
Abstract
Impaired lung development afflicts a range of newborns cared for by paediatric surgeons. As a result the speciality has led in the development of surgical models that illustrate the biomechanical regulation of lung growth. Using transgenic mutants, biologists have similarly discovered much about the biochemical regulation of prenatal lung growth. Airway smooth muscle (ASM) and its prenatal contractility airway peristalsis (AP) represent a novel link between these areas: ASM progenitors produce an essential biochemical factor for lung morphogenesis, whilst calcium-driven biomechanical ASM activity appears to regulate the same. In this invited paper, I take the opportunity both to review our recent findings on lung growth and prenatal ASM, and also to discuss mechanisms by which ASM contractility can regulate growth. Finally, I will introduce some novel ideas for exploration: ASM contractility could help to schedule parturition (pulmonary parturition clock) and could even be a generic model for smooth muscle regulation of morphogenesis in similar organs.
Similar content being viewed by others


Why lung development matters to paediatric surgeons
Lung development is important to paediatric surgeons. Lung hypoplasia and its lethal sequlae afflict a spectrum of our patients ranging from those with congenital diaphragmatic hernia (CDH) or prenatal uropathies to massive abdominal wall defects [1–4]. Similarly, congenital cystadenomatoid lung malformations and bronchopulmonary sequestrations present to paediatric surgeons as antenatally diagnosed masses or postnatally with symptomatic chest lesions [5, 6]. Moreover, several of our patients are born prematurely with incomplete development that predisposes to lung disease of prematurity [7]. Finally many of our patients with CDH and tracheo-oesophageal fistula/oesophageal atresia have chronic respiratory morbidity that affects growth and may affect decisions for further surgeries (such as fundoplication) [8, 9] (Fig. 1).

Problems of lung development and function are not confined to CDH: a Contrast swallow (lateral view) after delayed primary repair of oesophageal atresia (OA) without distal fistula. Such children may face major respiratory problems (tracheomalacia and aspiration +/− reflux) that can present as, e.g. life-threatening episodes or later with reactive airways disease; b Giant gastroschisis with major liver herniation: such newborns face significant respiratory difficulties requiring, in this case, long-term ventilation after eventual abdominal closure. Both images used with parental consent and courtesy of the author
Given this wide impact of lung development and disease on paediatric surgical practice, it is perhaps no surprise that neonatal surgeons have been at the forefront of investigation: the most notable contribution has been the creation of in vivo fetal models of lung maldevelopment that have allowed us to understand the regulation of pulmonary growth and to design strategies to promote it prenatally (e.g. surgically created models of CDH and obstructive uropathies) [10, 11] (Fig. 2). Moreover, paediatric surgeons have taken a leading role in development of extracorporeal membrane oxygenation as a rescue therapy for refractory respiratory failure and have collaborated in the evaluation and adoption of perinatology approaches such as antenatal glucocorticoids, inhaled nitric oxide, permissive hypercapnoea and sildenafil for surgical patients [12–17].

Surgical models elucidate the biomechanical regulation of lung growth: the gravid ovine uterus has been delivered at laparotomy. Sonography is being used to determine placental and fetal positions prior to hysterotomy. These methods of manipulating the ovine fetus have allowed several models to be created (CDH, obstructive uropathy, gastroschisis and myelomeningocele). Of course, forms of lung hypoplasia generated this way may differ importantly from both teratogenic/transgenic models and indeed humans. Image courtesy of the author
An overview of lung development reveals points of clinical importance
Anatomically the mammalian lung develops as a diverticulum from the primitive foregut [18]. Through iterated epithelial branching (branching morphogenesis), the airways take form [18, 19]. Early direct vascular supply from the aorta to the lung normally dwindles: abnormal persistence is observed in sequestrations [20, 21]. Pulmonary vascular development was previously held to follow airway morphogenesis. However recent studies indicate that vascularisation may be an important promoter of airway growth and terminal differentiation [22, 23]. Temporally, the pulmonary diverticulum emerges at around 4 weeks of gestation; failure of the diverticulum to separate normally is thought to explain tracheo-oesophageal fistulae [18, 24]; most airway branching occurs between 6 and 16 weeks [25, 26]; vascular connectivity to establish the respiratory surface for gas exchange emerges around 22 weeks onward (hence the putative limit to survival of preterm birth) [27]; human alveolarisation is in progress by birth and commonly reported to continue through early childhood [25]. Physiologically, the lung undergoes dramatic changes between fetal and neonatal life. Prenatally, the lung is fluid filled and respiratory gas exchange is undertaken instead via the placenta (providing the rationale for the EXIT procedure for, e.g. congenital airway obstructions) [28, 29]. Primitive endodermal cells differentiate into several lineages of which the most studied are the types 1 and 2 pneumocyte and which may also include pulmonary stem cells [30, 31]. Pulmonary epithelium is responsible for lung liquid production (via chloride-secretion): this results in prenatal lung having a small positive intraluminal pressure relative to the thoracic cavity [32]. Abnormal persistence of this lung fluid after birth is held responsible for transient tachypnoea of the newborn [33]. Surfactant production by type 2 pneumocytes is a further adaptation that assists in lung inflation at birth and resists airspace collapse. Surfactant deficiency is associated with respiratory distress syndrome in premature newborns and may occur in other disorders such as CDH [34, 35]. Estimation of lung maturity has been attempted by assaying surfactant-related metabolites in amniotic fluid [36–38]. Whilst the epithelium has absorbed much scientific interest for conferring tissue specialisation (liquid production, surfactant manufacture), the pulmonary mesenchyme is essential to lung development [39]. Stripping embryonic lung buds of mesenchyme halts their branching; replacing tracheal mesenchyme with that from end-buds restarts epithelial branching at the trachea [40]. Moreover, murine mutants lacking specific mesenchymal gene expression exhibit both lung lesions and diaphragmatic defects with some phenotypic similarities to human CDH [41–43]. These observations illustrate the importance of pulmonary mesenchyme and also support the concept that an early mesenchymal lesion in CDH can generate both lung hypoplasia and diaphragmatic defect [44–46] (Fig. 3). As we shall see, the latest evidence suggests that pulmonary mesenchyme and its derivatives have both biochemical and biomechanical roles in the regulation of lung development.

Non-surgical models of abnormal lung growth are also valuable: teratogenic and transgenic models illustrate that some clinical problems of lung function may have their roots much earlier in development than appreciated from surgical models alone. This shows photomicrographs of whole lung primordia in organ culture. Shown on the left of the figure is the longitudinal development of a normal lung rudiment. For comparison the development of a nitrofen-exposed lung rudiment is shown on the right at the same timepoints. All specimens are at the same magnification (scale bar 400μ m) and the trachea lies superiorly in each case. Nitrofen-exposed lung primordia are growth-impaired prior to the visceral hernia supervening. Image courtesy of the author
Understanding of lung development derives from a variety of sources
Our understanding of how the lung develops has been aided by comparative biology, transgenic and teratogenic mutants, surgically created models of in vivo physiology and human ‘experiments of nature’. Comparative biology has enabled us to appreciate that much of lung development (epithelial branching morphogenesis, vasculogenesis and angiogenesis) is conserved through evolution [47, 48]. This in turn has enabled the similarities with comparable processes in other mammalian organs (pancreas, salivary gland and kidney) to be exploited [49, 50]. Transgenic mutant gene knockout animals have been particularly helpful in establishing the biochemical regulation of airway development by demonstrating the necessary roles of certain growth and transcription factors [51, 52]. In contrast, surgical models of modified lung development have permitted the biomechanical regulation of lung development to be elucidated [10, 11, 53]. Furthermore, human cases have provided vivid illustrations of the biomechanical regulators of lung growth (congenital laryngeal atresia, Potter’s syndrome) [54, 55]. Despite the wealth of knowledge generated by these separate approaches, a unifying model that brings together biochemical and biomechanical regulation of lung growth is still being sought [56–58]. We have been addressing this by studying potential links between biomechanical and biochemical stimuli; in so doing we and others are appreciating new roles for pulmonary mesenchyme in lung organogenesis.
The biochemical regulation of lung development
A key biochemical interaction that underpins lung development is that of fibroblast growth factor-10 (FGF10) and its cognate receptor FGFR2IIIb. Both ligand and receptor have been conserved through evolution and are homologous with the branchless and breathless genes that are essential for airway branching in Drosophila [48]. In a parallel manner, both FGF10 and FGFR2IIIb have been shown to be necessary for lung development in transgenic knockout mice [59–62]. Functional loss of either leads to lung agenesis in the offspring. In vitro lung culture studies have shown that FGF10 induces epithelial branching and can in fact largely substitute in this regard for pulmonary mesenchyme: FGF10 stimulates branching of epithelial only cultures in Matrigel [63]. Abnormalities of FGFs and related heparan sulphate signalling have been demonstrated in experimental lung hypoplasia [64–68]. Several other biochemical regulators of lung morphogenesis are arrayed around the FGF10–FGFR2IIIb interaction (and extensively reviewed elsewhere) [51, 52]. However for the purposes of this review, a focus on this central pathway is sufficient. As we shall see, this archetypal biochemical effector of lung growth may in fact also have a role to play in the biomechanical regulation that we will now survey.
The biomechanical regulation of lung development
Fetal breathing movements, lung liquid production and maintenance of fetal thoracic and amniotic volumes are necessary for normal lung development [55]. Ablation of fetal breathing movements either by phrenic nerve ablation or generation of amyogenic transgenic murine mutants is associated with forms of lung hypoplasia [69, 70]. Excessive lung liquid loss induced by fetal tracheostomy similarly impairs lung growth [71]. Compromise of thoracic volume by visceral herniation in CDH models or due to external fetal compression from oligohydramnios in prenatal uropathies are both associated with forms of lung hypoplasia [11, 72]. Finally, and more difficult to explain, massive abdominal wall defects with extensive liver herniation are also associated with clinically significant lung impairment and often a narrowed thorax [4]. Speculating, it may be that gross reduction in abdominal domain results in commensurate limitation of thoracic growth. Others have suggested that liver herniation draws the diaphragm down leading to major airways becoming kinked across the pulmonary vessels with resulting bronchomalacia [73]. Finally, it is difficult to separate out the deleterious effects of barotrauma resulting from the prolonged ventilation that such babies often require. Having considered the substantial effects that biomechanical stimuli can have on lung development, we can now turn to the focus of our recent work on a biomechanical pulmonary phenomenon that has been long described but largely overlooked.
Airway smooth muscle: ‘appendix of the lung’ or regulator of growth?
From the earliest stages of lung development and throughout subsequent prenatal life the developing lung is itself vigorously mechanically active [74–77]. Peristaltic waves of airway smooth muscle (ASM) rhythmically propagate throughout the airway tree with resulting propulsion of all important lung liquid (Fig. 4). As a result the growing end buds of the lung (where ASM elaboration is at a minimum) experience phasic distension and relaxation [74, 75]. This curious activity has been observed in airway explants from avian, rodent and human species and they appear to increase in frequency as gestation progresses [74–77]. Why the developing lung should expend so much effort in the synthesis of contractile apparatus and its mechanical activity has remained a mystery. Certainly, rhythmic stretch-relaxation modulates growth and gene expression in a range of cell types including pulmonary epithelium and appears quite distinct from the effects of static stretch [78, 79]. However, why this vibrant peristaltic contractility should suddenly disappear at birth and how it relates to postnatal ASM function and dysfunction are also unknown. Given these uncertainties, we have explored prenatal ASM activity, its regulation and its potential purposes. Most fascinating has been emerging evidence that in ASM peristalsis, we may have an important bridge between biomechanical and biochemical stimuli to lung growth. Finally, the insights gained from our studies hold the promise that strategies to treat prenatal lung hypoplasia and postnatal reactive airway disease can be modified to achieve greater efficacy.

Airway peristalsis—a novel regulator of lung growth? Left hand panels: fluid flux within the epithelial lumen (dark) of cultured lung can be visualised due to the peristaltic propulsion of cellular debris (leading edge arrowed) seen here on rapid sequence photography of whole lung. Right hand panels: airway contraction (boxed) during a wave of airway peristalsis. A video of airway peristalsis from our work is viewable on the web. http://ajrcmb.atsjournals.org/content/vol32/issue2/images/data/118/DC1/Jesudason_Supplement_001.avi
Prenatal ASM peristalsis appears coupled to lung growth
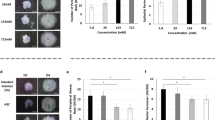
We began with the seemingly simple task of determining where and when in development ASM peristalsis emerges. Our data showed that even embryonic rodent explants equivalent to roughly 5 weeks of human gestation would exhibit powerful peristaltic contractility in vitro [75]. Contrary to expectation, these waves did not arise simply from the trachea and propagate distally. In fact it appears that ASM contraction can arise throughout the proximal airways at this stage but with a clear predilection for the proximal right lung (‘pulmonary pacemaker“) [75]. Moreover this area of the developing lung seems to dominate and be autonomous of the remainder of the lung: isolation of other areas of the lung from the influence of the right led to more frequent contractility in the isolated segment; in contrast, isolation of the right lung did not alter peristaltic frequency therein [75]. To ascertain what was driving this activity, we used calcium imaging and c-kit immunoreactivity to show that ASM peristalsis was underpinned by spontaneous propagating calcium waves that arise in the apparent absence of Cajal-type pacemaker cells [75, 80] (Fig. 5a). To explore the purpose of such contractility we alternately stimulated or inhibited either peristalsis or lung growth. In normal embryonic lung we discovered that growth and peristalsis were coupled: modulation of one was reliably accompanied by parallel alteration in the other [75]. In particular, we had an indication of a link between a typical biochemical regulator of lung growth (FGF10) and a biomechanical one (ASM peristalsis): FGF10-stimulated lung growth was associated with significant increase in peristalsis frequency [75]. Hence, these studies showed a degree of coupling between ASM peristalsis and FGF10-driven lung growth and supported the concept that one may regulate the other. Furthermore, given that ASM peristalsis depends on propagating calcium waves, these data indicate that such calcium waves may in turn regulate lung growth [80].

Ca2+ waves underpin airway peristalsis and hence potentially growth: a Stacks of pseudo-colour confocal images of Fluo-4–loaded embryonic airways showing propagating Ca2+ waves with the initiation site (marked by an asterisk) in the trachea (top row) and right lung rudiment (bottom row); b blockade of sarcoplasmic calcium reuptake with cyclopiazonic acid (CPA) leads to raised intracellular Ca2+ (y-axis) and rapid abolition of Ca2+ oscillations over time (x-axis). As described in the text, this is associated with replacement of peristalsis by sustained airway contraction, halting of branching and loss of SMA expression (not shown); c superimposed ASM Ca2+ transients measured from normal and nitrofen lung (as labelled) showing altered morphology in lung hypoplasia
Ca2+: biochemical mechanism for ASM regulation of lung development?
In preliminary studies to explore, how calcium may regulate lung growth we have identified some interesting phenomena. L-type calcium channel blockade with nifedipine reduces intracellular calcium, halts airway peristalsis (AP) (inducing a flaccid paresis) and stunts lung size but without significantly diminishing branching [75, 81]. In contrast, blocking sarcoplasmic calcium reuptake with cyclopiazonic acid raises intracellular calcium leading to a tetanic paresis (and halt of airway contractility) and is associated with a remarkable and complete abolition of branching morphogenesis [82, 83] (Fig. 5b). Moreover this latter effect is associated with loss of smooth muscle actin expression (SMA: an early marker of ASM lineage) [83]. Removal of cyclopiazonic acid leads to resumption of apparently normal branching and SMA expression [83]. Given that ASM progenitors specifically furnish the FGF10 required for airway branching [84], we therefore hypothesise that cyclopiazonic acid may be acting on ASM progenitors to halt both commitment (loss of SMA expression) and also FGF10 production (loss of branching). Supporting the idea that sarcoplasmic calcium regulates both FGF10-driven branching morphogenesis and ASM elaboration, one of the first described human genopathies relating to loss of sarcoplasmic calcium reuptake is associated with dysregulated proliferation an epithelium that, like the airway, is FGFR2IIIb-bearing and FGF10-responsive [85].
A biomechanical mechanism for ASM regulation of lung development?
We have also argued that coupled elaboration of FGF10 and ASM could operate as a biological feedback mechanism to maintain intraluminal pressures in the growing lung (loss of such pressure is associated with lung hypoplasia) [46]. As the lung lumen increases in volume due to FGF10 mediated growth, maintenance of intraluminal pressure would require increased lung liquid production and/or increased wall tension. The latter (and perhaps even the former—vide infra) could be supplied by increased ASM elaboration and activity: as more ASM progenitors produce FGF10 and growth, more differentiated ASM develops to influence airway wall tension and maintain luminal pressure. Moreover, early work suggests that ASM peristalsis is associated not only with propagating calcium waves but potentially with chloride efflux from ASM. We hypothesise that this chloride could be taken up at the adjacent basolateral epithelial surface to drive epithelial chloride-secretion led production of lung liquid. Hence ASM peristalsis may regulate growth-related intraluminal pressures not only via its influence on wall tension but also through modulation of all important lung liquid production.
What then are the implications for abnormal lung development? Lung hypoplasia is associated with abnormalities of prenatal ASM-related signalling and postnatal function in human CDH and in experimental CDH lung [8, 86, 87]. We have now shown that the coupling between peristalsis and growth, the underpinning calcium waves and the stereotypical maturation of peristaltic activity are all abnormal in teratogen-induced lung hypoplasia [88, 89] (Fig. 5c). Prenatal ASM may also play key roles in other heterogenous forms of experimental lung hypoplasia: myogenic knockout mice where intercostal musculature +/− diaphragmatic muscularisation are ablated appear to have normal ASM elaboration up to and beyond the point at which fetal breathing supervenes [70]. In only those mutants lacking both intercostal and diaphragmatic muscularisation, SMA expression then appears to diminish in late gestation [70]. Although the impact on SMA expression in other thoracic organs (e.g. oesophagus) was not described, it could be that fetal breathing is important for the maintenance of ASM. Assuming the degree of lung hypoplasia manifest in amyogenic mutants is worse than those lacking intercostals alone, the late loss of ASM peristalsis (when it should be most active and widespread) may contribute to this.
Therefore, contrary to the view that ASM is ‘the appendix of the lung’ with no useful function [90–93], it appears that ASM is critical to aspects of prenatal lung growth: it provides FGF10 (from progenitors), mechanical stretch-relaxation and even perhaps lung liquid production. Encouraged by our discoveries, we began to explore the role of smooth muscle contractility in the development of other hollow organs using gut organ culture and calcium fluorophores. Our idea has been eagerly taken over by colleagues who continue to look at prenatal gut contractility using these techniques.
Future directions and potential clinical applications
Having considered the biological roles of prenatal ASM (and some of the mechanisms by which it may perform them), we can now consider potential clinical applications. In translating prenatal ASM biology from bench to bedside, technological development may have a significant part to play. Improved imaging +/− remote telemetry could allow us to visualise AP and/or measure resulting luminal pressure changes in utero [94, 95]. Given the observed relationship between peristaltic frequency and gestation [76], such measures might be used to provide a readily repeatable, non-biochemical estimate of lung maturity. This would be of great benefit in selecting cases for prenatal intervention or expeditious early delivery. Moreover, lung development following, e.g. tracheal occlusion might be monitored in real time by looking at peristalsis frequency. Following evidence that cyclical tracheal occlusion may be superior to static devices [96], there is also potential for devices that allow balloon pump occlusion of the trachea to optimise resulting lung development (‘preventilation“/dynamicPLUG) [46]. Imaging prenatal ASM contractility could even perhaps allow us to identify asthma-predispostions prenatally and discover new drug targets to combat ASM hypertrophy and airway remodelling in asthma (the commonest chronic disease in the developed world) [97].
Finally, the increasing frequency of ASM peristalsis through gestation and the unexplained switch from peristalsis to tonic contractility around birth could be important in the timing of parturition. Teleologically, it would be advantageous for some aspect(s) of fetal lung development to signal readiness for parturition. Could ASM peristalsis play a role in scheduling the changes in uterine contractility that herald labour? If ASM peristalsis is part of a pulmonary parturition clock, could we identify pregnancies at risk of preterm labour from airway contractility patterns long before uterine contractility supervenes [98]? Moreover, could such understanding lead us to appreciate that some lung disease associated with prematurity is cause rather than simply effect?
Closing remarks
We already know that ASM can transform lung morphology postnatally. In asthma, transient ASM constriction contributes to reversible lung hyperinflation; chronic ASM-related airway remodelling then participates in irreversible hyper-expansion supervening [99]. Lung hyperinflation that follows tracheal occlusion [100, 101] could even be seen as mimicking hyperinflation due to ASM-mediated small airway occlusions in asthma. Promisingly, pharmacological suppression of ASM hyperreactivity may prevent, halt or even reverse many of these changes in postnatal lung morphology [93, 99]. Given such dramatic pulmonary sequelae of postnatal ASM activity and the effects of ASM-targeted therapy, it is rational to explore the roles of ASM in prenatal lung morphogenesis (and the potential of ASM-related therapy). Our clinical experience reminds us of the toll that abnormalities of lung growth (prematurity, hypoplasia, CCAM, etc.) take on our patients. Therefore by understanding the role(s) of prenatal ASM, paediatric surgeons can make a further contribution to knowledge and therapy of the lung pathologies that we encounter in daily practice.
References
Sweed Y, Puri P (1993) Congenital diaphragmatic hernia: influence of associated malformations on survival. Arch Dis Child 69(1 Spec No):68–70
Smith NP, Jesudason EC, Featherstone NC, Corbett HJ, Losty PD (2005) Recent advances in congenital diaphragmatic hernia. Arch Dis Child 90(4):426–428
Smith NP, Losty PD, Connell MG, Mayer U, Jesudason EC (2006) Abnormal lung development precedes oligohydramnios in a transgenic murine model of renal dysgenesis. J Urol 175(2):783–786
Tsakayannis DE, Zurakowski D, Lillehei CW (1996) Respiratory insufficiency at birth: a predictor of mortality for infants with omphalocele. J Pediatr Surg 31(8):1088–1090
Wilson RD, Hedrick HL, Liechty KW, Flake AW, Johnson MP, Bebbington M, Adzick NS (2006) Cystic adenomatoid malformation of the lung: review of genetics, prenatal diagnosis, and in utero treatment. Am J Med Genet A 140(2):151–155
Grethel EJ, Wagner AJ, Clifton MS, Cortes RA, Farmer DL, Harrison MR, Nobuhara KK, Lee H (2007) Fetal intervention for mass lesions and hydrops improves outcome: a 15-year experience. J Pediatr Surg 42(1):117–123
Jobe AH, Ikegami M (2000) Lung development and function in preterm infants in the surfactant treatment era. Annu Rev Physiol 62:825–846
Muratore CS, Kharasch V, Lund DP, Sheils C, Friedman S, Brown C, Utter S, Jaksic T, Wilson JM (2001) Pulmonary morbidity in 100 survivors of congenital diaphragmatic hernia monitored in a multidisciplinary clinic. J Pediatr Surg 36(1):133–140
Koivusalo A, Pakarinen MP, Turunen P, Saarikoski H, Lindahl H, Rintala RJ (2005) Health-related quality of life in adult patients with esophageal atresia—a questionnaire study. J Pediatr Surg 40(2):307–312
Harrison MR, Bressack MA, Churg AM, de Lorimier AA (1980) Correction of congenital diaphragmatic hernia in utero. II. Simulated correction permits fetal lung growth with survival at birth. Surgery 88(2):260–268
Harrison MR, Ross N, Noall R, de Lorimier AA (1983) Correction of congenital hydronephrosis in utero. I. The model: fetal urethral obstruction produces hydronephrosis and pulmonary hypoplasia in fetal lambs. J Pediatr Surg 18(3):247–256
Skinner SC, Hirschl RB, Bartlett RH (2006) Extracorporeal life support. Semin Pediatr Surg 15(4):242–250
Schnitzer JJ, Hedrick HL, Pacheco BA, Losty PD, Ryan DP, Doody DP, Donahoe PK (1996) Prenatal glucocorticoid therapy reverses pulmonary immaturity in congenital diaphragmatic hernia in fetal sheep. Ann Surg 224(4):430–437; discussion 437–439
Downard CD, Wilson JM (2003) Current therapy of infants with congenital diaphragmatic hernia. Semin Neonatol 8(3):215–221
Wung JT, Sahni R, Moffitt ST, Lipsitz E, Stolar CJ (1995) Congenital diaphragmatic hernia: survival treated with very delayed surgery, spontaneous respiration, and no chest tube. J Pediatr Surg 30(3):406–409
Keller RL, Moore P, Teitel D, Hawgood S, McQuitty J, Fineman JR (2006) Abnormal vascular tone in infants and children with lung hypoplasia: findings from cardiac catheterization and the response to chronic therapy. Pediatr Crit Care Med 7(6):589–594
Dillon PW, Cilley RE, Hudome SM, Ozkan EN, Krummel TM (1995) Nitric oxide reversal of recurrent pulmonary hypertension and respiratory failure in an infant with CDH after successful ECMO therapy. J Pediatr Surg 30(5):743–744
Que J, Choi M, Ziel JW, Klingensmith J, Hogan BL (2006) Morphogenesis of the trachea and esophagus: current players and new roles for noggin and Bmps. Differentiation 74(7):422–437
Cardoso WV, Lu J (2006) Regulation of early lung morphogenesis: questions, facts and controversies. Development 133(9):1611–1624
Ellis K (1991) Fleischner lecture. Developmental abnormalities in the systemic blood supply to the lungs. AJR Am J Roentgenol 156(4):669–679
Hislop A (2005) Developmental biology of the pulmonary circulation. Paediatr Respir Rev 6(1):35–43
van Tuyl M, Groenman F, Wang J, Kuliszewski M, Liu J, Tibboel D, Post M (2007) Angiogenic factors stimulate tubular branching morphogenesis of sonic hedgehog-deficient lungs. Dev Biol 303(2):514–526
Thebaud B, Abman SH (2007) Bronchopulmonary dysplasia-where have all the vessels gone? Am J Respir Crit Care Med 175(10):978–985
Diez-Pardo JA, Baoquan Q, Navarro C, Tovar JA (1996) A new rodent experimental model of esophageal atresia and tracheoesophageal fistula: preliminary report. J Pediatr Surg 31(4):498–502
Burri PH (1984) Fetal and postnatal development of the lung. Annu Rev Physiol 46:617–628
DiFiore JW, Wilson JM (1994) Lung development. Semin Pediatr Surg 3(4):221–232
Hislop AA (2002) Airway and blood vessel interaction during lung development. J Anat 201(4):325–334
Hirose S, Farmer DL, Lee H, Nobuhara KK, Harrison MR (2004) The ex utero intrapartum treatment procedure: looking back at the EXIT. J Pediatr Surg 39(3):375–380; discussion 375–380
Marwan A, Crombleholme TM (2006) The EXIT procedure: principles, pitfalls, and progress. Semin Pediatr Surg 15(2):107–115
Warburton D, Wuenschell C, Flores-Delgado G, Anderson K (1998) Commitment and differentiation of lung cell lineages. Biochem Cell Biol 76(6):971–995
Liu JY, Nettesheim P, Randell SH (1994) Growth and differentiation of tracheal epithelial progenitor cells. Am J Physiol 266(3 Pt 1):L296–L307
Olver RE, Walters DV, S MW (2004) Developmental regulation of lung liquid transport. Annu Rev Physiol 66:77–101
Jain L, Eaton DC (2006) Physiology of fetal lung fluid clearance and the effect of labor. Semin Perinatol 30(1):34–43
Finer NN (2004) Surfactant use for neonatal lung injury: beyond respiratory distress syndrome. Paediatr Respir Rev 5(Suppl A):S289–S297
Valls-i-Soler A, Alfonso LF, Arnaiz A, Alvarez FJ, Tovar JA (1996) Pulmonary surfactant dysfunction in congenital diaphragmatic hernia: experimental and clinical findings. Biol Neonate 69(5):318–326
Sullivan KM, Hawgood S, Flake AW, Harrison MR, Adzick NS (1994) Amniotic fluid phospholipid analysis in the fetus with congenital diaphragmatic hernia. J Pediatr Surg 29(8):1020–1023
Fenton BW, Lin CS, Seydel F, Macedonia C (1998) Lecithin can be detected by volume-selected proton MR spectroscopy using a 1.5 T whole body scanner: a potentially non-invasive method for the prenatal assessment of fetal lung maturity. Prenat Diagn 18(12):1263–1266
Clifton MS, Joe BN, Zektzer AS, Kurhanewicz J, Vigneron DB, Coakley FV, Nobuhara KK, Swanson MG (2006) Feasibility of magnetic resonance spectroscopy for evaluating fetal lung maturity. J Pediatr Surg 41(4):768–773
Shannon JM, Hyatt BA (2004) Epithelial-mesenchymal interactions in the developing lung. Annu Rev Physiol 66:625–645
Alescio T, Cassini A (1962) Induction in vitro of tracheal buds by pulmonary mesenchyme grafted on tracheal epithelium. J Exp Zool 150:83–94
Kreidberg JA, Sariola H, Loring JM, Maeda M, Pelletier J, Housman D, Jaenisch R (1993) WT-1 is required for early kidney development. Cell 74(4):679–691
You LR, Takamoto N, Yu CT, Tanaka T, Kodama T, Demayo FJ, Tsai SY, Tsai MJ (2005) Mouse lacking COUP-TFII as an animal model of Bochdalek-type congenital diaphragmatic hernia. Proc Natl Acad Sci USA 102(45):16351–16356
Jay PY, Bielinska M, Erlich JM, Mannisto S, Pu WT, Heikinheimo M, Wilson DB (2007) Impaired mesenchymal cell function in Gata4 mutant mice leads to diaphragmatic hernias and primary lung defects. Dev Biol 301(2):602–614
Jesudason EC, Connell MG, Fernig DG, Lloyd DA, Losty PD (2000) Early lung malformations in congenital diaphragmatic hernia. J Pediatr Surg 35(1):124–127; discussion 128
Jesudason EC, Connell MG, Fernig DG, Lloyd DA, Losty PD (2000) Cell proliferation and apoptosis in experimental lung hypoplasia. J Pediatr Surg 35(1):129–133
Jesudason EC (2006) Small lungs and suspect smooth muscle: congenital diaphragmatic hernia and the smooth muscle hypothesis. J Pediatr Surg 41(2):431–435
Metzger RJ, Krasnow MA (1999) Genetic control of branching morphogenesis. Science 284(5420):1635–1639
Ghabrial A, Luschnig S, Metzstein MM, Krasnow MA (2003) Branching morphogenesis of the Drosophila tracheal system. Annu Rev Cell Dev Biol 19:623–647
Hogan BL (2006) Building organs from buds, branches and tubes. Differentiation 74(7):323–325
Hogan BL (1999) Morphogenesis. Cell 96(2):225–233
Warburton D, Bellusci S, De Langhe S, Del Moral PM, Fleury V, Mailleux A, Tefft D, Unbekandt M, Wang K, Shi W (2005) Molecular mechanisms of early lung specification and branching morphogenesis. Pediatr Res 57(5 Pt 2):26R–37R
Maeda Y, Dave V, Whitsett JA (2007) Transcriptional control of lung morphogenesis. Physiol Rev 87(1):219–244
Zaupa P, Kleinlein B, Hollwarth ME (2007) Bilateral congenital diaphragmatic hernia and gastroschisis in a newborn: can low intrathoracic pressure prevent the pulmonary hypoplasia? Pediatr Surg Int. Epub ahead of print. 11 Jan 2007
Hedrick MH, Ferro MM, Filly RA, Flake AW, Harrison MR, Adzick NS (1994) Congenital high airway obstruction syndrome (CHAOS): a potential for perinatal intervention. J Pediatr Surg 29(2):271–274
Harding R, Hooper SB (1996) Regulation of lung expansion and lung growth before birth. J Appl Physiol 81(1):209–224
Moore KA, Polte T, Huang S, Shi B, Alsberg E, Sunday ME, Ingber DE (2005) Control of basement membrane remodeling and epithelial branching morphogenesis in embryonic lung by Rho and cytoskeletal tension. Dev Dyn 232(2):268–281
Sanchez-Esteban J, Wang Y, Filardo EJ, Rubin LP, Ingber DE (2006) Integrins beta1, alpha6, and alpha3 contribute to mechanical strain-induced differentiation of fetal lung type II epithelial cells via distinct mechanisms. Am J Physiol Lung Cell Mol Physiol 290(2):L343–L350
Ingber DE (2005) Mechanical control of tissue growth: function follows form. Proc Natl Acad Sci USA 102(33):11571–11572
Sekine K, Ohuchi H, Fujiwara M, Yamasaki M, Yoshizawa T, Sato T, Yagishita N, Matsui D, Koga Y, Itoh N, et al (1999) Fgf10 is essential for limb and lung formation. Nat Genet 21(1):138–141
Min H, Danilenko DM, Scully SA, Bolon B, Ring BD, Tarpley JE, DeRose M, Simonet WS (1998) Fgf-10 is required for both limb and lung development and exhibits striking functional similarity to Drosophila branchless. Genes Dev 12(20):3156–3161
Celli G, LaRochelle WJ, Mackem S, Sharp R, Merlino G (1998) Soluble dominant-negative receptor uncovers essential roles for fibroblast growth factors in multi-organ induction and patterning. EMBO J 17(6):1642–1655
Arman E, Haffner-Krausz R, Gorivodsky M, Lonai P (1999) Fgfr2 is required for limb outgrowth and lung-branching morphogenesis. Proc Natl Acad Sci USA 96(21):11895–11899
Bellusci S, Grindley J, Emoto H, Itoh N, Hogan BL (1997) Fibroblast growth factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 124(23):4867–4878
Jesudason EC, Connell MG, Fernig DG, Lloyd DA, Losty PD (2000) Heparin and in-vitro experimental lung hypoplasia. Pediatr Surg Int 16(4):247–251
Jesudason EC, Connell MG, Fernig DG, Lloyd DA, Losty PD (2000) In vitro effects of growth factors on lung hypoplasia in a model of congenital diaphragmatic hernia. J Pediatr Surg 35(6):914–922
Acosta JM, Thebaud B, Castillo C, Mailleux A, Tefft D, Wuenschell C, Anderson KD, Bourbon J, Thiery JP, Bellusci S, et al (2001) Novel mechanisms in murine nitrofen-induced pulmonary hypoplasia: FGF-10 rescue in culture. Am J Physiol Lung Cell Mol Physiol 281(1):L250–L257
Jesudason EC (2002) Challenging embryological theories on congenital diaphragmatic hernia: future therapeutic implications for paediatric surgery. Ann R Coll Surg Engl 84(4):252–259
Teramoto H, Yoneda A, Puri P (2003) Gene expression of fibroblast growth factors 10 and 7 is downregulated in the lung of nitrofen-induced diaphragmatic hernia in rats. J Pediatr Surg 38(7):1021–1024
Miller AA, Hooper SB, Harding R (1993) Role of fetal breathing movements in control of fetal lung distension. J Appl Physiol 75(6):2711–2717
Inanlou MR, Baguma-Nibasheka M, Keating MM, Kablar B (2006) Neurotrophins, airway smooth muscle and the fetal breathing-like movements. Histol Histopathol 21(9):931–940
Fewell JE, Hislop AA, Kitterman JA, Johnson P (1983) Effect of tracheostomy on lung development in fetal lambs. J Appl Physiol 55(4):1103–1108
Starrett RW, de Lorimier AA (1975) Congenital diaphragmatic hernia in lambs: hemodynamic and ventilatory changes with breathing. J Pediatr Surg 10(5):575–582
Headley BM, McDougall PN, Stokes KB, Dewan PA, Dargaville PA (2001) Left-lung-collapse bronchial deformation in giant omphalocele. J Pediatr Surg 36(6):846–850
McCray PB Jr (1993) Spontaneous contractility of human fetal airway smooth muscle. Am J Respir Cell Mol Biol 8(5):573–580
Jesudason EC, Smith NP, Connell MG, Spiller DG, White MR, Fernig DG, Losty PD (2005) Developing rat lung has a sided pacemaker region for morphogenesis-related airway peristalsis. Am J Respir Cell Mol Biol 32(2):118–127
Parvez O, Voss AM, de Kok M, Roth-Kleiner M, Belik J (2006) Bronchial muscle peristaltic activity in the fetal rat. Pediatr Res 59(6):756–761
Pandya HC, Innes J, Hodge R, Bustani P, Silverman M, Kotecha S (2006) Spontaneous contraction of pseudoglandular-stage human airspaces is associated with the presence of smooth muscle-alpha-actin and smooth muscle-specific myosin heavy chain in recently differentiated fetal human airway smooth muscle. Biol Neonate 89(4):211–219
Copland IB, Post M (2007) Stretch-activated signaling pathways responsible for early response gene expression in fetal lung epithelial cells. J Cell Physiol 210(1):133–143
Liu M, Skinner SJ, Xu J, Han RN, Tanswell AK, Post M (1992) Stimulation of fetal rat lung cell proliferation in vitro by mechanical stretch. Am J Physiol 263(3 Pt 1):L376–L383
Featherstone NC, Jesudason EC, Connell MG, Fernig DG, Wray S, Losty PD, Burdyga TV (2005) Spontaneous propagating calcium waves underpin airway peristalsis in embryonic rat lung. Am J Respir Cell Mol Biol 33(2):153–160
Roman J (1995) Effects of calcium channel blockade on mammalian lung branching morphogenesis. Exp Lung Res 21(4):489–502
Featherstone NC, Jesudason EC, Connell MG, Fernig DG, Wray S, Losty PD, Burdyga TV (2005) Sarcoplasmic reticulum regulates calcium transients, airway peristalsis and branching morphogenesis of lung explants in vitro. Proc Am Thorac Soc 2:A503
Featherstone NC, Connell MG, Fernig DG, Wray S, Burdyga TV, Losty PD, Jesudason EC (2006) Sarcoplasmic calcium reuptake is essential for lung branching morphogenesis and required for development and peristalsis of prenatal airway smooth muscle. In: 19th international symposium of pediatric surgical research, Florence, Italy
Mailleux AA, Kelly R, Veltmaat JM, De Langhe SP, Zaffran S, Thiery JP, Bellusci S (2005) Fgf10 expression identifies parabronchial smooth muscle cell progenitors and is required for their entry into the smooth muscle cell lineage. Development 132(9):2157–2166
Sakuntabhai A, Ruiz-Perez V, Carter S, Jacobsen N, Burge S, Monk S, Smith M, Munro CS, O‘Donovan M, Craddock N, et al (1993) Mutations in ATP2A2, encoding a Ca2+ pump, cause Darier disease. Nat Genet 21(3):271–277
Yang Y, Beqaj S, Kemp P, Ariel I, Schuger L (2000) Stretch-induced alternative splicing of serum response factor promotes bronchial myogenesis and is defective in lung hypoplasia. J Clin Invest 106(11):1321–1330
Belik J, Davidge ST, Zhang W, Pan J, Greer JJ (2003) Airway smooth muscle changes in the nitrofen-induced congenital diaphragmatic hernia rat model. Pediatr Res 53(5):737–743
Featherstone NC, Connell MG, Fernig DG, Wray S, Burdyga TV, Losty PD, Jesudason EC (2006) Airway smooth muscle dysfunction precedes teratogenic congenital diaphragmatic hernia and may contribute to hypoplastic lung morphogenesis. Am J Respir Cell Mol Biol 35(5):571–578
Jesudason EC, Smith NP, Connell MG, Spiller DG, White MR, Fernig DG, Losty PD (2006) Peristalsis of airway smooth muscle is developmentally regulated and uncoupled from hypoplastic lung growth. Am J Physiol Lung Cell Mol Physiol 291(4):L559–L565
Mitzner W (2004) Airway smooth muscle: the appendix of the lung. Am J Respir Crit Care Med 169(7):787–790
Seow CY, Fredberg JJ (2001) Historical perspective on airway smooth muscle: the saga of a frustrated cell. J Appl Physiol 91(2):938–952
Fredberg JJ, Mead J (2007) Airway smooth muscle is/is not useful. J Appl Physiol 102(4):1708–1710
Janssen LJ, Killian K (2006) Airway smooth muscle as a target of asthma therapy: history and new directions. Respir Res 7:123
Fox HE, Badalian SS, Timor-Tritsch IE, Marks F, Stolar CJ (1993) Fetal upper respiratory tract function in cases of antenatally diagnosed congenital diaphragmatic hernia: preliminary observations. Ultrasound Obstet Gynecol 3(3):164–167
Hellmeyer L, Exner C, Folz B, Hiller M, Sierra F, Lukasewitz P, Steinfeld T, Werner JA, Heldmaier G, Schmidt S (2007) Telemetric monitoring of tracheal pressure after tracheal occlusion for treatment of severe congenital diaphragmatic hernia. Arch Gynecol Obstet 275(4):245–248
Nelson SM, Hajivassiliou CA, Haddock G, Cameron AD, Robertson L, Olver RE, Hume R (2005) Rescue of the hypoplastic lung by prenatal cyclical strain. Am J Respir Crit Care Med 171(12):1395–1402
Barnes PJ (2004) The size of the problem of managing asthma. Respir Med 98(Suppl B):S4–S8
Kalache KD, Chaoui R, Marks B, Wauer R, Bollmann R (2002) Does fetal tracheal fluid flow during fetal breathing movements change before the onset of labour? BJOG 109(5):514–519
Holgate ST, Polosa R (2006) The mechanisms, diagnosis, and management of severe asthma in adults. Lancet 368(9537):780–793
DiFiore JW, Fauza DO, Slavin R, Peters CA, Fackler JC, Wilson JM (1994) Experimental fetal tracheal ligation reverses the structural and physiological effects of pulmonary hypoplasia in congenital diaphragmatic hernia. J Pediatr Surg 29(2):248–256
Jani J, Gratacos E, Greenough A, Piero JL, Benachi A, Harrison M, Nicolaides K, Deprest J (2005) Percutaneous fetal endoscopic tracheal occlusion (FETO) for severe left-sided congenital diaphragmatic hernia. Clin Obstet Gynecol 48(4):910–922
Acknowledgments
The author is supported by a National Clinician Scientist Fellowship (Academy of Medical Sciences/Health Foundation) and the Association of Laparoscopic Surgeons of Great Britain and Ireland David Dunn Scholarship. The Medical Research Council, Birth Defects Foundation and Children’s Research Fund have also funded the work described. I should like to thank (1) my patients and their families for consent to use their imaging, (2) Professor PD Losty, Mrs. MG Connell, Miss NP Smith, Mr. NC Featherstone, Dr. TV Burdyga, Professor DG Fernig, Professor S Wray, Professor MR White and Dr. DG Spiller for their contributions to these studies and (3) staff at UCSF Fetal Treatment Center/Pediatric Surgery for exposure to fetal surgical models.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Jesudason, E.C. Exploiting mechanical stimuli to rescue growth of the hypoplastic lung. Pediatr Surg Int 23, 827–836 (2007). https://doi.org/10.1007/s00383-007-1956-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00383-007-1956-0