Article Text

Abstract
Background Seasonal influenza A infection affects a significant cohort of the global population annually, resulting in considerable morbidity and mortality. Therapeutic strategies are of limited efficacy, and during a pandemic outbreak would only be available to a minority of the global population. Over-the-counter medicines are routinely taken by individuals suffering from influenza, but few studies have been conducted to determine their effectiveness in reducing pulmonary immunopathology or the influence they exert upon the generation of protective immunity.
Methods A mouse model of influenza infection was utilised to assess the efficacy of paracetamol (acetaminophen) in reducing influenza-induced pathology and to examine whether paracetamol affects generation of protective immunity.
Results Administration (intraperitoneal) of paracetamol significantly decreased the infiltration of inflammatory cells into the airway spaces, reduced pulmonary immunopathology associated with acute infection and improved the overall lung function of mice, without adversely affecting the induction of virus-specific adaptive responses. Mice treated with paracetamol exhibited an ability to resist a second infection with heterologous virus comparable with that of untreated mice.
Conclusions Our results demonstrate that paracetamol dramatically reduces the morbidity associated with influenza but does not compromise the development of adaptive immune responses. Overall, these data support the utility of paracetamol for reducing the clinical symptoms associated with influenza virus infection.
- Influenza
- pulmonary
- inflammation
- paracetamol
- virus
- not applicable
- infection control
- innate immunity
- respiratory infection
- viral infection
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/2.0/ and http://creativecommons.org/licenses/by-nc/2.0/legalcode.
Statistics from Altmetric.com
- Influenza
- pulmonary
- inflammation
- paracetamol
- virus
- not applicable
- infection control
- innate immunity
- respiratory infection
- viral infection
Introduction
Influenza A viruses are responsible for seasonal epidemics accounting for up to 500 000 deaths per annum worldwide; however, the propensity of these viruses to acquire new genetic material from animal reservoirs generates novel pandemic strains with greater morbidity and mortality. The global impact of influenza A has been ever-present throughout the 20th century with the Spanish (1918), Asian (1957) and Hong Kong (1968) influenza pandemics. The 1918–1919 Spanish influenza pandemic was estimated to have infected one-third of the world's population and accounted for 50–100 million deaths.1 In early 2009 attention was drawn to the emergence of a new pandemic H1N1 influenza A strain that quickly spread to all continents.2 Therapeutic strategies for prevention and treatment of influenza A viruses remains a complex problem since animal reservoirs and antigenic shift can create potentially pandemic strains. This makes it difficult and costly to eliminate influenza A or create effective vaccines. Antiviral drugs are the first line of defence against new strains, but these are costly, resistance may occur and availability is limited.
Many factors contribute to the pathophysiology of influenza infection, including direct effects of both the virus and host factors such as the magnitude and type of immune response.3–6 Over-the-counter medicines that inhibit cyclo-oxygenase (COX) such as aspirin, paracetamol (acetaminophen) and other non-steroidal anti-inflammatory drugs (NSAIDs) are often taken to alleviate symptoms of influenza virus infection. Despite their ubiquitous availability and use, the effects of this group of drugs, including paracetamol, on influenza infection have been poorly investigated. Although widely used, the pharmacotherapeutic effects of paracetamol remain controversial, with some studies suggesting that the drug serves largely as an analgesic while others report additional anti-inflammatory effects (reviewed in Bertolini et al7). In support of an anti-inflammatory role, Hinz and colleagues demonstrated that paracetamol inhibits the activity of COX-2, an inducible enzyme that generates prostaglandins (PGs), prostacyclin and thromboxane.8 The role of this enzyme in influenza infection has been investigated further in COX-2-deficient mice which exhibit attenuated pulmonary inflammation and delayed viral clearance.9 Administration of paracetamol might therefore be expected to limit the severity of influenza-induced lung pathology, but this could adversely affect patients by impeding antiviral responses.10 Furthermore, perturbation of the inflammatory response could impair the development of protective immunity. Although influenza-specific immune responses have not been assessed in COX-2-deficient mice, Prymula et al have reported significantly lower antibody responses to some vaccines in infants receiving prophylactic paracetamol compared with those that did not.11
In view of the widespread availability of paracetamol and NSAIDs amongst the general population, we sought to examine the effects of paracetamol and a selective COX-2 inhibitor (celecoxib) in a mouse model of influenza virus infection. We hypothesised that administration of these drugs following infection would diminish the immune response, delay virus clearance and reduce immunopathology in target tissues. We further hypothesised that the adaptive immune response would be adversely affected, thereby compromising the establishment of protective immunity. In this study, each hypothesis was addressed by comparing innate and adaptive immune responses, virus clearance, lung pathology and the ability to resist challenge with heterologous influenza virus in both drug-treated and untreated mice.
Methods
Refer to the Supplementary information for a detailed description regarding the dose selection for celecoxib and paracetamol used during this study, flow cytometric analysis of pulmonary cells and lung intracellular cytokine staining, determination of serum alanine aminotransferase (ALT), quantification of pulmonary PGE2, in vivo lung function, in vitro virus titration assays and the statistical tests used.
Murine influenza infection model
Female C57BL/6 mice were sourced from Charles River UK and used at 8–10 weeks of age in experimental procedures in compliance with UK Home Office regulations.
Recombinant influenza A virus strains A-PR8-34 (PR8, H1N1) and A/X31 (X31, H3N2) were obtained from the National Institute for Medical Research (NIMR; London, UK). Mice were infected intranasally with 50 plaque-forming units (pfu) of PR8 in 50 μl of sterile phosphate-buffered saline (PBS), under light anaesthesia for primary influenza infection. Mice were injected daily intraperitioneally with either 10 mg/kg of celecoxib or 125 mg/kg paracetamol for 10 days as described in the Supplementary material. Memory recall responses to prior influenza infection were examined by rechallenging previously infected mice at 7–9 weeks after primary PR8 administration. Mice were infected intranasally with 200 pfu of X31 in 50 μl of sterile PBS under light anaesthesia. Body weight was recorded daily during the course of infection until mice were sacrificed or until day 16 for memory studies.
Mice were sacrificed at days 4, 7 and 10 during primary infection and day 4 after rechallenge, and bronchoalevolar lavage (BAL) was performed. Following BAL, lungs were isolated and kept on ice. A detailed description regarding BAL and lung cell isolation is provided in the Supplemental information.
Histopathology
At day 7 postinfection lungs were perfused with 5 ml of PBS and fixed with 10% neutral buffered formalin solution. Following fixation, lungs were embedded in paraffin and 5 μm sections were cut. Sections were stained with H&E and scored blinded. Lungs were scored using modified scoring criteria from Longhi et al12 and Ashcroft et al.13 The degree of leucocyte infiltration of the airway spaces and the formation of lymphoid aggregates were independently scored on an increasing scale (0–3). These were combined with the extent of perivascular lymphocytic aggregation within each section (0–10), to give a score ranging between 0 and 13. The mean score for each clinical parameter was statistically compared between groups.
Determination of PR8-neutralising antibody titres
Serum was isolated from mice at 7–9 weeks postinfection with PR8 and from naive uninfected mice. Madin–Darby canine kidney (MDCK) cells at 50% confluence were plated into flat-bottomed 96-well plates and cultured for 48 h in the presence of 720 pfu of PR8 and serum at a 1:8 dilution. Cell viability was calculated using alamar blue (Biosource International, Camarillo, California, USA) according to the manufacturer's guidelines. The efficacy of serum from influenza-primed mice to neutralise PR8 was compared with the neutralising efficacy of serum from influenza-naive mice.
Results
Celecoxib and paracetamol diminish influenza-associated pulmonary PGE2 and reduce the extent of influenza-induced airway inflammation
The anti-inflammatory effects of NSAIDs such as celecoxib are attributed mainly to their ability to inhibit the activity of COX-2, an enzyme which catalyses the biosynthesis of PGs from arachidonic acid. COX-2 is upregulated during an inflammatory response, thereby increasing levels of PGs, most notably PGE2, a metabolite known to modulate immune cell activity. To determine whether PGE2 in the lungs of influenza-infected mice is COX-2 derived, lipids from lungs of infected mice and infected mice administered either celecoxib or paracetamol were analysed using liquid chromatography/tandem mass spectometry. Our data indicate that PGE2 is elevated in the lungs of influenza-infected animals compared to uninfected controls (763 +/− 75ng/mg vs 408 +/− 128), and that this is inhibited by administration of either celecoxib (376±122 ng/mg) or paracetamol (368±111 ng/mg) (data not shown). These observations indicate that elevated PGE2 following influenza infection is COX-2 derived. A similar extent of PGE2 inhibition was observed with celecoxib and paracetamol, in line with reports of their comparable ability to inhibit PGE2 production in human blood.8
Infection of mice with influenza virus typically results in occlusion of the lung airway spaces by inflammatory cell infiltration. To gain insight into the inflammatory response in the airways, a quantitative analysis of the cellular composition of BAL fluid of infected mice was performed by flow cytometry. As expected, there was a highly significant increase in the numbers of cells recovered from BAL of infected mice compared with uninfected animals (figure 1A–F). Paracetamol administration significantly reduced cell infiltration at days 4, 7 and 10 postinfection. Detailed analysis showed significantly fewer neutrophils, inflammatory monocytes, eosinophils and natural killer (NK) cells in the BAL of paracetamol-treated animals compared with untreated mice (figure 1C–F). A similar pattern was observed in the celecoxib-treated group where a trend for fewer cells was observed at each time point, although in this case a statistically significant difference was observed only at day 10. Overall, these data imply a protective effect of paracetamol in reducing the extent of influenza-induced immunopathology.

Effect of celecoxib or paracetamol upon influenza-induced airway inflammation and immunopathology. Bronchoalveolar lavage (BAL) fluid was aspirated at day 4, 7 or 10 postinfection and the airway-infiltrating cells were isolated. The total numbers of cells (A) and the numbers of alveolar macrophages (B), neutrophils (C), inflammatory monocytes (D), eosinophils (E) and natural killer (NK) cells (F) at the selected time points were determined by flow cytometry. (G) At day 7 postinfection, airway hyper-responsiveness to inhaled methacholine (10 mg/ml for 5 min) was determined by measuring changes in Penh (enhanced pause) 5 min after methacholine challenge, expressed as a percentage of the premethacholine baseline. Changes in Penh were measured from naive mice, used as an uninfected control, and virus-infected mice treated or not with celecoxib or paracetamol. Results are expressed as the mean±SEM for each group, n=6 mice/group. The significance of differences between drug-treated and untreated mice was assessed using one-way analysis of variance with Tukey–Kramers post hoc multicomparison test (A–F) and between infected and uninfected mice using an unpaired t test (G), *p<0.05; **p<0.01; ***p<0.001.
Airway function is improved in mice receiving celecoxib and paracetamol
The observation that inflammation is reduced by paracetamol administration is further supported by an analysis of airway function in the same groups of mice. A group of influenza-infected, untreated control animals and animals treated with either paracetamol or celecoxib were subjected to airway function tests performed by whole-body plethysmography. This technique measures changes in air pressure as a result of inspiration and expiration. In these experiments, enhanced pause (Penh), a measurement of changes in expiratory effort, was used as an indicator of airway obstruction. Specifically, control and drug-treated animals infected 7 days previously with influenza virus were challenged with aerosolised methacholine (10 mg/ml) and Penh was measured. There was a significant increase in the peak Penh response in all infected mice compared with uninfected controls; however, this difference was less pronounced in celecoxib- and particularly paracetamol-treated mice (figure 1G).
To assess if this improvement in lung function corresponded to a reduction in influenza-induced pulmonary immunopathology, lungs, recovered at day 7 postinfection, were assessed histologically. Extensive leukocyte infiltration (figure 2A,C) and perivascular aggregation (figure 2B,D) were observed in all influenza-infected animals, when compared with uninfected control animals. These observations, while not significantly different between drug-treated and untreated mice, when combined with a notable improvement in lung function in paracetamol-treated mice, imply a marked reduction in influenza-associated pathology during the acute stages of infection.
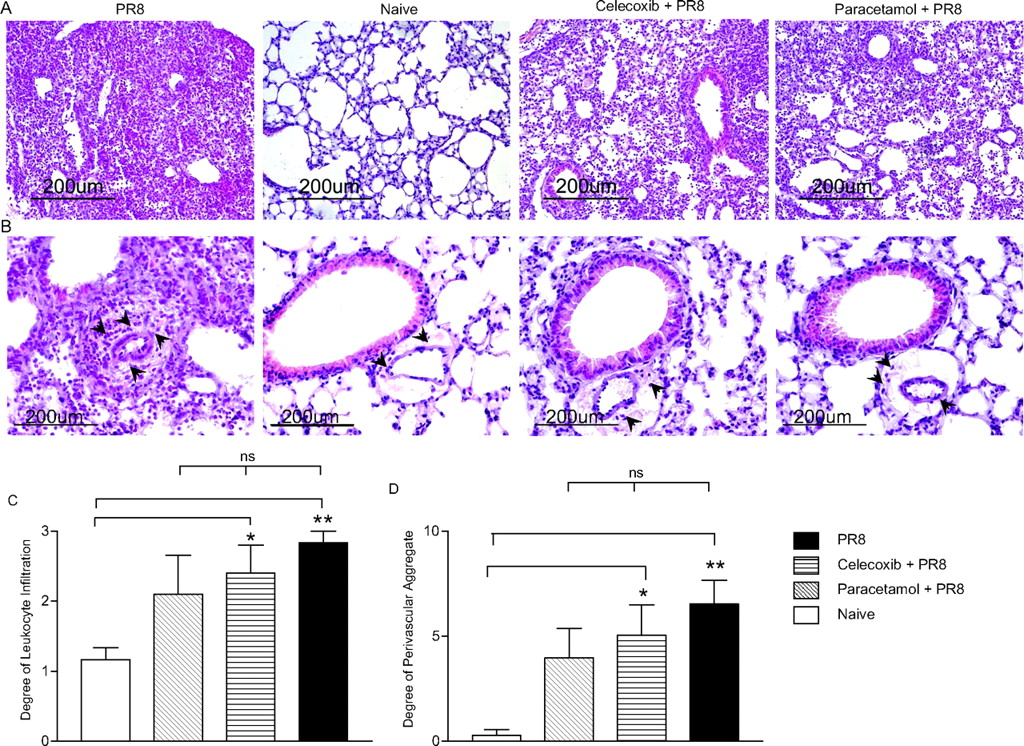
Histological analysis of pulmonary sections from treated and untreated mice postinfection with influenza virus. Lungs were harvested from mice at day 7 postinfection and, following perfusion, were formalin fixed, embedded in paraffin wax, sectioned and stained with H&E. (A and B) Representative sections depicting pulmonary leukocytes from influenza-infected, treated and untreated mice were compared with uninfected mice (original magnification×10 (A)) Arrowheads highlight areas of lymphocytic perivascular aggregate (original magnification×20 (B)). The mean score±SEM for each group is given, n=6 mice/group for the severity of influenza infiltrate (C) and degree of perivascular lymphoid aggregate (D). The significance of differences between influenza-infected mice and uninfected mice was determined using one-way analysis of variance with Tukey–Kramers multicomparison test, *p<0.05; **p<0.01.
The effects of celecoxib or paracetamol on influenza-induced liver dysfunction
Influenza infection of humans can be associated with an increase in serum ALT levels and a transient and self-limiting hepatitis.14 Studies of mouse models of influenza infection indicate that liver injury during influenza infection is not attributable to influenza infection of the liver but rather represents collateral damage caused by antigen-independent sequestration of immune cells, most notably influenza-specific CD8+ T cells in the liver. We sought to determine whether ALT levels were increased in the serum of influenza-infected mice at day 7 postinfection and whether this was reduced by administration of paracetamol or celecoxib. The results, shown in figure 3A, indicate that both drugs virtually abolished influenza-induced increases in ALT levels. Weight loss is also commonly used as an indicator of the severity of influenza-induced pathology. However, we observed no change in weight loss following influenza infection as a consequence of celecoxib or paracetamol administration (figure 3B and Supplementary figure 1).
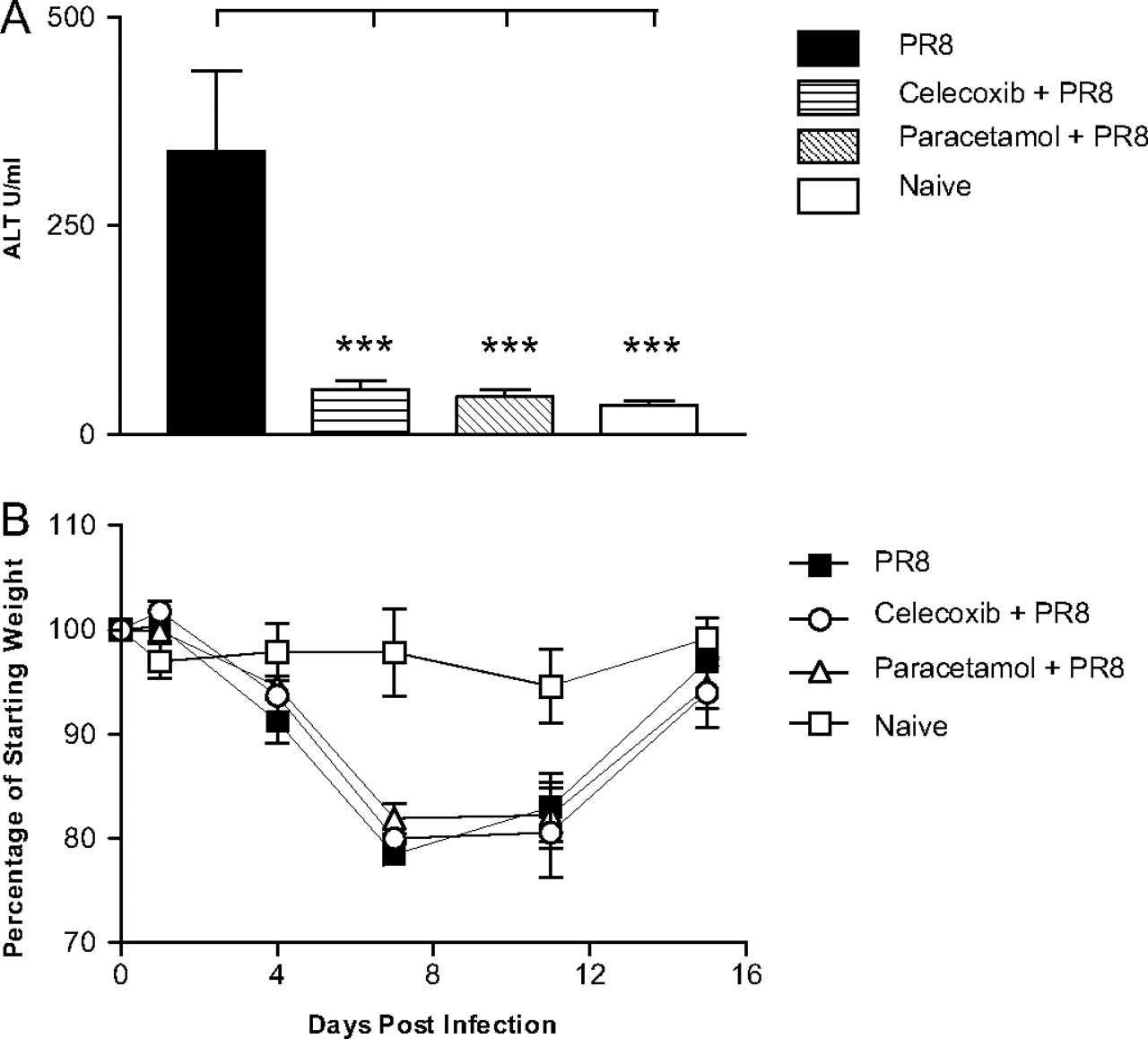
Effect of cyclo-oxygenase 2 (COX-2) inhibition upon clinical parameters of primary influenza infection. (A) Serum collected from mice at day 7 postinfection was used to determine circulating alanine aminotransferase (ALT) levels during acute influenza infection. Results are expressed as the mean±SEM, n=6–16 mice/group. (B) Post-PR8 infection mice were weighed daily for 16 days, and changes in body weight were expressed as a percentage of their starting weight. Each data point represents the mean±SEM, n=6 mice/group. The significance of differences between drug-treated and untreated mice was assessed using one-way analysis of variance with post hoc Tukey–Kramer multiple mean comparison,*p<0.05; **p<0.01; ***p<0.001.
Virus clearance is unimpaired in celecoxib- or paracetamol-treated mice
As paracetamol and, to a lesser extent, celecoxib reduces the magnitude of the inflammatory response to influenza virus, we hypothesised that viral clearance would be delayed in these animals compared with non-treated controls. Viral titres in the lungs of influenza-infected mice declined on days 7 and 10 compared with day 4 postinfection. Surprisingly we observed no significant differences in viral titres among the groups at any of the time points analysed (figure 4).

Pulmonary viral load during acute influenza infection. At days 4, 7 and 10 postinfection mice were sacrificed and the lungs were extracted and homogenised. Pulmonary viral titres were determined using the previously described MDCK (Madin–Darby canine kidney) virus quantitation assay; results are given as the mean for each group±SEM, n=4–9 mice/group.
Protective immunity is not impaired in mice treated with paracetamol or celecoxib during primary infection with influenza virus
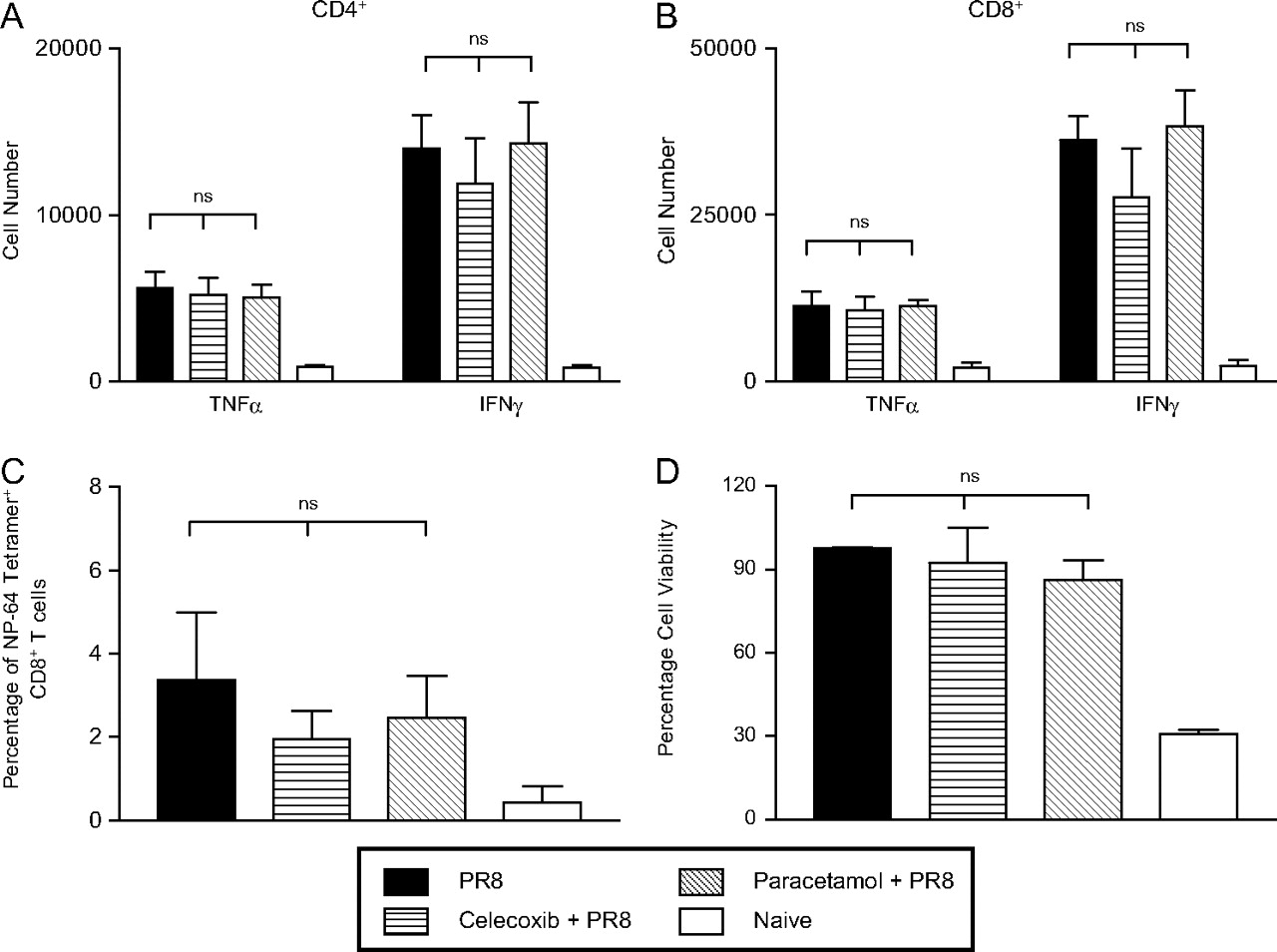
A reduction in influenza-induced innate immune responses as a consequence of celecoxib or paracetamol treatment could alter the magnitude of the virus-specific adaptive immunity. To investigate this, interferon γ (IFNγ)- and tumour necrosis factor α (TNFα)-producing CD4+ and CD8+ T cells in the lungs were measured at day 10 postinfection. The number of cytokine-producing cells was, as expected, elevated as a result of the infection, but there was no statistically significant difference between influenza-infected groups treated or not with drugs (figures 5A,B and Supplementary figure 2). As an additional measure of the influenza-specific CD8+ T cell response, nucleoprotein-64 (NP-64) tetramers were used. The results shown in figure 5C indicate that the administration of either celecoxib or paracetamol did not significantly alter the induction of virus-specific CD8+ T cell responses.

Adaptive immune responses following acute influenza infection and inhibition of cyclo-oxygenase 2 (COX-2). (A–C) Lungs were harvested at day 10 postinfection and a single cellular suspension was achieved following collagenase digestion. Lung cells were stimulated for 5 h in vitro with PMA (phorbol 12-myristate 13-acetate) and ionomycin; the numbers of CD4+ (A) and CD8+ T cells (B) secreting tumour necrosis factor α (TNFα) and interferon γ (IFNγ) were determined by flow cytometry. (C) Alternatively, lung cells were not stimulated and the percentage of CD8+ influenza-specific tetramer+ cells was determined by flow cytometry. (D) Serum from both treated and untreated mice was isolated at 7–9 weeks postinfection with 50 plaque-forming units of PR8. The relative neutralising antibody activity of each group of mice was determined using the neutralising antibody titre assay described in the Methods section. Each data point given is the mean±SEM, n=3–6 mice/group.
Finally, the virus-neutralising capacity of serum recovered from treated and untreated mice was assessed and, again, no significant difference was observed between mice in the different treatment groups (figure 5D). As expected, the neutralising ability of serum from all influenza-infected memory mice was significantly greater than that from naive uninfected mice. Overall these data indicate that while administration of paracetamol or celecoxib reduces the inflammatory response induced by influenza virus infection, there is no effect on the adaptive immune response.
As the adaptive immune response was unimpaired in mice treated with COX-2 inhibitors, we considered it likely that there would be no loss of protective immunity in these mice. To determine this, mice infected with the PR8 virus at least 8 weeks previously were rechallenged with the heterologous influenza virus, X-31, enabling assessment of hetero-subtypic immunity. In this case, T cells would mediate protective immunity given that neutralising antibodies raised following PR8 infection do not cross-neutralise the X-31 virus. As expected, prior infection with the PR8 virus did confer some protection against infection with X-31 as these animals displayed significantly less weight loss than animals that had not experienced a prior exposure to influenza (figure 6A). In agreement with this, viral titres were significantly reduced in PR8-immune mice, including those treated with either paracetamol or celecoxib during the primary infection, compared with mice exposed to influenza virus for the first time (figure 6B). These findings indicate that administration of paracetamol or celecoxib does not compromise the development of protective T cell-mediated immunity.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Protective immune responses to secondary influenza infection. (A and B) At 7–9 weeks postinfection mice were rechallenged with 200 plaque-forning units (pfu) of X31. (A) Mice were weighed prior to secondary infection to provide a baseline 100% starting weight and for 4 days thereafter. Each data point represents the mean±SEM for each group, n=3–6 mice/group. (B) At day 4 after secondary infection, mice were sacrificed and the lungs were harvested following perfusion with phosphate-buffered saline. Lungs were homogenised and the viral load was determined using the previously described MDCK (Madin–Darby canine kidney) plaque assay. Plots represent the mean of each group±SEM, n=6 mice/group, 3 mice/group for PR8-naive mice. The significance of differences between previously PR8-infected mice and PR8-naive mice was evaluated using one-way analysis of variance with Tukey–Kramer multiple comparison post hoc test, ***p<0.001.
Discussion
The current study set out to examine the impact of paracetamol on the course of influenza virus infection, focusing on viral clearance, the generation of innate and adaptive immune responses and the establishment of protective immunity. The data clearly demonstrate that in the context of the influenza-infected lung, paracetamol reduces pulmonary PGE2 and inflammation without compromising virus clearance or the generation of protective immunity.
During the early stages of virus infection (days 4–10) we found that paracetamol reduced the magnitude of infiltration of innate cells into the lungs. A similar trend, although to a lesser extent, was observed in those treated with the selective COX-2 inhibitor celecoxib. These results are similar to findings using COX-2-deficient mice.9 The effect of influenza infection was less severe in COX-2-deficient animals, as demonstrated by decreased mortality and a blunted inflammatory response compared with wild-type animals. However, elevated virus titres were observed in the animals, indicating, in contrast to our own findings, that the abrogated inflammatory response was associated with poorer control of virus. The more pronounced effects observed in COX-2-deficient animals compared with those treated with COX-2 inhibitors may reflect developmental differences in the genetically modified animals affecting the function of the immune system and/or the difference between a complete (COX-2−/− animals) and transient (drug administration) block of COX activity.
Influenza infection caused airway hyper-responsiveness to the inhaled spasmogen, methacholine, in agreement with previous reports,15 which was reduced following treatment with paracetamol or celecoxib. Thus, paracetamol appears to play a protective role, reducing lung pathology and helping to preserve lung function. Patients infected with seasonal influenza A, who subsequently develop severe respiratory failure, are characterised by a profound early inflammatory infiltrate and cytokine release. The same was true for the 2009 novel H1N1 influenza A pandemic where severe lung injury occurred in young adults in whom it was frequently difficult to maintain oxygenation. Autopsies on patients succumbing to this new influenza strain showed pulmonary pathology similar to previous pandemics, with profound cellular infiltrates and diffuse alveolar damage.16–18 Thus paracetamol may, through inhibition of COX-2, afford protection for patients infected with influenza and complicated by respiratory failure through attenuation of the proinflammatory response. The importance of the COX-2 pathway in severe respiratory failure is further substantiated by experiments using avian H5N1 influenza A virus.19 Avian H5N1 infection has a case death approaching 30% but is not transmissible between humans; it is characterised by severe respiratory failure with inflammatory infiltrates within the lung and high levels of inflammatory cytokines such as TNFα and IFNγ.20 21 Compared with seasonal H1N1 influenza A virus, avian H5N1 differentially upregulates COX-2 in primary human macrophages in vitro and epithelial cells in vivo, the increased expression resulting from autocrine or paracrine mediators rather than direct effects of the virus.19 Expression of inflammatory cytokines is driven by COX-2 upregulation and can be effectively inhibited with selective COX-2 inhibitors.19 In another murine model of avian H5N1 infection, addition of the selective COX-2 inhibitor celecoxib to the neuraminidase inhibitor zanamivir reduced lung inflammation and virus titres and increased survival from 13.3% to 53.3% and mean survival time from 8.4 to 13.3 days compared with zanamivir alone.22 It is clear from this study and our own that attenuating the inflammatory response to influenza does not compromise virus clearance. Indeed, overall we observed a trend for better virus control in the treated animals.
Overall these data support a protective role for paracetamol during the acute stage of infection with influenza virus. We considered it important that the impact of paracetamol administration at this time on the generation of influenza-specific responses was also assessed. As the innate immune response influences the development of the adaptive response, attenuation of this response through the effects of paracetamol could suppress the virus-specific immune response. In addition, PGE2 has been shown in vivo to promote T helper 1 (Th1) cell differentiation, thus inhibition of PGE2 could directly downmodulate development of influenza-specific T cell and antibody responses.23 In support of this, a recent report detailing antibody responses in 226 infants receiving vaccines demonstrated significantly lower antibody titres in those receiving prophylactic paracetamol.11 Our data clearly show, however, no significant differences in the influenza-specific immune response in drug-treated and untreated animals. Importantly, the ability of drug-treated mice to resist rechallenge with a heterotypic influenza virus was unimpaired.
The current study demonstrates that both paracetamol and celecoxib, which inhibit COX-2, attenuate pulmonary inflammation without impeding virus clearance or the development of specific immunity in mice. While the doses of paracetamol used are different for mice and humans, as described in the Supplementary material, the implications of the study are clear. Over-the-counter medicines such as paracetamol, which inhibit COX-2, are cheap, widely available and target the host response rather than the virus, and therefore offer an attractive therapeutic option.24 However, with the information currently available on the effects of paracetamol upon influenza infection, it is possible that its impact may differ in mice and humans. It is important that these observations, generated using a mouse model, are validated with respect to the action of paracetamol and its impact on the immune system in clinical studies of human influenza virus infection.
Acknowledgments
The authors would like to thank Dr Emma Jones for her expertise with the histological images in figure 2, Dr Sivasankar Baalasubramanian for assistance in performing the experiments in figure 1, Dr Mike Badminton for facilitating the measurements of alanine transaminase, and Mr Lloyd Morgan for providing assistance in the generation of preliminary data for the study.
References
Supplementary materials
Web Only Data thx.2010.150318
Files in this Data Supplement:
Web Only Data thx.2010.150318
Files in this Data Supplement:
Footnotes
VO and AMG contributed equally to this work.
Funding This work was supported by a Wellcome Trust project grant (080340), an MRC Senior Non-Clinical Fellowship awarded to AMG (G117/488) and PRT (G0601617), and a European Union Marie Curie Fellowship awarded to SRC.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Airwaves