Article Text

Abstract
Background The death receptor ligand tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) shows considerable clinical promise as a therapeutic agent. TRAIL induces leukocyte apoptosis, reducing acute inflammatory responses in the lung. It is not known whether TRAIL modifies chronic lung injury or whether TRAIL has a role in human idiopathic pulmonary fibrosis (IPF). We therefore explored the capacity of TRAIL to modify chronic inflammatory lung injury and studied TRAIL expression in patients with IPF.
Methods TRAIL−/− and wild-type mice were instilled with bleomycin and inflammation assessed at various time points by bronchoalveolar lavage and histology. Collagen deposition was measured by tissue hydroxyproline content. TRAIL expression in human IPF lung samples was assessed by immunohistochemistry and peripheral blood TRAIL measured by ELISA.
Results TRAIL−/− mice had an exaggerated delayed inflammatory response to bleomycin, with increased neutrophil numbers (mean 3.19±0.8 wild type vs 11.5±5.4×104 TRAIL−/−, p<0.0001), reduced neutrophil apoptosis (5.42±1.6% wild type vs 2.47±0.5% TRAIL−/−, p=0.0003) and increased collagen (3.45±0.2 wild type vs 5.8±1.3 mg TRAIL−/−, p=0.005). Immunohistochemical analysis showed induction of TRAIL in bleomycin-treated wild-type mice. Patients with IPF demonstrated lower levels of TRAIL expression than in control lung biopsies and their serum levels of TRAIL were significantly lower compared with matched controls (38.1±9.6 controls vs 32.3±7.2 pg/ml patients with IPF, p=0.002).
Conclusion These data suggest TRAIL may exert beneficial, anti-inflammatory actions in chronic pulmonary inflammation in murine models and that these mechanisms may be compromised in human IPF.
- Neutrophil
- apoptosis
- death ligand
- bleomycin
- cytokine biology
- innate immunity
- interstitial fibrosis
- primary pulmonary hypertension
- bacterial infection
- lymphocyte biology
- macrophage biology
- pneumonia
- respiratory infection
- neutrophil biology
- histology/cytology
- COPD mechanisms
- allergic lung disease
- asthma
- eosinophil biology
- allergic alveolitis
- lung proteases
- sarcoidosis
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/2.0/ and http://creativecommons.org/licenses/by-nc/2.0/legalcode.
Statistics from Altmetric.com
- Neutrophil
- apoptosis
- death ligand
- bleomycin
- cytokine biology
- innate immunity
- interstitial fibrosis
- primary pulmonary hypertension
- bacterial infection
- lymphocyte biology
- macrophage biology
- pneumonia
- respiratory infection
- neutrophil biology
- histology/cytology
- COPD mechanisms
- allergic lung disease
- asthma
- eosinophil biology
- allergic alveolitis
- lung proteases
- sarcoidosis
Key messages
What is the key question?
Does the cell death-inducing molecule tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) regulate the severity of chronic lung inflammation and does it have a role in human idiopathic pulmonary fibrosis (IPF)?
What is the bottom line?
TRAIL-deficient mice have worse lung inflammation and more lung collagen than wild-type mice and patients with IPF show lower levels of TRAIL than healthy controls.
Why read on?
TRAIL is a potential biomarker of IPF progression and correction of TRAIL deficiency might have clinical utility in treatment of IPF.
Introduction
Idiopathic pulmonary fibrosis (IPF) is a chronic scarring disease of the lung of uncertain pathogenesis and to date no therapy has convincingly improved survival or modified clinical course.1 IPF is a heterogeneous condition both in terms of clinical course and pathological appearance. This suggests it may be initiated by different forms of lung injury and alveolar epithelial injury followed by aberrant repair is currently regarded as a central pathogenic mechanism.2 The role of inflammation, and particularly of neutrophils, in IPF is controversial but neutrophilic alveolitis is a frequent feature of the disease.3 ,4 Moreover, higher neutrophil counts in bronchoalveolar lavage (BAL) are associated with more rapid disease progression and with worse lung function.5 ,6 Neutrophilic inflammation is also a component of experimental models of IPF, including bleomycin-induced lung injury in mice.7 ,8
Tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) has important roles in regulating cell survival, particularly of immune cell populations. TRAIL is a type II membrane protein that is principally expressed by leucocytes, including monocytes, macrophages, lymphoid cells and neutrophils.9 In humans, TRAIL interacts with four membrane receptors belonging to the TNF receptor family. TRAIL receptor 1 (TRAIL-R1) and TRAIL-R2 have cytoplasmic death domains, and can activate caspases and nuclear factor κB (NFκB). The receptors TRAIL-R3 and TRAIL-R4 have truncated death domains and do not activate caspases; they are usually described as decoy receptors but may activate NFκB. A soluble decoy receptor for TRAIL, osteoprotegerin (OPG), is also described. Mice have a single TRAIL death receptor, which shares 79% sequence homology with human TRAIL-R2, together with two decoy receptors.10 The clinical promise of TRAIL as an agent with which to manipulate apoptosis is emphasised by its use in phase I and II studies in cancer therapy.11
We previously showed exogenous TRAIL can accelerate apoptosis of neutrophils, a key cellular process for resolving inflammation in vivo.12 ,13 TRAIL is implicated in the elimination of senescent neutrophils from the human circulation.14 Our recent data in a murine model of acute lung injury show TRAIL regulates neutrophil lifespan at sites of inflammation.15 There is little data on roles of TRAIL in chronic inflammation. TRAIL can, however, modify fibrosis, since TRAIL treatment of mice has been shown to promote resolution of hepatic fibrosis and, importantly, there is evidence TRAIL may directly induce apoptosis of primary lung fibroblasts.16 ,17
Because of these dual effects upon neutrophils and fibroblasts we hypothesised TRAIL may regulate chronic inflammation in the lung, modifying both the inflammatory and fibroproliferative components of this condition, and play a role in human IPF.
Materials and methods
Animals
TRAIL−/− mice on a C57/BL6 background were a kind gift from Amgen (Seattle, Washington, USA). All procedures were approved by the University of Sheffield Ethics Committee and were performed in accordance with the Home Office Animal (Scientific Procedures) Act 1986.
Human subjects
All lung tissue was supplied by the Cambridge Tissue Bank with approval from the Papworth Hospital Ethics Committee, with further details in the online supplement. Serum samples were collected from 31 patients with IPF. Controls were age and sex matched to patients with IPF and were recruited via the patient's family physician who confirmed they did not have a diagnosis of interstitial lung disease. Clinical data, including pulmonary function tests, are provided in the online supplement. These studies were performed with the approval of the South Sheffield Research Ethics Committee.
Bleomycin model of lung injury
Female mice were used between 8 and 12 weeks of age, in accordance with the UK Home Office Animals (Scientific Procedures) Act, 1986. Clinical-grade bleomycin (Nippon Kayaku, Slough, UK) 1 mg/kg body weight in 30 μl of saline, or an equal volume of saline as a control, was administered by single intratracheal injection into a surgically exposed trachea under anaesthesia.
Bronchoalveolar lavage
Following BAL a haemocytometer total cell count was performed and differential cell counts calculated from cytospin preparations (Cytospin 3; Thermo Shandon, Runcorn, UK) stained with Diff-Quik and assessed by blinded reviewers.
Assessment of neutrophil apoptosis
The proportion of apoptotic neutrophils was determined by blinded reviewers, counting duplicate cytospins stained by Diff-Quik (>300 cells per slide).
Measurement of total lung collagen
Total lung collagen was calculated by measuring hydroxyproline content in aliquots of pulverised lung. Details are given in the online supplement.
Preparation of lung tissue for histological studies
Unlavaged lungs were insufflated with 4% paraformaldehyde in phosphate-buffered saline (PBS) at a pressure of 20 cm H2O, followed by removal of the heart and inflated lungs en bloc and immersion for 4 h in fresh fixative. Subsequently lungs were transferred into 15% sucrose in PBS and left overnight at 4°C, before transfer to 70% ethanol.
Masson's trichrome staining
Unlavaged lungs were formalin fixed, processed and embedded in paraffin wax blocks. Sections of lung tissue were stained as described in the online data supplement.
Immunohistochemistry
Staining with specific antibodies and TUNEL staining was performed as detailed in the online data supplement.
ELISAs
ELISAs for TRAIL and OPG were from R&D Systems (Abingdon, UK) and performed according to the manufacturer's instructions.
Statistical analysis
Unless otherwise stated, results were analysed using a one-way or two-way analysis of variance as appropriate, followed by a Bonferroni's post-test for multiple comparisons. Patient and control data for serum TRAIL and OPG levels were compared using two-tail paired t-tests. Correlations between serum TRAIL levels and total diffusing capacity of the lung for carbon monoxide (TLco) or survival were assessed using Pearson's correlation coefficient. Results were considered significant if p<0.05.
Results
Bleomycin-mediated pulmonary inflammation is enhanced in TRAIL-deficient mice
The composition of BAL was assessed at time points up to 23 days following bleomycin administration. Bleomycin instillation resulted in inflammatory cell recruitment into the lungs of challenged mice. BAL fluid from control mice (PBS instilled) consisted overwhelmingly of alveolar macrophages and contained <0.1×105 neutrophils at all time points in wild-type and TRAIL−/− strains (data not shown). There was an enhanced inflammatory response in TRAIL−/− compared with wild-type mice, with significantly increased total and percentage neutrophil counts at day 7 following bleomycin instillation (mean 3.19±0.8×104 in wild-type vs 11.5±5.4×104 in TRAIL−/− mice, p<0.0001; mean 2.87±1.0% in wild-type vs 8.25±1.3% neutrophils in TRAIL−/− mice, p<0.0001) (figure 1A,B). Macrophages (as total or percentage count) were not significantly different between the groups of mice (figure 1C,D) but TRAIL−/− mice also had a significant increase in total lymphocytes at day 7 when compared with wild-type mice (mean 7.32±2.8×104 in wild-type vs 14.6±4.8×104 in TRAIL−/− mice, p=0.035) (figure 1E). To characterise the neutrophilic inflammation in more detail, we performed additional time points at day 5 and 10 and confirmed peak neutrophil counts were detected at day 7 (data not shown). Lymphocyte numbers also peaked at day 7 and were predominantly CD3+ as opposed to CD19+ cells (data not shown).

Differential leucocyte counts in bronchoalveolar lavage (BAL) fluid following intratracheal administration of bleomycin in wild-type and tumour necrosis factor-related apoptosis-inducing ligand (TRAIL)-deficient mice. Differential leucocyte counts were obtained from cytocentrifuge preparations of BAL fluid lavaged from the lungs at time points up to 23 days. (A,D) Total and percentage neutrophil counts (mean ± SEM of at least five independent experiments) for each time point were obtained by multiplying the differential count by the total leucocyte number obtained from haemocytometer counts for wild-type (solid line) and TRAIL−/− mice (dashed line). (B,E) Total and percentage macrophage counts for each time point. (C,F) Total and percentage lymphocyte counts for each time point. Both absolute (***p<0.0001) and percentage (*p<0.0001) neutrophil counts were increased in TRAIL−/− mice compared with wild-type mice at day 7, as were total lymphocyte counts (p=0.035).
TRAIL-deficient mice have reduced apoptosis following bleomycin-induced lung injury
TUNEL staining of day 23 lung sections from both wild-type and TRAIL−/− mice revealed significantly fewer apoptotic cells in TRAIL−/− mice (mean 46.4±25.1 cells/high-power field (HPF) in wild-type vs 8.90±6.30 cells/HPF in TRAIL−/− mice, p<0.0001) (figure 2). The majority of apoptotic events seen in wild-type mice were in areas with numerous influxing inflammatory cells (figure 2A). Very few such events were seen in TRAIL−/− mice, despite the presence of inflammatory infiltrates (figure 2B). That differences in rates of apoptosis in the inflammatory cell infiltrate explained the differences in numbers of TUNEL-positive events was apparent by analysing the numbers of apoptotic cells in BAL (figure 3). The proportion of the neutrophil BAL population after bleomycin instillation that had the microscopic appearances of apoptosis was <2% in BAL from both wild-type and TRAIL−/− mice at day 3. This increased as neutrophil numbers peaked at day 7 and then decreased (figure 3A). At later time points there was a significantly lower proportion of apoptotic neutrophils in the TRAIL−/− mice, for example, mean 5.42±1.6% in wild-type vs 2.47±0.5% in TRAIL−/− mice at day 7, p=0.0003. This suggested reduced apoptosis was contributing to the greater neutrophil numbers in BAL of TRAIL−/− mice. Despite the greater neutrophil numbers in TRAIL−/− mice, these mice also showed fewer absolute numbers of apoptotic neutrophils than their wild-type counterparts (figure 3B).

TUNEL-positive apoptotic events are reduced in tumour necrosis factor-related apoptosis-inducing ligand (TRAIL)-deficient mice. Representative TUNEL-stained lung tissue sections from wild-type (A) and TRAIL−/− mice (B) at day 23 after bleomycin instillation. Apoptotic events are visible as brown-stained cells. (C) Numbers of TUNEL-positive cells per high-power field (HPF) were significantly higher in wild-type than in TRAIL−/− mice (**p<0.0001, n=3 mice per group, counting 10 random fields per section).

Neutrophil apoptosis in bronchoalveolar lavage (BAL) fluid following intratracheal administration of bleomycin in wild-type and tumour necrosis factor-related apoptosis-inducing ligand (TRAIL)-deficient mice. The percentage of BAL fluid neutrophils with morphological appearances of apoptosis was assessed at day 3, 7, 16 and 23 after bleomycin instillation. (A) At time points up to day 7, <4% of neutrophils were apoptotic in both wild-type (solid line) and TRAIL−/− mice (dashed line). At days 7 and 16, the percentage of apoptotic neutrophils was increased in both populations, but there was a significantly lower proportion of apoptotic neutrophils in TRAIL−/− mice (**p=0.0003 at day 7 and *p=0.012 at day 16). (B) Total numbers of apoptotic neutrophils were also significantly reduced in TRAIL−/− mice (*p=0.0316 at day 7).
TRAIL-deficient mice have increased total lung collagen
The effect of TRAIL deficiency on total lung collagen accumulation, 16 days after intratracheal administration of bleomycin or saline, is shown in figure 4. This time point has previously identified significant differences in fibro-proliferative responses to bleomycin.18 TRAIL−/− and wild-type mice demonstrated no significant difference in lung collagen levels after saline challenge. Lung collagen accumulation was significantly increased in bleomycin-instilled wild-type mice compared with saline control (mean 2.08±0.60 mg with saline treatment vs 3.46±0.30 mg with bleomycin, p=0.0033) and was of at least equivalent magnitude to previous studies using the same experimental protocol.18 In comparison to wild-type mice there was a significantly greater increase in collagen accumulation following bleomycin treatment in TRAIL−/− mice (mean 3.46±0.3 mg in wild-type vs 5.8±1.3 mg in TRAIL−/− mice, p=0.005).

Lung collagen accumulation in response to bleomycin is increased in tumour necrosis factor-related apoptosis-inducing ligand (TRAIL)-deficient mice. (A–C) Representative lung tissue sections stained with Masson's Trichrome from wild-type mice at day 7 and 23 after saline and bleomycin instillations. (A) Lung architecture was normal in TRAIL−/− mice given saline. (B) and (C) Extensive patchy fibrotic foci with increased deposition of collagen were seen in wild-type mice given bleomycin at day 7 and 23, respectively. (D) Total lung collagen as measured by reverse phase high-performance liquid chromatography quantitation of lung hydroxyproline in acid hydrolysates of pulverised lung. Data represent the mean ± SEM of values obtained in groups of five mice. Wild-type and TRAIL−/− mice instilled with saline had similar collagen levels in the lungs. Wild-type mice instilled with bleomycin had a significant increase in collagen compared with the saline control (*p=0.0033). TRAIL−/− mice had a significant increase in collagen compared with wild-type mice instilled with bleomycin (**p=0.005) and with saline controls (**p=0.0004).
TRAIL immunolocalisation following bleomycin-induced lung injury
Immunohistochemical examination of TRAIL expression in the lung was performed at key time points in wild-type mice, with TRAIL−/− mice acting as controls. In wild-type animals receiving saline intratracheally, TRAIL staining of the bronchial epithelium was observed, together with some staining in alveolar type 2 pneumocytes, in keeping with previous studies.19 Following bleomycin administration, no specific TRAIL immunostaining was detected in TRAIL−/− mice (figure 5A), while in wild-type mice staining was seen in the bronchial and alveolar epithelium and also in well defined groups of cells adjacent to the bronchial epithelium and in cells in the alveolar air spaces (figure 5B–D). These positively staining cells were identified as bronchus-associated lymphoid tissue and alveolar macrophages, as illustrated by staining with anti-CD3 and anti-galectin-3 antibodies respectively (online supplement).

Tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) expression localises to bronchus associated lymphoid tissue (BALT) and alveolar macrophages. Lung sections 16 days after bleomycin instillation were immunohistochemically stained for TRAIL. (A) Lung section demonstrating the presence of alveolar macrophages and BALT tissue and absence of specific TRAIL immunostaining in a TRAIL−/− mouse. (B) and (C) Lung sections (representative of sections from five mice) demonstrating TRAIL expression localised to epithelium and BALT in a wild-type mouse. (D) Lung section demonstrating TRAIL expressing alveolar macrophages in a wild-type mouse.
Immunohistochemical expression of TRAIL in human IPF
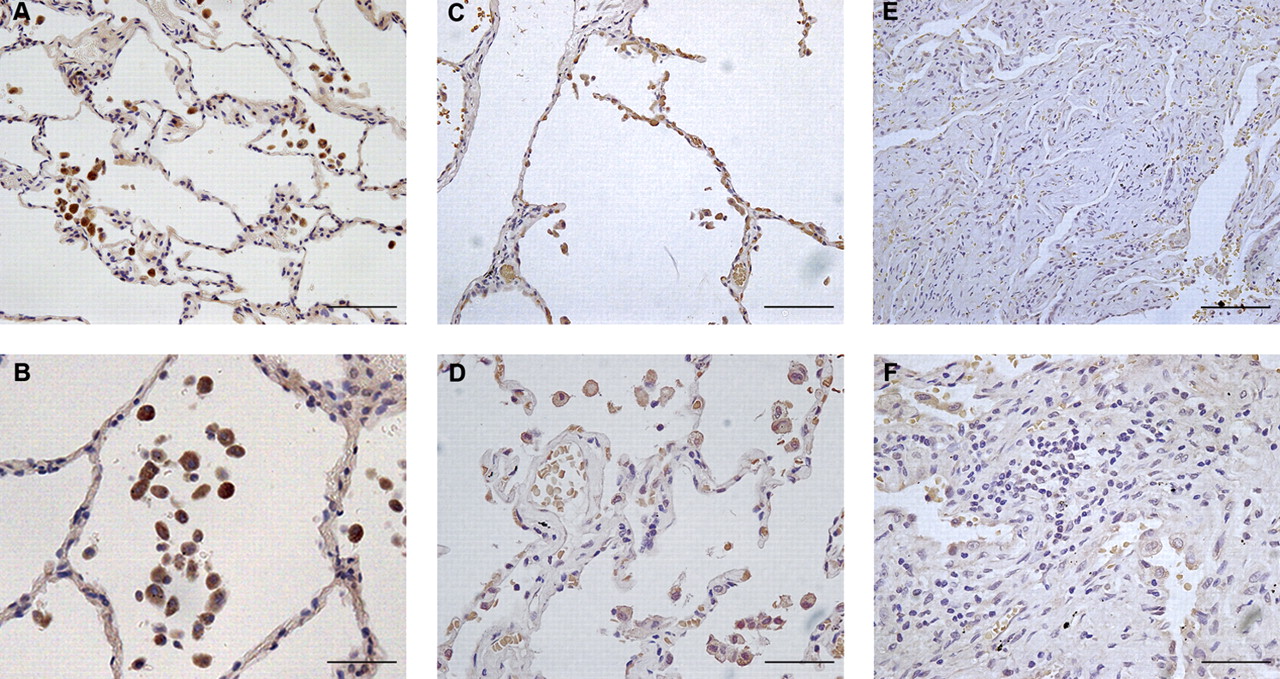
To examine the potential role of TRAIL in human fibrotic lung disease, we examined TRAIL expression in human lung tissue specimens by immunohistochemistry. In lung sections from control patients there was positive staining for TRAIL associated with resident alveolar macrophages and in the epithelium (figure 6A,B), in keeping with findings in unchallenged wild-type mice (figure 5 and Weckmann et al19). In lung tissue obtained from patients with established IPF, TRAIL staining was detected in areas of relatively normal lung, both in alveolar macrophages and epithelial cells (figure 6C,D). In areas of dense fibrosis, however, there was little staining either of alveolar macrophages or of lymphoid tissue with variable but generally low-level staining of the respiratory epithelium (figure 6E,F).

Immunohistochemical localisation of tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) in human lung specimens. Immunostaining for TRAIL on representative lung tissue sections from human lung material. (A) and (B) Intense immunostaining for TRAIL in control lung was associated with macrophages and alveolar epithelium. Representative images are shown from lung sections of three individuals. (C–F) Analysis of lung tissue from patients with idiopathic pulmonary fibrosis. Representative images are shown from lung sections of five individuals. (C, D) Areas of lung with relatively normal architecture showed TRAIL staining similar to that from control lung samples whereas matched samples from fibrotic areas of the same individuals (E, F) demonstrated almost no positive immunostaining for TRAIL. Scale bars for panels (A), (C) and (E) represent 100 μm and for (B), (D) and (F) 50 μm.
Serum TRAIL is reduced in patients with IPF compared with controls
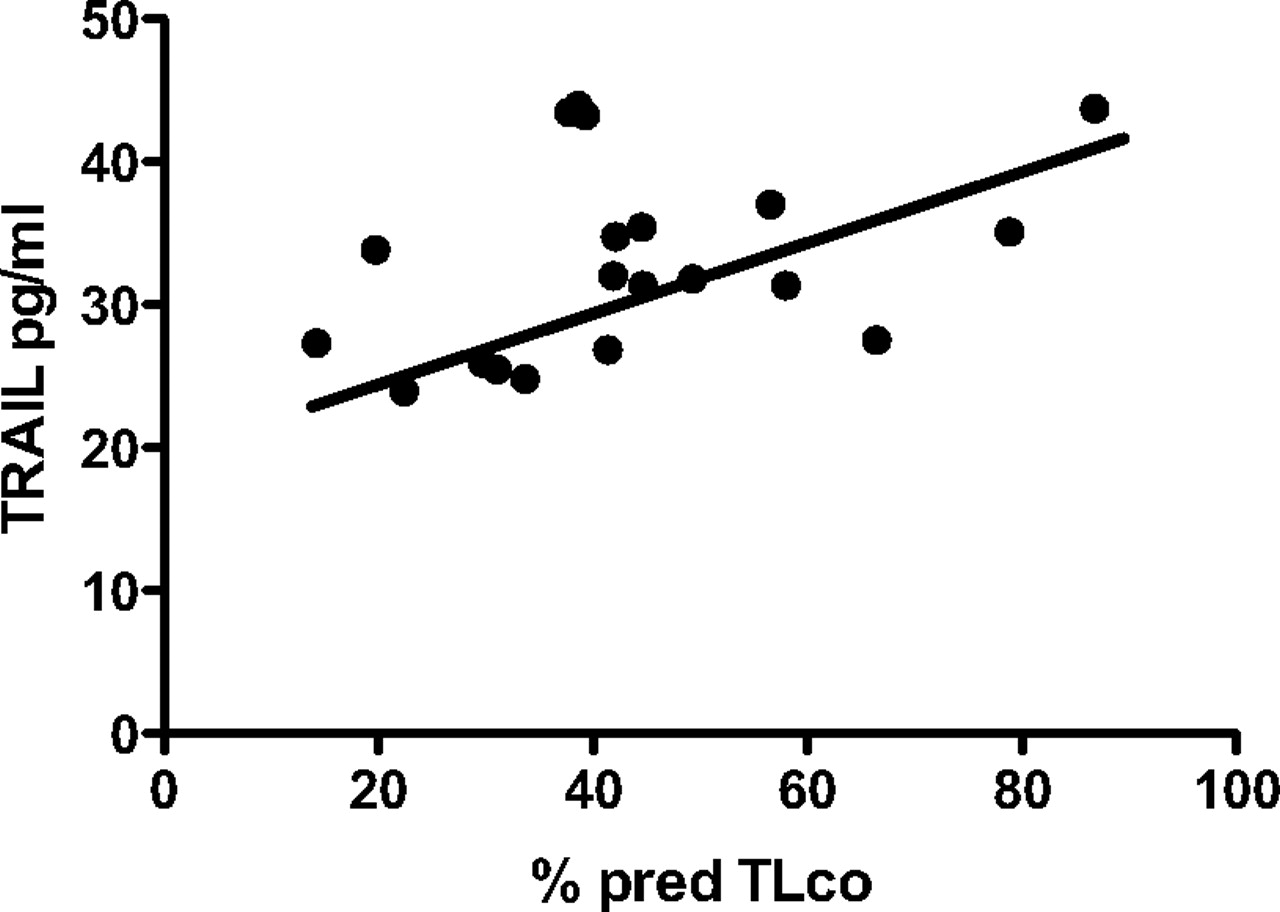
The soluble form of TRAIL is normally present in serum and can be detected by ELISA, as can its soluble receptor, OPG.9 ,20 Absolute serum levels of TRAIL were reduced in patients with IPF relative to their matched controls (p=0.002, figure 7A) although OPG levels were not significantly altered (p=0.230, figure 7B). The ratio of OPG:TRAIL was significantly increased (p=0.033, figure 7C). Within the cohort of patients with IPF (online supplement), serum TRAIL levels were significantly correlated with predicted TLco (p=0.046, figure 8), although the magnitude of the correlation was not large. There was no relationship identified between forced vital capacity (% predicted) and serum TRAIL levels and no correlation of serum OPG levels with any of these parameters (data not shown).

Serum levels of tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) and its soluble receptor osteoprotegerin (OPG) in patients with idiopathic pulmonary fibrosis (IPF) and matched controls. Serum levels of (A) TRAIL and (B) OPG were measured by ELISA in patients with IPF and matched controls. Data was also expressed as (C) the ratio of OPG to TRAIL. TRAIL levels were significantly reduced in patients with IPF (**p=0.002) and the OPG:TRAIL ratio correspondingly increased (*p=0.033).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Serum levels of tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) correlate with TLco values. Serum TRAIL, measured at time of clinical presentation with idiopathic pulmonary fibrosis (IPF), was significantly correlated with TLco (% predicted) at time of diagnosis (r2=0.149, p=0.046).
Discussion
Neutrophils are thought to play an important role in the pathogenesis of pulmonary fibrosis. Accumulation of neutrophils and increased release of neutrophil chemotactic factors are observed in patients with a variety of causes of pulmonary fibrosis, including IPF.4 ,21 Neutrophil-dependent, T-cell independent mechanisms make an important contribution to collagen deposition in several murine models of pulmonary fibrosis while factors which reduce neutrophil accumulation and production of neutrophil chemotactic factors reduce collagen deposition.22–25 Thus accumulation of neutrophils, which can perpetuate pulmonary inflammation through the release of matrix metalloproteinases and serine proteases, can enhance pulmonary fibrosis.8 ,26 Conversely, compounds such as cyclin-dependant kinase inhibitors that accelerate neutrophil apoptosis reduce lung fibrosis.13
We examined the role of TRAIL in chronic inflammation in the lung using the well established bleomycin model. This model reproduces many of the histological features of IPF and is associated with recruitment of inflammatory cells, including neutrophils, into the injured lung during the first 7 days, with fibrotic change and deposition of matrix occurring up to day 35.27 Assessment of the inflammatory responses in wild-type and TRAIL−/− mice revealed TRAIL deficiency resulting in increased BAL neutrophilia and reduced neutrophil apoptosis. The total number of neutrophils present in BAL in the bleomycin-treated wild-type mice was in keeping with previous data.8 The recruitment of neutrophils to the lung cannot be directly measured in murine models but the two strains showed equivalent cell numbers at day 3, which subsequently diverged, suggesting no difference in rates of initial recruitment. The data could reflect ongoing neutrophil recruitment at later time points in TRAIL−/− mice but also suggest a contribution of delayed apoptosis and clearance of these cells. Delay of apoptosis is supported both by the finding of reduced TUNEL-positive events in inflammatory cells (figure 2) and of reduced numbers of apoptotic neutrophils in BAL (figure 3) and by our previous data in lipopolysaccharide (LPS)-mediated lung injury.15
TRAIL-deficient mice showed biochemical evidence of increased collagen deposition. In both the bleomycin model and human pulmonary fibrosis tissue, there is increased deposition of collagen and other molecules of the extracellular matrix by lung fibroblasts.8 Previous work by Yurovsky showed TRAIL induces apoptosis of primary lung fibroblasts derived from patients with IPF.17 The increased lung collagen we observed in the absence of TRAIL could, therefore, result from the lack of its pro-apoptotic effects on pulmonary fibroblasts, and/or the increased inflammatory drive to fibroproliferation consequent upon increased numbers of neutrophils.
TRAIL is widely expressed on leucocyte populations and, in the normal human lung, TRAIL expression has been detected on alveolar septa, bronchial epithelium and vascular endothelium.9 ,28 In bleomycin-treated wild-type mice, TRAIL was also detected in a population of CD3-positive, peribronchial inflammatory cells with the appearances of bronchus associated lymphoid tissue and in galectin-3-positive air-space cells with the characteristic appearance of alveolar macrophages. These findings were in keeping with BAL analysis in which macrophages and lymphocytes were found to express TRAIL by flow cytometry (data not shown). Macrophage expression of death receptor ligands has been shown to regulate apoptosis induction in a variety of inflammatory cells. A role for macrophage Fas ligand (FasL) induction of target cell killing has been described and for TNFα acting in concert with integrins in the macrophage induction of apoptosis in neutrophils.29 ,30 We did not detect neutrophil expression of TRAIL in our experiments, in keeping with other recent data.31 The distribution of TRAIL expression in wild-type mice following bleomycin lung injury, with ongoing expression on epithelial cells but upregulated expression on inflammatory cells, is very different from that in control samples and in LPS-mediated lung injury or allergen challenge models in which TRAIL expression is predominantly on the airway epithelium.15 ,19 In a recently described model of influenza-mediated inflammatory lung disease, TRAIL was expressed on alveolar macrophages and also on a small proportion of natural killer cells in BAL.31 Our data suggest that macrophage-expressed and possibly lymphocyte-expressed TRAIL could play an important role in regulating the inflammatory response in chronic pulmonary inflammation.
TRAIL is not the only death receptor ligand shown to induce apoptosis of inflammatory leucocytes. FasL and TNFα have well characterised roles in acceleration of neutrophil apoptosis, but each also has significant proinflammatory effects. TRAIL mostly induces apoptosis of transformed cells, although effects on some primary cell types are described.9 We previously showed TRAIL had no chemotactic effects in human neutrophils in vitro but did accelerate neutrophil apoptosis via ligation of the TRAIL-R2 receptor.12 The TRAIL system has since been implicated in the elimination of senescent circulating neutrophils, suggesting a role in physiologic regulation of neutrophil lifespan. In addition, we recently found TRAIL-deficient mice have an enhanced inflammatory response to LPS in vivo.14 ,15 The majority of primary cell types, although not all, are resistant to TRAIL, in part due to expression of cell surface decoy receptors TRAIL-R3 and TRAIL-R4, reducing the likelihood of unwanted cytotoxicity in other cell types with TRAIL treatment in vivo.9 In contrast, FasL induces apoptosis of bronchial epithelial cells and pulmonary overexpression of TNFα induces chronic pulmonary inflammation and fibrosis.32 ,33
Recent work in a murine model of asthma showed increased pulmonary epithelial expression of TRAIL and that inhibition of TRAIL function attenuated allergic inflammation.19 In contrast, we found the absence of TRAIL is associated with enhanced inflammation in the lung and that TRAIL is expressed on macrophage and lymphoid cells. Moreover, absence of TRAIL results in more severe inflammation and fibrosis, suggesting this ligand may have a role in ameliorating non-allergic inflammation. In keeping with this, recent studies of TRAIL-R-deficient mice showed development of severe pneumonitis following ionising radiation that was absent from wild-type mice.34
We show there may be a relative paucity of TRAIL expression in the fibrotic regions of interstitial lung disease in man. Lung samples from patients with IPF showed low levels of TRAIL expression in densely fibrosed areas, with relative preservation of TRAIL expression in more histologically normal areas. Circulating levels of soluble TRAIL were lower in patients with IPF than in controls, whereas levels of the soluble receptor, OPG, were unchanged. The magnitude of difference seen in TRAIL levels, although relatively small, was comparable to other studies using the same ELISA and equivalent to the changes seen in patients with atherosclerosis compared with healthy controls, where lower levels of TRAIL have been shown to be of prognostic significance.20 ,35
The biological roles of TRAIL in human IPF clearly require further exploration. In the bleomycin model, deficiency of TRAIL leads to increased inflammation and collagen deposition, suggesting TRAIL expression by leucocytes and/or tissue cells acts to limit these features of disease and thus might act as a disease modifier, influencing disease progression. TRAIL also has possible utility as a biomarker of disease progression in IPF, which could be addressed in longitudinal studies. Whether TRAIL has a role in susceptibility to disease is unknown, although functional genetic polymorphisms of TRAIL and its receptors exist and could be studied in patients with IPF and control populations. It is also conceivable that treatment with exogenous TRAIL might have a beneficial anti-inflammatory and antifibrotic effect in IPF. Forms of recombinant TRAIL and TRAIL-R2 agonistic antibodies are in clinical trials in a range of malignant diseases and could be considered for a non-malignant disease, such as IPF, with an equally poor prognosis.11
Acknowledgments
We thank Amgen Inc (Seattle, Washington, USA) and Dr Mark Smyth for the gift of TRAIL-deficient mice.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
Footnotes
Funding This work was funded by a Wellcome Trust Clinical Research Training Fellowship to EMcG (075776), by the NIHR Cardiovascular Biomedical Research Unit Sheffield (IS and MKBW) and by generous donations in memory of Mrs Eileen Hawley, Mrs Susan Utley and Mr Derrick Woolley. AL holds an MRC Career Development Award Fellowship (G0800318), SRW is a Wellcome Clinician Scientist Fellow (078244), SAR is an MRC Senior Clinical Fellow (G0701932) and DHD a Wellcome Trust Senior Clinical Fellow (076945).
Competing interests None.
Ethics approval Ethics approval was provided by Papworth Hospital Ethics Committee and South Sheffield Research Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.