. Furthermore, the results of this assay are correlated very well with those of methods based on the inhibition of urokinase-type plasminogen activator. The concentration of urokinase-type plasminogen activator in the BALF was not assessed, but with these high concentrations of PAI-1 these are most likely extremely low.
TNF-α measurements
Levels of TNF-α in BALF were measured by commercial enzyme-linked immunosorbent assay (Rat TNF-α/TNFSF2, catalogue no. DY510, R&D Systems, Abingdon, UK).
Lung histology
To determine alveolar fibrin depositions samples were taken from all lung lobes and 30 fields were analyzed. The analyzing pathologist was not informed about the study purpose and was asked only to prepare rat lungs for illustration of alveolar fibrin deposition. For fixation 4% paraformaldehyde was instilled in the lungs under a positive airway pressure for lung recruitment of 10 cmH2O. The specimens were embedded in paraffin, sectioned in tissue blocks from all lobes, and stained with hematoxylin and eosin.
For fibrin staining slides were deparaffinized, and endogenous peroxidase activity was quenched by a solution of methanol/0.03% H2O2 (Merck, Darmstadt, Germany). After digestion with a solution of pepsine 0.25% (Sigma, St. Louis, Mo., USA) in 0.01 M HCl, the sections were incubated in 10% normal goat serum (Dako, Glostrup, Denmark) and then exposed to biotin-labeled goat anti-human fibrinogen antibody (Ixell, Accurate Chemical & Scientific, Westbury, N.Y., USA). After washes slides were incubated in a streptavidin-ABC solution (Dako) and developed using 1% H2O2 and 3.3′-diaminobenzidin-tetra-hydrochloride (Sigma) in Tris-HCl. The sections were mounted in glycerin gelatin and counterstained with hematoxylin.
Statistics
Statistical analysis was performed using SPSS version 11.5 (SPSS, Chicago, Ill., USA). Data of physiological parameters are presented as mean ± standard error. Intergroup differences were analyzed by analysis of variance and Bonferroni’s post-hoc test; differences at the level of p<0.05 (one-tailed) were considered significant. For multiple comparisons Bonferroni’s correction was used: 1−α=0.05/3=0.017 and p<0.017 (one-tailed) was considered significant. Data on D-dimer and PAI-1 are presented as median and interquartile range (IQR), and on the Kruskal-Wallis test corrected for multiple comparisons differences at the level of p<0.017 (one-tailed) were considered significant. The Mann-Whitney U test for comparisons between two groups was performed, and the level of p<0.05 (one-tailed) was considered significant. In each ventilation group we calculated post hoc the correlations between D-dimer and PAI-1 concentrations in BALF with survival time. Linear regression analysis was performed to evaluate the relationship between ventilation pressure, D-dimer, PAI-1, and lung weight (g)/kg bodyweight (BW), with p<0.05 being considered significant.
Results
Lung injury
Table 1 shows the physiological parameters and their differences between the ventilation groups. To document the traumatic effects of mechanical ventilation on lungs we compared changes in lung weight and P/V curves of the ventilation groups with those of healthy controls (unventilated animals). We found that the mean lung weights per BW in groups 2 and 3 were significantly higher than those in healthy controls and group 1 (Table 2). In groups 2 and 3 lung function was impaired, as demonstrated by significantly lower values for the P/V plots (Fig. 1); however, no differences were found between group 1 and healthy controls. The higher relative lung weight in “fibrin controls” was due to the fact that they were killed 5 min after the instillation procedure (2 ml volume load).

Effect of mechanical ventilation on lung function: P/V plots, lines connect group means ± SE at each step of 5 cmH2O increase in airway pressure. X Healthy controls without fibrin instillation; enclosed X healthy controls with fibrin instillation; squares group 1 (16/5 cmH2O); circles group 2 (26/5 cmH2O); diamonds group 3 (35/5 cmH2O). A Healthy controls without vs. controls with fibrin instillation; mean difference 117±41 ml/kg. B Healthy controls vs. group 1; mean difference 66±33 ml/kg (p=0.21). C Healthy controls vs. group 2; mean difference 170±34 ml/kg (p<0.001). D Healthy controls vs. group 3; mean difference 241±33 ml/kg (p<0.001). Mean difference ± SE between group 2 (B) and 3 (C) 70±14 ml/kg (p=0.023) and that between controls with fibrin instillation and group 3 124±33 ml/kg (p=0.005). *p<0.05 (analysis of variance)
Fibrinolytic capacity
As shown in Fig. 2, fibrin-rich material was deposited in alveoli in fibrin-treated animals. Levels of D-dimers were below detection limit in healthy control rats (no instillation of fibrin, no mechanical ventilation). “Fibrin controls” (animals killed 5 min after the instillation procedure) showed a small but significant increase in D-dimer in BALF compared to healthy controls rats: 0 vs. 3.4 mg/l (IQR 3.2). In animals that had been mechanically ventilated the median BALF level of D-dimer (median 192 mg/l; IQR 119) was highest in group 1 (lowest pressure amplitude). A further increase in pressure amplitude (groups 2 and 3) resulted in significantly lower D-dimer concentrations in BALF (Fig. 3A, group 2 66 mg/l, IQR 107; group 3 29 mg/l, IQR 30); p<0.001, Kruskal Wallis). BALF concentrations of PAI-1 were below detection limit in all control rats (with and without fibrinogen/thrombin instillation) and in rats ventilated with the lowest pressure amplitude (group 1; Fig. 3B). In inverse relationship to the decrease in D-dimer levels, median BALF concentrations of PAI-1 increased the higher pressure amplitude during mechanical ventilation (group 2, 0.55 U/ml, IQR 4.55, p=0.007; group 3, 3.05 U/ml, IQR 4.85, p=0.001, Fig. 3B).

Microscopy of representative fibrin-stained paraffin sections of rat lung tissue. A Healthy animals, normal lung tissue, no fibrin in the alveolar space. B Group 1 (16/5 cmH2O) after 225 min of ventilation C Group 2 (26/5 cmH2O) after 195 min of ventilation. D: Group 3 (35/5 cmH2O) after 135 min of ventilation. Arrows Intra-alveolar fibrin depositions; original magnification ×140

Effect of mechanical ventilation on D-dimer and PAI-1 concentrations in BALF. Values are median, boxes represent interquartile range (25th–75th percentiles),* p<0.05 was considered significant (Mann-Whitney U test). N indicates the number of animals. A: D-dimer, * p<0.05 group 1, II and III vs. both control groups. B: PAI-1, * p<0.05 group 2 and III vs. group 1 and both control groups
Analysis of whether the measurement of fibrinolytic parameters depended on the duration of mechanical ventilation (time dependence) is shown in Fig. 4. In a post-hoc analysis no intragroup differences were found in D-dimer or PAI-1 regarding the time at which BAL measurements were performed (p>0.05, analysis of variance). Linear regression revealed that PAI-1 levels were correlated with the increase in lung weight per BW (R2=0.496; Spearman’s rank correlation coefficient 0.708, p<0.001; Fig. 5).

Median D-dimer (A) and PAI-1 (B) levels at time of death (survival time, min) in BALF are shown for the three ventilation groups. Boxes Interquartile range (25th–75th percentiles); N number of animals used for analysis at each time point. There were no significant differences at the different time points between ventilation groups (p>0.05, analysis of variance)

Linear regression: correlation between lung weight/body weight (BW) and PAI-1 concentrations in BALF (p<0.001, R2=0.496, Spearman’s rank correlation coefficient 0.708)
Levels of TNF-α
TNF-α was undetectable in all controls, as in the majority of ventilated rats (12/16 in group 1, 4/15 in group 2, 10/16 in group 3). Group means did not differ significantly (0 pg/ml in group 1, IQR 1.5; 36 pg/ml in group 2, IQR 212; 0 pg/ml in group 3, IQR 58).
Discussion
The present study analyzed the effect of mechanical ventilation on the alveolar fibrinolytic capacity in an in vivo model of iatrogenic intra-alveolar fibrin formation. Interestingly, mechanical ventilation with a small pressure amplitude (16/5 cmH2O, group 1) neither altered lung weight nor caused production of PAI-1. These animals showed the highest D-dimer concentrations in BALF. In contrast, mechanical ventilation with larger pressure amplitudes (groups 2 and 3) resulted in significantly greater lung weight and depressed intra-alveolar fibrinolysis. Since high levels of PAI-1 were directly related to the severity of lung injury (i.e., lung weight per BW), PAI-1 seems to be responsible for this observation. This is the first study to demonstrate that mechanical ventilation alone may depress the alveolar fibrinolytic capacity by upregulation of PAI-1 and thus may enhance fibrin deposition in patients with ALI.
Experimental model
Our experimental model to create intra-alveolar fibrin formation is similar in some aspects to a model described previously [25]; doses were derived from the latter study and from pilot experiments in our laboratory. The illustrations from microscopy (Fig. 2) and the large amounts of D-dimers in BALF (Fig. 3A) indicate intra-alveolar fibrin formation in all groups subjected to fibrin instillation. Because all animals received the same amounts of fibrinogen/thrombin, the extent of fibrin degradation (D-dimers) may be considered as an indication of alveolar fibrinolysis [27]. We cannot exclude that intra-alveolar instillation of thrombin caused some alveolar damage. Thrombin might affect the endothelial monolayer and increase permeability, enhancing the formation of edema [28]. However, because all groups received the same amount of thrombin, the observed differences between the groups were caused by the differences in ventilation pressure.
Lung injury
The aim of the present study was to investigate the effects of lung injury on alveolar fibrinolysis. Lung injury was achieved by mechanical ventilation applying high pressure amplitudes, as in previous studies [29, 30]. The significant decrease in PaO2, survival time, P/V plots, and the increase in lung weight per BW in groups with larger pressure amplitudes (groups 2 and 3) clearly document VILI.
Depressed fibrinolytic capacity
Fibrinogen/thrombin instillations resulted in fibrin controls in a measurable fibrin breakdown within 5 min after the instillation procedure without PAI-1 production (Fig. 3A). In ventilated animals fibrinolysis was most preserved in animals of group 1 (highest levels of D-dimer and no PAI-1 production). In groups 2 and 3, however, PAI-1 production increased and fibrin breakdown decreased. Further, fibrin breakdown did not depend on the duration of mechanical ventilation (survival time) (Fig. 4) but was correlated with the size of the pressure amplitude and the extent of lung injury (Fig. 5). With respect to the large amounts of fibrinogen/thrombin which had been instilled, the results of the present study must be interpreted qualitatively and confirm particularly that mechanical ventilation may disturb alveolar fibrin breakdown. From clinical and experimental lung injury studies (e.g., induced by sepsis, pneumonia) we know that intra-alveolar fibrinolysis is downregulated due to increased production of PAI-1 [3, 6, 31]. Indeed, groups 2 and 3 showed increased PAI-1 levels whereas in group 1 PAI-1 was not detectable (Fig. 3B). This demonstrates that aggressive mechanical ventilation with high pressure amplitudes causes PAI-1 production, which downregulates alveolar fibrinolysis. Interestingly, PAI-1 was not detectable in any of the control rats or in rats of group 1 (no increase in lung weight and normal P/V plots). This finding suggests that mechanical ventilation with low pressure amplitude might preserve alveolar fibrinolytic capacity.
Because we studied the effects of mechanical ventilation in previously healthy rats, it is unclear whether there may be a similar PAI-1 production in lungs that have been already injured, for example, by sepsis or pneumonia. The possibility of additive effects on PAI-1 synthesis in preinjured lungs can be expected because elevated PAI-1 levels in BALF from ALI/ARDS lungs [31] and infected lungs [32] have been demonstrated.
The exact mechanisms of PAI-1 production due to mechanical ventilation are not known. From experiments in sepsis or pneumonia-induced ALI it was concluded that PAI-1 related depressed fibrinolysis depends on the interplay between inflammation and coagulation [4, 33], with TNF-α as an important link. However, in the present study mechanical ventilation for 225 min triggered TNF-α production in some animals only. No correlation between either TNF-α, PAI-1, or fibrin degradation was found. Our findings reflect the current debate about differences in cytokine response in VILI. Obviously the role of TNF-α and possibly other inflammatory mediators seems to depend on the experimental model applied [30, 34]. With respect to our objective, their role of up- or downregulation of ventilation-induced PAI-1 production remains unclear and needs further elucidation.
In this context, other mechanisms, unrelated to inflammatory mediators, may be responsible for mechanical ventilation-associated depressed fibrinolytic capacity [22, 35]. The newest concept of VILI considers plasma membrane wounding due to shear stress as the initial trigger for multiple intracellular signal induction pathways (e.g., plasma membrane lipid trafficking, nuclear factor κ B, proteinkinase systems) and the synthesis of a variety of mediators and proteases [36, 37]. Links to the fibrinolytic system (e.g., PAI-1 production) may be possible, which is supported by new insights from other research areas where cell wounding is considered the major determinant of PAI-1 mediated fibrin formation. In idiopathic pulmonary fibrosis, alveolar epithelial cell damage and pathological wound repair directly initiate (without inflammation) overexpression of PAI-1 and persistence of fibrin [38]. Furthermore, it has been shown that PAI-1 production is triggered by transforming growth factor-β1 after mechanical stretch in lung epithelial cells [3, 39, 40, 41]. The fact that cells in general react on mechanical stress with downregulation of the fibrinolytic system has also been demonstrated on vascular endothelial cells, where flow-induced shear stress triggered PAI-1 production [42]. The clinical importance of an intra-alveolar antifibrinolytic milieu in mechanically ventilated patients with ALI has recently been underscored by the results of a study in adult patients with ARDS in which high levels of PAI-1 in pulmonary edema fluid were associated with higher mortality [43]. Our findings clearly suggest that mechanical ventilation is one of the factors explaining this latter observation.
Conclusion
Alveolar fibrinolytic capacity can be suppressed (PAI-1 dependently) by mechanical ventilation with high pressure amplitudes. This finding may add new information to the pathogenesis of VILI. Our results need to be confirmed by investigations in other lung injury models with intrinsic fibrin formation mimicking diseases such as pneumonia or sepsis.
Reference
Liebermann J, Ambrus CM (1963) Fibrinolysin, fibrin and hyaline membranes. Pediatrics 32:940–942
Bachofen M, Weibel ER (1982) Structural alterations of lung parenchyma in the adult respiratory distress syndrome. Clin Chest Med 3:35–56
Idell S (2003) Coagulation, fibrinolysis, and fibrin deposition in acute lung injury. Crit Care Med 31:S213–S220
Levi M, Schultz MJ, Rijneveld AW, Van der Poll T (2003) Bronchoalveolar coagulation and fibrinolysis in endotoxemia and pneumonia. Crit Care Med 31:S238–S242
Bertozzi P, Astedt B, Zenzius L, Lynch K, LeMaire F, Zapol W, Chapman HA Jr (1990) Depressed bronchoalveolar urokinase activity in patients with adult respiratory distress syndrome. N Engl J Med 322:890–897
Idell S (1994) Extravascular coagulation and fibrin deposition in acute lung injury. New Horiz 2:566–574
Barazzone C, Belin D, Piguet PF, Vassalli JD, Sappino AP (1996) Plasminogen activator inhibitor-1 in acute hyperoxic mouse lung injury. J Clin Invest 98:2666–2673
Van der Poll T, de Jonge E, Levi M (2001) Regulatory role of cytokines in disseminated intravascular coagulation. Semin Thromb Hemost 27:639–651
Seeger W, Stohr G, Wolf HR, Neuhof H (1985) Alteration of surfactant function due to protein leakage: special interaction with fibrin monomer. J Appl Physiol 58:326–338
Fukuda Y, Ishizaki M, Masuda Y, Kimura G, Kawanami O, Masugi Y (1987) The role of intraalveolar fibrosis in the process of pulmonary structural remodeling in patients with diffuse alveolar damage. Am J Pathol 126:171–182
Chung SI, Lee SY, Uchino R, Carmassi F (1996) Factors that control extravascular fibrinolysis. Semin Thromb Hemost 22:479–488
Gunther A, Kalinowski M, Elssner A, Seeger W (1994) Clot-embedded natural surfactant: kinetics of fibrinolysis and surface activity. Am J Physiol 267:L618–L624
Fan J, Kapus A, Li YH, Rizoli S, Marshall JC, Rotstein OD (2000) Priming for enhanced alveolar fibrin deposition after hemorrhagic shock: role of tumor necrosis factor. Am J Respir Cell Mol Biol 22:412–421
Tremblay L, Valenza F, Ribeiro SP, Li J, Slutsky AS (1997) Injurious ventilatory strategies increase cytokines and c-fos m-RNA expression in an isolated rat lung model. J Clin Invest 99:944–952
Dreyfuss D, Saumon G (1998) Ventilator-induced lung injury: lessons from experimental studies. Am J Respir Crit Care Med 157:294–323
Ranieri VM, Suter PM, Tortorella C, De Tullio R, Dayer JM, Brienza A, Bruno F, Slutsky AS (1999) Effect of mechanical ventilation on inflammatory mediators in patients with acute respiratory distress syndrome: a randomized controlled trial. JAMA 282:54–61
Slutsky AS, Tremblay LN (1998) Multiple system organ failure. Is mechanical ventilation a contributing factor? Am J Respir Crit Care Med 157:1721–1725
Slutsky AS (1999) Lung injury caused by mechanical ventilation. Chest 116:9S-15S
Acute Respiratory Distress Syndrome Network (2000) Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. N Engl J Med 342:1301–1308
Amato MB, Barbas CS, Medeiros DM, Magaldi RB, Schettino GP, Lorenzi-Filho G, Kairalla RA, Deheinzelin D, Munoz C, Oliveira R, Takagaki TY, Carvalho CR (1998) Effect of a protective-ventilation strategy on mortality in the acute respiratory distress syndrome. N Engl J Med 338:347–354
Brochard L, Roudot-Thoraval F, Roupie E, Delclaux C, Chastre J, Fernandez-Mondejar E, Clementi E, Mancebo J, Factor P, Matamis D, Ranieri M, Blanch L, Rodi G, Mentec H, Dreyfuss D, Ferrer M, Brun-Buisson C, Tobin M, LeMaire F (1998) Tidal volume reduction for prevention of ventilator-induced lung injury in acute respiratory distress syndrome. The Multicenter Trail Group on Tidal Volume reduction in ARDS. Am J Respir Crit Care Med 158:1831–1838
Frank JA, Matthay MA (2003) Science review: mechanisms of ventilator-induced injury. Crit Care 7:233–241
Dreyfuss D, Saumon G (1993) Role of tidal volume, FRC, and end-inspiratory volume in the development of pulmonary edema following mechanical ventilation. Am Rev Respir Dis 148:1194–1203
Parker JC, Hernandez LA, Longenecker GL, Peevy K, Johnson W (1990) Lung edema caused by high peak inspiratory pressures in dogs. Role of increased microvascular filtration pressure and permeability. Am Rev Respir Dis 142:321–328
Schermuly RT, Gunther A, Ermert M, Ermert L, Ghofrani HA, Weissmann N, Grimminger F, Seeger W, Walmrath D (2001) Conebulization of surfactant and urokinase restores gas exchange in perfused lungs with alveolar fibrin formation. Am J Physiol Lung Cell Mol Physiol 280:L792–L800
Lachmann B, Robertson B, Vogel J (1980) In vivo lung lavage as an experimental model of the respiratory distress syndrome. Acta Anaesthesiol Scand 24:231–236
Sato N, Takahashi H, Shibata A (1995) Fibrinogen/fibrin degradation products and D-dimer in clinical practice: interpretation of discrepant results. Am J Hematol 48:168–174
Coughlin SR (2000) Thrombin signalling and protease-activated receptors. Nature 407:258–264
Haitsma JJ, Uhlig S, Goggel R, Verbrugge SJ, Lachmann U, Lachmann B (2000) Ventilator-induced lung injury leads to loss of alveolar and systemic compartmentalization of tumor necrosis factor-alpha. Intensive Care Med 26:1515–1522
Verbrugge SJ, Uhlig S, Neggers SJ, Martin C, Held HD, Haitsma JJ, Lachmann B (1999) Different ventilation strategies affect lung function but do not increase tumor necrosis factor-alpha and prostacyclin production in lavaged rat lungs in vivo. Anesthesiology 91:1834–1843
Gunther A, Mosavi P, Heinemann S, Ruppert C, Muth H, Markart P, Grimminger F, Walmrath D, Temmesfeld-Wollbruck B, Seeger W (2000) Alveolar fibrin formation caused by enhanced procoagulant and depressed fibrinolytic capacities in severe pneumonia. Comparison with the acute respiratory distress syndrome. Am J Respir Crit Care Med 161:454–462
Schultz MJ, Millo J, Levi M, Hack CE, Weverling GJ, Garrard CS, Van der Poll T (2004) Local activation of coagulation and inhibition of fibrinolysis in the lung during ventilator associated pneumonia. Thorax 59:130–135
Idell S (1995) Coagulation, fibrinolysis and fibrin deposition in lung injury and repair.743–776
Ricard JD, Dreyfuss D, Saumon G (2001) Production of inflammatory cytokines in ventilator-induced lung injury: a reappraisal. Am J Respir Crit Care Med 163:1176–1180
Ricard JD, Dreyfuss D, Saumon G (2002) Ventilator-induced lung injury. Curr Opin Crit Care 8:12–20
Uhlig U, Haitsma JJ, Goldmann T, Poelma DL, Lachmann B, Uhlig S (2002) Ventilation-induced activation of the mitogen-activated protein kinase pathway. Eur Respir J 20:946–956
Vlahakis NE, Hubmayr RD (2003) Response of alveolar cells to mechanical stress. Curr Opin Crit Care 9:2–8
Geiser T (2003) Idiopathic pulmonary fibrosis-a disorder of alveolar wound repair? Swiss Med Wkly 133:405–411
Vassalli JD, Sappino AP, Belin D (1991) The plasminogen activator/plasmin system. J Clin Invest 88:1067–1072
Idell S, Zwieb C, Boggaram J, Holiday D, Johnson AR, Raghu G (1992) Mechanisms of fibrin formation and lysis by human lung fibroblasts: influence of TGF-beta and TNF-alpha. Am J Physiol 263:L487–L494
Yamamoto H, Teramoto H, Uetani K, Igawa K, Shimizu E (2002) Cyclic stretch upregulates interleukin-8 and transforming growth factor-beta1 production through a protein kinase C-dependent pathway in alveolar epithelial cells. Respirology 7:103–109
Cheng JJ, Chao YJ, Wung BS, Wang DL (1996) Cyclic strain-induced plasminogen activator inhibitor-1 (PAI-1) release from endothelial cells involves reactive oxygen species. Biochem Biophys Res Commun 225:100–105
Prabhakaran P, Ware LB, White KE, Cross MT, Matthay MA, Olman MA (2003) Elevated levels of plasminogen activator inhibitor-1 in pulmonary edema fluid are associated with mortality in acute lung injury. Am J Physiol Lung Cell Mol Physiol 285:L20–L28
Acknowledgements
The authors thank S. Krabbendam (Department of Anesthesiology, Erasmus MC Faculty, Rotterdam) for expert technical assistance, M. Weijne for laboratory assistance (Department of Vascular Medicine, University of Amsterdam), H. Thygesen (Department of Biostatistics and Epidemiology, University of Amsterdam) for statistical advice, L.A. Noorduyn (Department of Pathology, University of Amsterdam) for expert histological preparation, and Laraine Visser-Isles for English language editing.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dahlem, P., Bos, A.P., Haitsma, J.J. et al. Alveolar fibrinolytic capacity suppressed by injurious mechanical ventilation. Intensive Care Med 31, 724–732 (2005). https://doi.org/10.1007/s00134-005-2588-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00134-005-2588-2