Article Text

Abstract
Rationale Cellular senescence is observed in the lungs of patients with COPD and may contribute to the disease pathogenesis. Growth differentiation factor 11 (GDF11) belongs to the transforming growth factor β superfamily and was recently reported to be a circulating protein that may have rejuvenating effects in mice. We aimed to investigate the amounts of GDF11 in the plasma and the lungs of patients with COPD and elucidate the possible roles of GDF11 in cellular senescence.
Methods The plasma levels of GDF11 were investigated in two separate cohorts by western blotting. The localisation and expression of GDF11 in the lungs were investigated by immunohistochemistry and quantitative reverse transcription PCR, respectively. The effects of GDF11 on both cigarette smoke extract (CSE)-induced cellular senescence in vitro and on elastase-induced cellular senescence in vivo were investigated.
Results The levels of plasma GDF11 in the COPD group were decreased compared with the control groups in the two independent cohorts. The levels of plasma GDF11 were significantly positively correlated with pulmonary function data. The mRNA expression of GDF11 in mesenchymal cells from the COPD group was decreased. Chronic exposure to CSE decreased the production of GDF11. Treatment with GDF11 significantly inhibited CSE-induced cellular senescence and upregulation of inflammatory mediators, partly through Smad2/3 signalling in vitro. Daily GDF11 treatment attenuated cellular senescence and airspace enlargement in an elastase-induced mouse model of emphysema.
Conclusions The decrease in GDF11 may be involved in the cellular senescence observed in COPD.
- COPD Pathology
- Oxidative Stress
Statistics from Altmetric.com
Key messages
What is the key question?
Is growth differentiation factor 11 (GDF11), a possible rejuvenating factor, decreased in the plasma and lungs of patients with COPD?
What is the bottom line?
The levels of GDF11 are decreased in the plasma and lungs of patients with COPD and are inversely correlated with the severity of airflow limitation; treatment with GDF11 inhibits cigarette smoke extract induced cellular senescence in vitro and cellular senescence in an elastase-induced in vivo mouse model of emphysema.
Why read on?
Our findings suggest that GDF11 may be associated with the cellular senescence observed in COPD and that the investigation of GDF11-related signalling may potentially clarify the mechanism of cellular senescence in COPD.
Introduction
COPD is a major cause of chronic morbidity and mortality throughout the world.1 A protease–antiprotease imbalance and oxidative stress have been reported to be involved in the pathogenesis of COPD.2 ,3 The age-dependent increase in the prevalence of COPD suggests an intimate relationship between the pathogenesis of COPD and ageing.4 ,5 Tsuji et al6 demonstrated that cellular senescence was accelerated in the alveolar cells of patients with COPD. Many reports have shown that cellular senescence in COPD is accelerated in many types of cells, including bronchial epithelial cells,7 lung fibroblasts,8 ,9 and circulating leukocytes,10 and that oxidative stress accelerates cellular senescence.11 ,12 For example, aged lung fibroblasts from patients with COPD showed an impaired tissue repair function that limited the renewal of damaged tissue.13 Furthermore, patients with COPD are well known to have comorbidities of chronic degenerative diseases such as hypertension, cardiovascular diseases and diabetes mellitus.14 ,15 Chronic degenerative diseases are thought to be associated with accelerated ageing. These findings suggest that cellular senescence not only occurs in the lungs of patients with COPD but may be able to explain several of the key features identified in the disease, and investigating cellular senescence could therefore become a useful approach for understanding the pathogenesis of COPD.
Growth differentiation factor 11 (GDF11) is a transforming growth factor member first identified in 1999.16 ,17 Recent studies have shown that restoring the systemic GDF11 levels in old mice reversed age-related skeletal muscle dysfunction18 and promoted neovascularisation and the proliferation of neural stem cells in the brain.19 Further, Loffredo et al20 showed that a circulating factor in young mice reversed age-related cardiac hypertrophy, and they identified that the rejuvenating factor was GDF11. Recently, a large cohort study in the US was carried out to examine the link between plasma GDF11 levels and cardiovascular events. The levels of plasma GDF11/8 were closely associated with cardiovascular outcomes and overall mortality in humans.21 These results suggest that GDF11 might play an anti-ageing role in cardiovascular diseases. However, the anti-ageing role of GDF11 in COPD, in which senescence could be involved in the pathogenesis, has not been elucidated yet.
In the current study, to reveal the possible link between GDF11 and senescence in COPD, we measured the plasma GDF11 levels in patients with COPD and control subjects in two independent cohorts. Further, we examined the expression of GDF11 in the lungs. We also investigated the anti-senescent effects of GDF11 on stress-induced or replicative senescence in in vitro and in vivo experiments.
Materials and methods
Additional details on the materials and methods are provided in an online data supplement.
supplementary data
Recruitment of the study subjects
Study subjects from two separate cohorts took part in the study. Study subjects from Tohoku University Hospital and Tohoku Rosai Hospital participated in the cohort 1 study between January 2015 and July 2016. All patients with COPD satisfied the Global Initiative for Chronic Obstructive Lung Disease (GOLD) guideline criteria.1 Age-matched control never-smokers and control ex-smokers were enrolled. Ex-smokers had smoking history of ≥10 pack-years and had quit smoking for at least 1 year. Current smokers and patients with asthma were excluded. To validate the data obtained from the first cohort, study subjects who visited Japanese Red Cross Ishinomaki Hospital between November 2012 and December 2014 were included in the cohort 2 study. In total, 112 of 126 subjects in cohort 1 and 87 of 99 subjects in cohort 2 met our inclusion/exclusion criteria.
Measurement of plasma GDF11
The production of GDF11 in plasma was investigated by western blotting. An anti-GDF11 antibody (R&D Systems, Minneapolis, Minnesota, USA) was used for the western blotting analyses in the current study. Further details are available in the online data supplement.
Preparation of human lung tissues and primary lung cells
The study subjects had undergone a surgical operation for lung cancer after receiving pulmonary function tests. Peripheral lung tissues were obtained from the subpleural parenchyma of the lobe resected at surgery, avoiding areas involving tumours. The tissues were used for immunohistochemical staining, single cell studies and culture of lung fibroblasts and bronchial epithelial cells.
Animal studies
To induce pulmonary emphysema, mice were treated intratracheally with porcine pancreatic elastase (25 µg/mouse) on day 0.22 Treatment with recombinant GDF11 (0.1 mg/kg) or vehicle was repeated daily (5 days/week) until day 4 or 21, after which the mice were sacrificed for analyses. Further details are available in the online data supplement.
Ethics
Written informed consent was obtained from all subjects who participated in the current study. All experiments in the current study were approved by the ethics committee of Tohoku University Graduate School of Medicine, Tohoku Rosai Hospital, and Japanese Red Cross Ishinomaki Hospital. All animal experiments were approved by the Tohoku University Animal Experiment Ethics Committee and performed in accordance with the Regulations for Animal Experiments and Related Activities at Tohoku University.
Statistical analysis
The data are expressed as the means±SD. A linear regression analysis was performed to assess the association between GDF11 and FEV1% predicted, diffusing capacity of the lung for carbon monoxide (DLCO)/alveolar volume (VA)% predicted, smoking history, age and bone density using the method of least squares. The strength of association was evaluated using Spearman's rank test. A multiple regression analysis was used to evaluate the associations between plasma GDF11 levels, age, sex, smoking history, FEV1% predicted values and the use of inhaled corticosteroids. Experiments with multiple comparisons were evaluated by a one-way analysis of variance followed by Tukey's test to adjust for multiple comparisons. GraphPad Prism V.6 (GraphPad Software, Inc., San Diego, California, USA) and JMP Pro V.12.2.0 (SAS Institute Inc., Tokyo, Japan) were used for all statistical analyses. Probability values of <0.05 were considered significant.
Results
Analysis of plasma GDF11 levels
Twenty control never-smokers, 23 control ex-smokers and 69 ex-smokers with COPD took part in the study (table 1). The levels of plasma GDF11 were significantly decreased in patients with COPD (0.57±0.27) compared with the control never-smokers (0.95±0.29, p<0.001) or control ex-smokers (0.84±0.27, p<0.001; figure 1A, B). The levels of GDF11 were significantly correlated with FEV1% predicted (r=0.57, p<0.001; figure 1C), DLCO/VA% predicted (r=0.41, p<0.01; figure 1D) and smoking history (r=−0.33, p<0.01; figure 1E), but not with age (figure 1F). To confirm whether GDF11 production was decreased in the patients with COPD, we examined a separate cohort (cohort 2; table 2). The levels of plasma GDF11 were significantly lower in the patients with COPD (0.55±0.24) compared with the control never-smokers (0.98±0.30 p<0.001) and control ex-smokers (0.97±0.29 p<0.001; figure 2A, B). The levels of plasma GDF11 were significantly correlated with FEV1% predicted (r=0.59, p<0.001; figure 2C) and smoking history (r=−0.50, p<0.001; figure 2D), but not with age (figure 2E). In both cohorts, the relationship between the levels of plasma GDF11 and FEV1% predicted remained after adjusting for age, sex, smoking history and use of inhaled corticosteroids (see online supplementary table S1). To investigate the relationship of GDF11 with ageing, we measured the levels of GDF11 in 60 control never-smokers between 20 and 70 years old. The levels of plasma GDF11 did not change with age (see online supplementary figure S1A,B). We investigated the relationship between the levels of plasma GDF11 and the number of comorbidities of all patients with COPD in both cohorts. Interestingly, the levels of GDF11 were significantly lower in the patients with COPD who had more than three age-related comorbidities (p<0.05; figure 2F). The levels of plasma GDF11 were correlated with bone density in cohort 1 (r=0.35, p<0.01; see online supplementary figure S1C).
Characteristics of the subjects in cohort 1
Characteristics of the subjects in cohort 2

The levels of plasma growth differentiation factor 11 (GDF11) and the correlations between the levels of GDF11 and clinical parameters in cohort 1. Plasma was obtained from the control never-smokers (CNS, n=20), the control ex-smokers (CES, n=23), and the ex-smokers with COPD (COPD, n=69) in cohort 1. The protein expression of GDF11 in plasma was investigated by western blotting (A). Ponceau S staining was used to evaluate the amount of protein loaded in each lane. The GDF11 level was calculated by measuring the intensity of the band (B). Correlations between the levels of GDF11 and the values of FEV1% predicted (C), DLCO/VA% predicted (D), smoking (E) and age (F) were investigated. r is the correlation coefficient; the lines and p values correspond to the regression equation. Open circles: CNS; closed circles (grey): CES; closed circles (black): COPD. ***p<0.001 compared with CNS, †††p<0.001 compared with CES; DLCO, diffusing capacity of the lung for carbon monoxide; VA, alveolar volume.

The levels of plasma growth differentiation factor 11 (GDF11) and the correlations between the levels of GDF11 and clinical parameters in cohort 2. Plasma was obtained from the control never-smokers (CNS, n=15), the control ex-smokers (CES, n=16), and the ex-smokers with COPD (COPD, n=56) in cohort 2. The protein expression of GDF11 in plasma was investigated by western blotting (A). Ponceau S staining was used to evaluate the amount of protein loaded in each lane. The GDF11 level was calculated by measuring the intensity of the band (B). Correlations between the levels of GDF11 and the values of FEV1% predicted (C), smoking (D) and age (E) were investigated. The relationship between the levels of plasma GDF11 and the number of comorbidities is shown for the patients with COPD in the two cohorts (F). Comorbidities including hypertension, hyperlipidaemia, diabetes mellitus, cardiovascular diseases and cancer were examined. r is the correlation coefficient; the lines and p values correspond to the regression equation. Open circles: CNS; closed circles (grey): CES; closed circles (black): COPD. ***p<0.001 compared with CNS, †††p<0.001 compared with CES, ‡p<0.05 compared with the patient group without any comorbidities.
Analysis of GDF11 expression in the lung
Next, we examined the expression of GDF11 in human lungs by immunohistochemistry. The expression of GDF11 was mainly detected in mesenchymal cells within the airway walls and airway epithelial cells (figure 3).

Immunohistochemical localisation of growth differentiation factor 11 (GDF11) in human small airways. Representative photographs of the immunoreactivity against GDF11 in human small airways are shown. The upper panels show low-magnification images (×40; scale bar, 500 µm), and the lower panels show magnified images (×100; scale bar, 200 µm). Arrows indicate GDF11 immunopositive cells. CNS, control never-smoker; CES, control ex-smoker; COPD, ex-smoker with COPD.
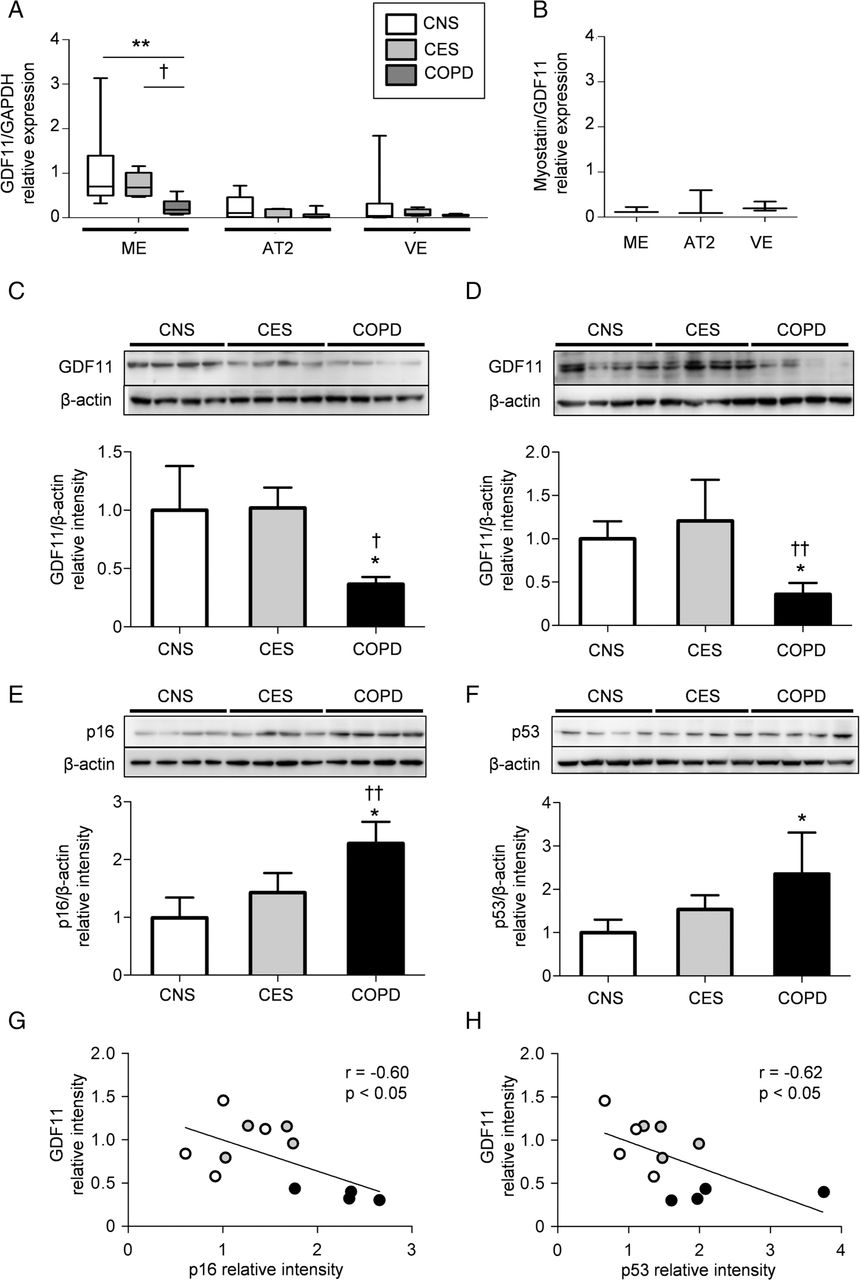
To specify which types of cells mainly express GDF11 in lungs, we prepared human lung single cell suspensions and sorted them into mesenchymal cells, alveolar type II cells and vascular endothelial cells using a cell sorter. The characteristics of the study subjects are shown in table 3. The expression of GDF11 mRNA in mesenchymal cells from the patients with COPD was significantly lower than in those from the control subjects (figure 4A). Because myostatin is highly related to GDF11 and the two proteins exhibit 90% homology in their mature active regions,16 we investigated the expression of myostatin/GDF11 in the lung cells. The expression of myostatin was much lower than that of GDF11 in lung resident cells (figure 4B). We further examined the protein levels of GDF11 in primary lung fibroblasts and bronchial epithelial cells from the study subjects. In an unstimulated condition, the amounts of GDF11 in the lung fibroblasts and the bronchial epithelial cells from the COPD group were significantly lower than in those from the control subjects (figure 4C, D). To clarify the relationship between GDF11 and cellular senescence, we examined the production of senescence-associated proteins in the primary lung fibroblasts. As expected, the amounts of p16 and p53 in the cells from patients with COPD were significantly increased compared with those in cells from the control groups (figure 4E, F). The level of GDF11 was negatively correlated with those of p16 (r=−0.60, p<0.05; figure 4G) and p53 (r=−0.62, p<0.05; figure 4H), suggesting that GDF11 might be associated with lung cellular senescence.
Characteristics of the subjects in the single cell study

Expression of growth differentiation factor 11 (GDF11) in human lung cells. Lung tissues were obtained from the study subjects. Single lung cell suspensions were sorted into mesenchymal cells (ME), alveolar type II cells (AT2) and vascular endothelial cells (VE) using a cell sorter. Gene expression was determined by quantitative reverse transcription PCR (qRT-PCR). GDF11 relative expression was calculated by dividing the expression of GDF11 by the expression of the appropriate GAPDH mRNA in each cell type (A). Values are expressed as the mean±SD (n=5–10). The relative expression of myostatin/GDF11 in each cell type from the CNS group is shown (B). Values are expressed as the mean±SD (n=3). Four different strains of lung fibroblasts and bronchial epithelial cells were obtained from each study group. The amounts of GDF11 in the lung fibroblasts (C) and bronchial epithelial cells (D) and the expression of p16 (E) and p53 (F) in the lung fibroblasts were analysed by western blotting. The band intensity of GDF11, p16 and p53 was standardised to that of β-actin. Values are expressed as the mean±SD (n=4). Correlations between the levels of GDF11 and p16 (G) or p53 (H) in the primary lung fibroblasts from the study subjects were investigated. Open circles: CNS; closed circles (grey): CES; closed circles (black): COPD. *p<0.05, **p<0.01 compared with CNS, †p<0.05, ††p<0.01 compared with CES. CNS, control never-smoker; CES, control ex-smoker; COPD, ex-smoker with COPD.
Effects of GDF11 on cigarette smoke extract induced cellular senescence and replicative cellular senescence in lung resident cells
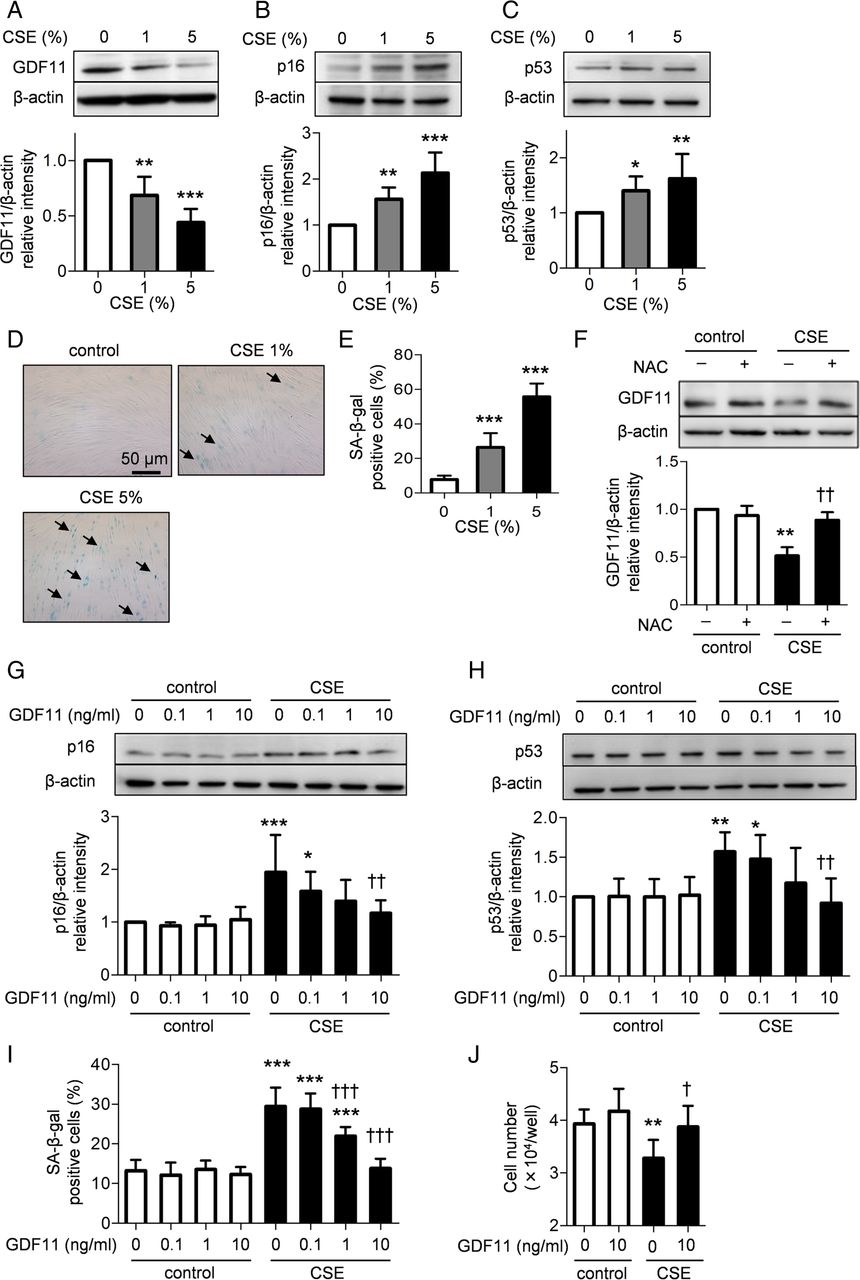
We next evaluated whether GDF11 ameliorated the cellular senescence in lung resident cells. In general, two main senescence-associated pathways have been identified. One is replicative senescence caused by telomere shortening, and the alternative pathway is stress-induced premature senescence.4 ,23 First, we investigated the effects of GDF11 on stress-induced cellular senescence. To examine the role of oxidative stress on stress-induced cellular senescence, we used an in vitro model of cigarette smoke extract (CSE) exposure. Human fetal lung fibroblasts (HFL-1) were continually exposed to various concentrations of CSE for 10 days. Chronic exposure to 1% CSE significantly reduced the production of GDF11 compared with that of the control group (p<0.01; figure 5A), whereas 1% CSE significantly augmented the expression of p16 (p<0.01; figure 5B) and p53 (p<0.05; figure 5C) as well as the intracellular accumulation of senescence-associated β-galactosidase (SA-β-gal, p<0.001; figure 5D, E). We hypothesised that oxidative stress might be involved in the CSE-mediated attenuation of GDF11 expression. Pretreatment with N-acetyl cysteine (NAC), a scavenger of reactive oxygen species (ROS), prevented the reduction in GDF11 expression by exposure to CSE (p<0.01; figure 5F). Treatment with GDF11 significantly inhibited the CSE-augmented expression of p16 (p<0.01; figure 5G) and p53 (p<0.01; figure 5H) and the CSE-augmented SA-β-gal activity (p<0.001; figure 5I) in a concentration-dependent manner. CSE significantly delayed cell proliferation, but treatment with GDF11 ameliorated the delay in cell growth (p<0.05; figure 5J). Similar anti-senescent effects of GDF11 were observed in primary bronchial epithelial cells. Treatment with GDF11 significantly ameliorated the CSE-induced cellular senescence as assessed by the expression of senescence-associated proteins (figure 6A, B), SA-β-gal activity (figure 6C, D) and cell proliferation (figure 6E). These results suggest that GDF11 could prevent cigarette smoke from accelerating lung cellular senescence.

Effects of growth differentiation factor 11 (GDF11) on cigarette smoke extract (CSE)-induced cellular senescence in human fetal lung fibroblasts (HFL-1). HFL-1 cells were exposed to various concentrations of CSE from the 19th passage to the 21st passage for a total of 10 days. The expression of GDF11 and senescence-associated proteins was evaluated by western blotting. Relative intensity was calculated by dividing the intensity of the GDF11 (A), p16 (B) and p53 (C) bands by that of the appropriate β-actin band. Senescence-associated β-galactosidase (SA-β-gal)-positive cells at various concentrations of CSE were investigated (D and E). Arrows indicate SA-β-gal-positive cells. The cells were exposed to 1% CSE in the presence or absence of 1 mM N-acetyl cysteine (NAC) for 10 days. The expression of GDF11 was evaluated by western blotting (F). The cells were exposed to 1% CSE in the presence or absence of various concentrations of GDF11 for 10 days. The expression of p16 (G) and p53 (H), SA-β-gal positive cells (I) and cell number (J) were investigated at the 21st passage. Values are expressed as the mean±SD (n=8) and are representative of two independent experiments. *p<0.05, **p<0.01, ***p<0.001 compared with the control group, †p<0.05, ††p<0.01, †††p<0.001 compared with the CSE-treated group.

Effects of growth differentiation factor 11 (GDF11) on cigarette smoke extract (CSE)-induced cellular senescence in bronchial epithelial cells. Lung samples were obtained from healthy control subjects, and primary bronchial epithelial cells were cultured. The bronchial epithelial cells were exposed to 1% CSE with or without 10 ng/mL GDF11 at the 5th passage for 10 days. The expression of senescence-associated proteins was evaluated by western blotting. Relative intensity was calculated by dividing the intensity of each p16 (A) and p53 (B) band by that of the appropriate β-actin band. SA-β-gal-positive cells were investigated (C and D). Arrows indicate SA-β-gal-positive cells (C). Cell numbers were investigated (E). Values are expressed as the mean±SD (n=4). *p<0.05, **p<0.01, ***p<0.001 compared with the control group, †p<0.05, †††p<0.001 compared with the CSE-treated group.
To investigate the effects of GDF11 on replicative senescence, HFL-1 cells were cultured to the 27th passage. As the cells had been cultured for 5 weeks, the production of GDF11 had dramatically declined, whereas the expression of p16 and p53 and the activity of SA-β-gal were augmented (see online supplementary figure S2A–D). To evaluate the effects of GDF11 on replicative senescence, the cells were cultured with or without GDF11 from the 16th to the 27th passage for 5 weeks. GDF11 had no effect on p53 expression, SA-β-gal activity or cell proliferation (see online supplementary figure S2E–G), suggesting that GDF11 was unlikely to prevent replicative senescence in vitro.
Effects of GDF11 on CSE-induced ROS generation, inflammatory mediators, and impairment of tissue repair function
Oxidative stress is thought to be an important amplifying mechanism in COPD and is well known to accelerate cellular senescence in various types of cells.11 ,12 We hypothesised that GDF11 might have a protective effect against CSE-induced cellular senescence through the suppression of ROS generation. Exposure to CSE significantly increased the ROS activity in HFL-1 cells, and treatment with GDF11 suppressed the CSE-induced ROS activity to the control level (figure 7A, B). To further examine the effects of GDF11, the release of CSE-induced inflammatory mediators including matrix metalloproteinases (MMPs) and interleukin (IL)-8 was evaluated in HFL-1 cells. CSE significantly enhanced the release of MMP-9 and IL-8, and treatment with GDF11 significantly suppressed the CSE-induced inflammatory mediators (figure 7C–F). Further, we investigated the effects of GDF11 on the impaired tissue repair function induced by CSE. Treatment with GDF11 significantly restored fibroblast-mediated tissue repair as assessed by a collagen gel contraction assay (figure 7G).

Effects of growth differentiation factor 11 (GDF11) on cigarette smoke extract (CSE)-induced reactive oxygen species (ROS) generation, release of matrix metalloproteinases (MMPs) and interleukin (IL)-8 and fibroblast-mediated tissue repair function. HFL-1 cells were exposed to 7% CSE in the presence or absence of 10 ng/mL GDF11 for 24 hours. The cells were stained with CellRox Deep Red reagent to determine the intracellular ROS level. The flow cytometry data are shown as the mean fluorescence intensity (MFI) (A and B). The cells were exposed to 7% CSE in the presence or absence of 10 ng/mL GDF11 for 24 hours. The effect of GDF11 on the CSE-augmented release of MMP-9 was investigated by gelatine zymography. The latent form of MMP-9 (C and D) and the active form of MMP-9 (C and E) were evaluated. The cells were exposed to 1% CSE from the 19th passage to the 21st passage for 10 days. IL-8 release in the medium was quantified using an ELISA kit (F). Fibroblast-mediated tissue repair function was evaluated by a three-dimensional collagen gel contraction assay. The collagen gels were treated with or without 5 ng/mL GDF11 and then 7% CSE was added to the medium. The gel size was evaluated on day 5 (G). Values are expressed as the mean±SD (n=3–4). *p<0.05, **p<0.01 compared with the control group, †p<0.05, ††p<0.01 compared with the CSE-treated group.
Signal transduction of GDF11 and the effects of an activin receptor-like kinase inhibitor on CSE-induced cellular senescence
To explore the signalling of GDF11, we investigated the phosphorylation of Smad in lung fibroblasts. As previously reported in other types of cells,19 ,24 ,25 GDF11 phosphorylated Smad2/3, and the phosphorylation was inhibited by an activin receptor-like kinase (ALK)4/5 inhibitor, SB431542, in HFL-1 cells (see online supplementary figure S2A). The ALK4/5 inhibitor partly but significantly inhibited the anti-senescent effect of GDF11 on CSE-induced SA-β-gal accumulation (see online supplementary figure S2B).
Effects of GDF11 on CSE-induced cellular senescence in COPD lung cells
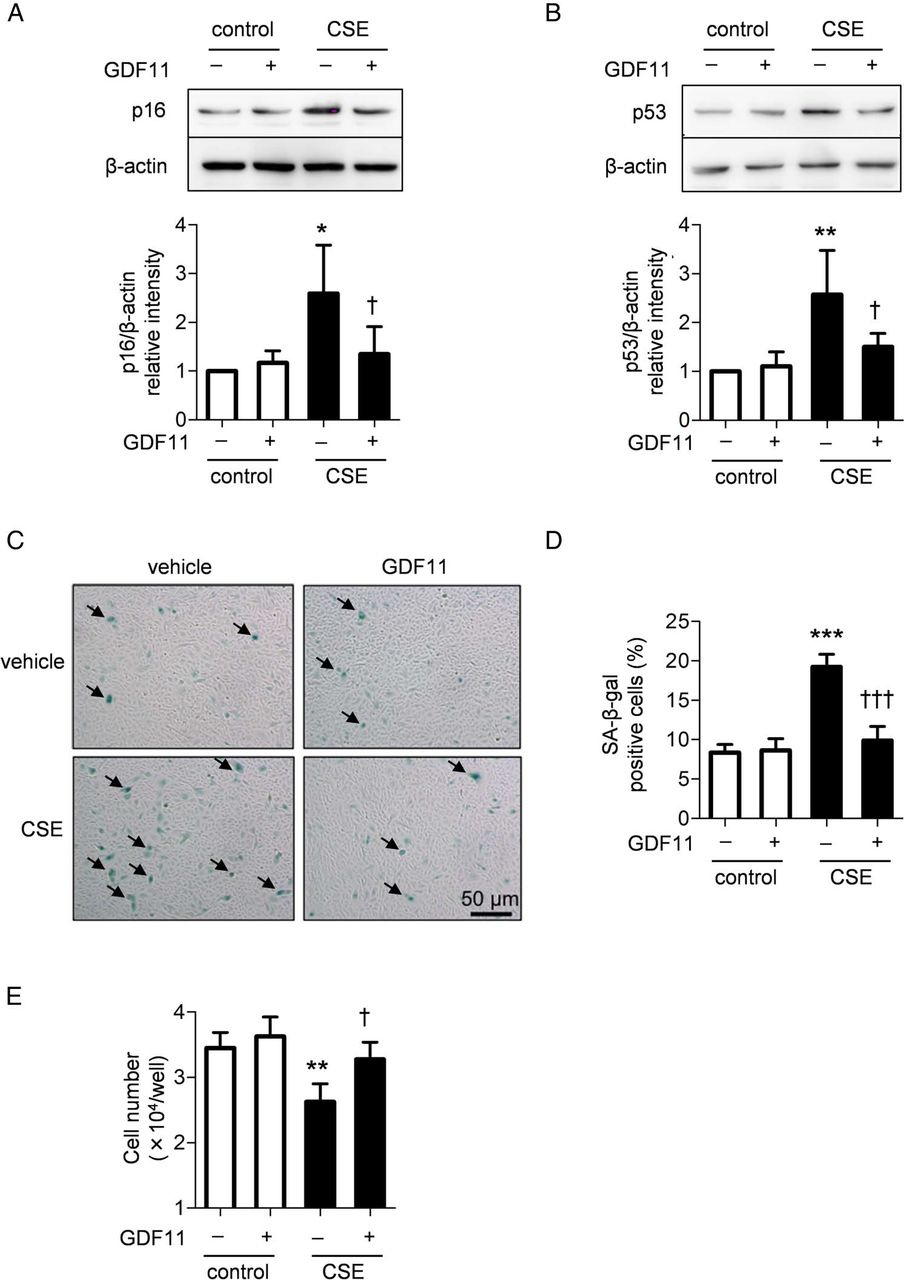
To investigate the effects of GDF11 on cellular senescence in COPD lung cells, primary lung fibroblasts and bronchial epithelial cells from the patients with COPD were treated with GDF11 or vehicle in the presence or absence of CSE. Exposure to CSE significantly enhanced the levels of all senescence markers in both types of cells (figure 8A–D). Treatment with GDF11 significantly suppressed the CSE-augmented expression of p16 (p<0.01; figure 8A), SA-β-gal activity in the COPD lung fibroblasts (p<0.01; figure 8C) and SA-β-gal activity in the COPD bronchial epithelial cells (p<0.05; figure 8D), whereas GDF11 had no effect on p53 expression in the lung fibroblasts (figure 8B). These results suggest that GDF11 inhibited CSE-accelerated cellular senescence, even in COPD lung cells.

Effects of growth differentiation factor 11 (GDF11) on cigarette smoke extract (CSE)-induced cellular senescence in COPD lung cells. Four different strains of lung fibroblasts and bronchial epithelial cells were obtained from the patients with COPD. COPD lung fibroblasts were exposed to 1% CSE in the presence or absence of 10 ng/mL GDF11 for 10 days. The expression of p16 (A) and p53 (B) and SA-β-gal activity (C) were investigated at the 8th passage. COPD bronchial epithelial cells were exposed to 5% CSE in the presence or absence of 10 ng/mL GDF11 for 24 hours. SA-β-gal activity was investigated at the 5th passage (D). Values are expressed as the mean±SD (n=4). *p<0.05, **p<0.01 compared with the control group, †p<0.05, ††p<0.01 compared with the CSE-treated group.
Effects of GDF11 on cellular senescence in an elastase-induced mouse model of emphysema
Finally, to investigate the effects of GDF11 on cellular senescence in mice, mice were exposed to elastase and received daily GDF11 treatments for 3 weeks. Intraperitoneal administration of GDF11 elevated the plasma GDF11 level and induced lung Smad2/3 phosphorylation (see online supplementary figure S4A,B), suggesting that the GDF11 administration was effective in the lungs. Intratracheal injection of elastase caused lung inflammation on day 4, whereas administration of GDF11 did not alter the inflammatory responses and airspace enlargement induced by elastase (see online supplementary figure S4C–G). On day 21, elastase exposure reduced the production of GDF11 in the lungs (figure 9A) and augmented the expression of p16 (figure 9B) and p53 (figure 9C) compared with the phosphate-buffered saline administration group. GDF11 attenuated the accelerated cellular senescence (figure 9B–E) and ameliorated elastase-induced airspace enlargement (p<0.05; figure 9F, G). These results showed the protective effect of GDF11 against elastase-induced cellular senescence and pulmonary emphysema in mice.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of growth differentiation factor 11 (GDF11) on cellular senescence in an elastase-induced mouse model of emphysema. Mice were exposed to elastase (25 µg/mouse) or phosphate-buffered saline (PBS) intratracheally on day 0 and treated with 0.1 mg/kg GDF11 or vehicle intraperitoneally for 3 weeks. The expression of GDF11 (A), p16 (B) and p53 (C) in the whole lung homogenates was investigated by western blotting on day 21. SA-β-gal activity was assessed by staining of the OCT-embedded frozen lung tissues (D) (magnification, ×200; scale bar, 100 µm) and measurement of fluorescence intensity in the whole lung homogenates (E). Lung sections were stained with H&E (F). The upper panels show low-magnification images (×40; scale bar, 500 µm), and the lower panels show magnified images (×200; scale bar, 100 µm). Semi-quantitative analysis of lung tissues using mean linear intercept (Lm) is shown (G). Values are expressed as the mean±SD (n=4–8). **p<0.01, ***p<0.001 compared with the PBS-exposed vehicle-treated group, †p<0.05, ††p<0.01 compared with the elastase-exposed vehicle-treated group.
Discussion
In the present study, we demonstrated that the levels of plasma GDF11, an anti-ageing factor, were significantly decreased in patients with COPD and that the GDF11 levels were significantly correlated with the values of FEV1% predicted, lung diffusing capacity and smoking, but not age. GDF11 was localised in mesenchymal cells within the airway walls and airway epithelial cells. The expression of GDF11 in parenchymal cells from the patients with COPD was decreased. The levels of p16 and p53 in the primary lung fibroblasts were significantly negatively correlated with the level of GDF11. In the in vitro culture study, we observed that CSE attenuated the expression of GDF11 and that NAC inhibited the CSE-mediated attenuation of GDF11 expression, suggesting that oxidative stress could be involved in the CSE-induced attenuation. Administration of GDF11 ameliorated the stress-induced cellular senescence and production of ROS and inflammatory mediators in the lung cells. To further explore the in vivo effect of GDF11 on cellular senescence, we demonstrated that treatment with GDF11 attenuated elastase-accelerated cellular senescence in an emphysema mimetic model. Our data suggest that GDF11 may have an anti-senescent effect against stress-induced cellular senescence both in vitro and in vivo.
Previous reports have demonstrated that cellular senescence is accelerated not only in lungs but also in the whole body in patients with COPD.6–10 Although the precise mechanisms of cellular senescence in COPD have not been elucidated yet, oxidative stress and chronic inflammation are believed to play a key role in the observed senescence.26 Excessive oxidative stress reportedly occurs in patients with COPD,27 ,28 and it induces DNA damage and premature senescence.12 ,29 Alternatively, apoptosis occurs in the lung cells of patients with COPD, and this loss of lung cells might promote cell replication, resulting in replicative senescence.30 Meanwhile, there are few reports concerning the circulating anti-ageing factors in COPD.31 ,32 Recent reports in mouse models showed that GDF11 may have an anti-ageing role in other organs, including skeletal muscle,18 heart20 and brain.19 In this study, we revealed for the first time that GDF11 was significantly decreased in patients with COPD and that the levels of GDF11 were significantly correlated with disease severity. In vitro experiments showed that CSE attenuated GDF11 production, and treatment with NAC blocked this attenuation, suggesting that oxidative stress might be involved in the production GDF11.
We demonstrated that treatment with GDF11 inhibited the production and activation of senescence markers including p16, p53 and SA-β-gal in the lungs of an animal model. Interestingly, administration of GDF11 partly but significantly inhibited the elastase-induced enlargement of air spaces in this model. In the elastase model, oxidative stress33 and digestion of extracellular matrix proteins34 ,35 have been reported, and stress-induced senescence is observed in the lung cells.36 We also showed that treatment with GDF11 ameliorated stress-induced cellular senescence, ROS generation, the overproduction of MMPs, the release of IL-8 and the impairment of the tissue repair function in vitro. Thus, these effects of GDF11 may contribute to the anti-senescent effect and inhibition of alveolar destruction observed in the animal model.
There are limitations in this study. First, the sample size of each cohort was relatively small. To measure the levels of GDF11 in larger cohorts, a measuring system better than western blotting needs to be established. Second, in vitro experiments using CSE are not clearly relevant to the pathogenesis of COPD. However, in vitro CSE experiments have been widely used to explore signal transduction events and biological responses caused by smoking-related oxidative stress in the lungs of patients with COPD. Therefore, we used this model to assess stress-induced cellular senescence. Third, we used the elastase-induced emphysema mimetic animal model, not the cigarette smoke-induced emphysema model. Cigarette smoke is a better inducer of emphysema, and the cigarette smoke model reflects a more similar pathophysiology to that of human COPD. However, the elastase-induced emphysema mouse model is also widely used as an emphysema mimetic model and is known to cause stress-induced cellular senescence in the emphysematous lesions.36 Because we attempted to investigate the effects of GDF11 on cellular senescence in lungs, we used this model in the current study.
In conclusion, we demonstrated that the circulating and lung GDF11 decreased in COPD and that the levels of GDF11 were associated with pulmonary function data and comorbidities. GDF11 exhibited anti-senescent effects in lung cells both in vitro and in vivo. These results suggest that GDF11 may be involved in the senescence observed in COPD and may have potential for revealing the mechanisms of cellular senescence in COPD.
Acknowledgments
We thank Mr Brent Bell for reading this manuscript. Our manuscript has been proofread and edited by NPG Language Editing.
References
Footnotes
Contributors KO: cell culture, biochemical studies, immunohistochemical analysis, interpretation of results. HS: design of the study, interpretation of results, technical advice, writing of the manuscript. MY: design of the study, quantification of mRNAs, technical advice, interpretation of results. AK, NF, SY, YH, TO and TT: technical advice, interpretation of results. RT, TN, KS, YK, SK, MY, MM, YH, YO and SS: recruitment of patients, informed consent of patients. ST: preparation of rat tail tendon collagen, technical advice. MI: design of the study, interpretation of results, writing of the manuscript.
Funding This study was supported by grants from the Japan Society for the Promotion of Science (grant number: #26293195, #16H05307, #16K15453) and the Japan Agency for Medical Research and Development (grant number: #16ek0410018h0002, #16ek0410036h001, #17ek0410036h0002).
Competing interests None declared.
Patient consent Obtained.
Ethics approval Ethics committee of Tohoku University Graduate School of Medicine, Tohoku Rosai Hospital, and Japanese Red Cross Ishinomaki Hospital
Provenance and peer review Not commissioned; externally peer reviewed.