Article Text

Abstract
Background Idiopathic pulmonary fibrosis (IPF) has a poor prognosis and limited responsiveness to available treatments. It is characterised by epithelial cell injury, fibroblast activation and proliferation and extracellular matrix deposition. Serotonin (5-hydroxytryptamine; 5-HT) induces fibroblast proliferation via the 5-HTR2A and 5-HTR2B receptors, but its pathophysiological role in IPF remains unclear. A study was undertaken to determine the expression of 5-HT receptors in IPF and experimental lung fibrosis and to investigate the effects of therapeutic inhibition of 5-HTR2A/B signalling on lung fibrosis in vivo and in vitro.
Methods and results Quantitative RT-PCR showed that the expression of 5-HTR1A/B and 5-HTR2B was significantly increased in the lungs of patients with IPF (n=12) and in those with non-specific interstitial pneumonia (NSIP, n=6) compared with transplant donors (n=12). The expression of 5-HTR2A was increased specifically in IPF lungs but not in NSIP lungs. While 5-HTR2A protein largely localised to fibroblasts, 5-HTR2B localised to the epithelium. To assess the effects of 5HTR2A/B inhibition on fibrogenesis in vivo, mice were subjected to bleomycin-induced lung fibrosis and treated with the 5-HTR2A/B antagonist terguride (or vehicle) in a therapeutic approach (days 14–28 after bleomycin). Terguride-treated mice had significantly improved lung function and histology and decreased collagen content compared with vehicle-treated mice. Functional in vitro studies showed that terguride is a potent inhibitor of transforming growth factor β1- or WNT3a-induced collagen production.
Conclusion The studies revealed an increased expression of 5-HTR2A specifically in IPF. Blockade of 5-HTR2A/B signalling by terguride reversed lung fibrosis and is thus a promising therapeutic approach for IPF.
- Serotonin
- collagen
- tissue remodelling
- Interstitial fibrosis
Statistics from Altmetric.com
Introduction
Idiopathic pulmonary fibrosis (IPF) is a progressive and fatal lung disease of unknown origin.1 2 It is characterised by alveolar epithelial cell damage, increased deposition of extracellular matrix (ECM) in the lung interstitium, enhanced fibroblast/myofibroblast proliferation and activation and, ultimately, distortion of normal lung architecture and loss of respiratory function.3 4 The hallmark lesions of IPF are fibroblast foci, sites featuring activated myofibroblasts mainly responsible for altered ECM deposition in the lung interstitium.2
The pathogenesis of IPF includes inappropriate alveolar regeneration in response to repetitive epithelial injuries perpetuated by dysregulated wound repair in response to local inflammation.5 6 While the initial trigger of this disease is most likely represented by repetitive epithelial injuries of unknown origin, the interstitial fibroblast/myofibroblast represents the key effector cell responsible for ECM deposition in patients with IPF and in animal models of lung fibrosis such as the bleomycin-induced mouse model.3 7 8 It is currently unclear, however, which mediators and pathways are involved in disease initiation and progression. While soluble cytokines such as transforming growth factor (TGF) β1, interleukin (IL)-1β or WNT ligands have been assigned a pathogenic role in IPF and experimental models thereof,9–12 therapeutic options neutralising their activity have not been studied in appropriate clinical trials.
Serotonin (5-hydroxytryptamine, 5-HT) exhibits well-characterised regulatory functions in multiple physiological systems.13 14 It is involved in the regulation of cell migration, proliferation, cytokine production and vasoregulation.15 16 The multiple actions of 5-HT are mediated by seven different 5-HT receptor subtypes (5-HTR1–5-HTR7).17 Importantly, 5-HT and its receptors 5-HTR2A and 5-HTR2B have been implicated in fibrotic disorders such as retroperitoneal or pleural fibrosis, carcinoid heart disease and liver fibrosis.18–20 In addition, cumulative evidence suggests an important biological role for 5-HT signalling in the lung, supported by increased levels of 5-HT and active signalling thereof in asthma and pulmonary hypertension.21 22 In addition, recent evidence has suggested that the 5-HTR2A antagonists ketanserin and SB215505 reduced collagen mRNA and protein levels in the bleomycin-induced mouse model.23 Accordingly, this evidence demands a detailed characterisation of 5-HT receptor expression in IPF and analysis to determine whether 5-HT2A/B antagonists represent a therapeutic option in IPF, particularly since 5-HTR2 antagonists are already in clinical use for other diseases.24 25 Among these, terguride (1,1-diethyl-3-(6-methyl-8a-ergolinyl)urea) is a potent antagonist of 5-HTR2A and HTR2B, a partial dopaminergic agonist and an α2-adrenolytic agent.24 26 Terguride is clinically approved, well-tolerated and has been shown to be efficacious in the treatment of ovulation disorders due to hyperprolactinaemia.27
The expression and localisation of the 5-HT system in IPF lungs was investigated and compared with non-specific interstitial pneumonia (NSIP) and donor lungs. We performed a therapeutic approach in vivo using the mouse model of bleomycin-induced lung fibrosis to determine whether 5-HTR2A/B antagonism by terguride is a suitable therapeutic option for IPF.
Material and methods
Human tissue and cells
Lung tissues were obtained from 12 patients with IPF with a histological pattern of usual interstitial pneumonia (UIP) (4 women, 8 men; mean±SD age 57±9 years), 6 patients with non-specific interstitial pneumonia (NSIP, fibrotic phenotype) (4 women, 2 men; mean±SD age 55±9 years) and 12 control subjects (transplant organ donors; 6 women, 6 men; mean±SD age 40±10 years) after explantation. Detailed patient characteristics are shown in table S1 in the online supplement. Human ATII cells were isolated as previously described.28 The purity and viability of ATII cell preparations was consistently >90% and >95%, respectively. Freshly isolated human ATII cells were used for gene expression analysis.
Animals
Pathogen-free 6–8-week-old female C57BL/6N mice (Charles River Laboratories, Sulzfeld, Germany) weighing 18–22 g were used throughout the study. All experiments were performed in accordance with the guidelines of the ethics committee of University of Giessen School of Medicine and approved by the local authorities. Mice had ad libitum access to water and rodent laboratory chow. Bleomycin sulphate (Laboratories Almirall SA, Barcelona, Spain) was dissolved in sterile 0.9% saline and applied as a single dose of 1.5 U/kg body weight in a total volume of 80 μl by orotracheal application. Control mice received 80 μl saline only. Mice were treated with terguride 14 days after bleomycin application. As an internal control, a treatment arm with daily administration of 50 mg/kg imatinib was included in the study. Treatment with imatinib was started immediately following instillation of bleomycin. The detailed treatment scheme is shown in figure S4 in the online supplement.
Immunohistochemistry
Human lungs were submerged in 4% (w/v) paraformaldehyde after explantation and processed for paraffin embedding.29 30 Immunohistochemical staining of sections (3 μm) was performed according to the manufacturer's protocol (Histostain Plus Kit; Zymed Laboratories/Invitrogen, Carlsbad, California, USA). Antigen retrieval was performed in 6.5 mM sodium citrate pH 6.0 in a pressure cooker, after which endogenous peroxidase activity was quenched with 3% (v/v) hydrogen peroxide for 20 min. The following antibodies were used in the study: 5HTR2A (polyclonal, sc-50396, dilution 1:200; Santa Cruz Biotechnologies, California, USA); 5HTR2B (polyclonal, ab12926, dilution 1:200; Abcam, Cambridge, UK). An antibody against 5HTR2B (polyclonal, HPA012867, dilution 1:100, Sigma-Aldrich, St Louis, Missouri, USA) was also used and staining was performed using the Vulcan Fast Red Chromogen Kit (Zymed Laboratories/Invitrogen). For the negative control, an isotype-matched non-specific antibody was used (ab2410, dilution 1:200, Abcam).
Further experimental procedures
The detailed procedures for the determination of terguride levels, cell culture, quantitative RT-PCR, collagen assay, lung function and determination of the fibrotic score are provided in the online supplement.
Statistical analysis
All ΔCt values obtained from quantitative real-time PCR (qRT-PCR) were analysed for normal distribution with the Shapiro-Wilk test using assignment of a normal distribution with p>0.05 and confirmed using quantile-quantile plots. The means of indicated groups were compared using the two-tailed Student t test or one-way analysis of variance (ANOVA) with Tukey HSD post hoc test for studies with more than two groups. The results were considered statistically significant when p<0.05.
Results
Expression analysis of 5-HT receptors and the 5-HT transporter 5-HTT in donor and IPF lung specimens
Initially we analysed the expression of 5-HT receptors and the 5-HT transporter 5-HTT in lung tissue samples from controls (transplant donors) and from patients with NSIP or IPF using qRT-PCR. As shown in figure 1, all investigated 5-HT receptors except 5-HTR2C were expressed in normal as well as diseased lung tissue. 5-HTR1A, 5-HTR1B (figure 1A), 5-HTR2A and 5-HTR2B (figure 1B) were markedly upregulated in IPF lungs (mean±SEM log-fold change 2.36±0.43, 2.14±0.46, 3.74±0.41 and 1.60±0.32, respectively). 5-HTT was expressed at high levels in control lungs; it was dramatically downregulated in IPF (log-fold change −4.05±0.71, figure 1C) but not in NSIP lungs (fibrotic NSIP). Interestingly, in NSIP lung specimens 5-HTR2A and 5-HTT were not differentially regulated compared with control subjects but 5-HTR1A, 5-HTR1B and 5-HTR2B were also upregulated (figure 1). The increased expression of 5-HTR2A thus presented a specific event in UIP.

mRNA expression profile of serotonin (5-HT) receptors (A) 5-HTR1A,1B, (B) 5-HTR2A,2B and (C) the 5-HT transporter (5-HTT) in lung samples from controls (transplant donors) and patients with non-specific interstitial pneumonia (NSIP) and idiopathic pulmonary fibrosis/usual interstitial pneumonia (IPF/UIP) assessed by quantitative real-time PCR. The results are derived from 12 controls, 6 patients with NSIP and patients with 12 IPF/UIP and are presented as single values per lung sample. *p<0.05.
Localisation of 5-HTR2A and 5-HTR2B in donor and IPF lung specimens
Following the initial results, we focused our study on protein localisation of 5-HTR2A and 5-HTR2B. In order to localise cell types capable of 5-HT signalling via these 5-HTR, we assessed the distribution of 5-HTR2A and 5-HTR2B in donor and IPF lung tissue sections by immunohistochemistry (figures 2 and 3, respectively). 5-HTR2A was mainly expressed in bronchial and vascular smooth muscle cells; in IPF, additional staining in interstitial fibroblasts was observed (figure 2, arrows). As shown in figure 3 and figure S2 in the online supplement, 5-HTR2B was expressed largely in endothelial cells in control tissue. In IPF lungs, 5-HTR2B staining was mainly observed in bronchial and alveolar epithelial cells (figure 3A, arrowheads). To further corroborate these results we quantified the cell-specific gene expression patterns of 5-HTR2A and 5-HTR2B in primary human ATII cells derived from patients with IPF or transplant donors using qRT-PCR. As shown in figure 3B, there was a significant increase in 5-HTR2B mRNA expression in IPF ATII cells whereas the 5-HTR2A level was not significantly different in human ATII cells from patients with IPF and transplant donors.

Expression and localisation of the serotonin (5-HT) receptor 5-HTR2A in lung tissue from controls (transplant donors) and patients with idiopathic pulmonary fibrosis (IPF). Immunohistochemical staining was performed on tissue sections of donor lungs (left panels) and IPF lungs (right panels). Stainings are representative of four independent experiments using at least three different donor or IPF lung tissues. Scale bars indicate 100 μm. Arrows indicate positive staining of fibroblasts in IPF.

Expression and localisation of the serotonin (5-HT) receptor 5-HTR2B in lung tissue from controls (transplant donors) and patients with idiopathic pulmonary fibrosis (IPF). (A) Immunohistochemical stainings are representative of four independent experiments using at least three different donor or IPF lung tissues and two different primary antibodies. Scale bars indicate 100 μm. Arrows indicate positive staining of fibroblast in IPF, arrowheads highlight staining in bronchial and alveolar epithelial cells. (B) The mRNA levels of 5-HTR2A and 5-HTR2B were determined by qRT-PCR in primary human ATII cells (n=4) isolated from IPF lung tissue and compared with ATII cells from donor lungs. The results are plotted as fold change (2ΔΔCt) of mRNA levels in IPF-derived vs donor-derived cells and presented as mean±SEM, *p<0.05.
Administration of the 5-HTR2A/B antagonist terguride in vivo
We next assessed whether inhibition of 5-HTR2A/B represented an effective therapeutic option in lung fibrosis using mice subjected to orotracheal instillation of bleomycin, a well-accepted experimental model of lung fibrosis.31 First we analysed the expression of 5-HT receptors and 5-HTT in experimental lung fibrosis. As shown in figure S1 in the online supplement, 5-HTR2A and 5-HTR2B were significantly upregulated 7 and 14 days after bleomycin injury, which is in agreement with previous reports.23
Next we administered the clinically approved 5-HTR2A/B antagonist terguride to mice subjected to bleomycin. In order to ensure that terguride partitions into lung tissue, we next determined the concentrations of terguride in plasma and lung tissue under our experimental conditions (see figure S3 in online supplement). One hour after intraperitoneal administration of 1.2 mg/kg terguride the plasma concentrations of terguride ranged from 2.06–20.26 nM. In perfused lung tissue, comparable terguride concentrations to plasma were detected in all animals independent of the degree of fibrosis (figure S3 in online supplement). This indicated that terguride rapidly equilibrated in lung tissues and during the time course of bleomycin-induced lung fibrosis. Treatment with terguride was well tolerated and no side effects were observed throughout the study. The body weights of the mice were not significantly different between bleomycin-treated mice, independent of treatment with vehicle or terguride (see figure S3 in online supplement).
Effects of 5-HTR2A/B antagonism on lung fibrosis in vivo
To determine whether 5-HTR2A/B antagonism affected the progression of lung fibrosis we initiated treatment with terguride 14 days after bleomycin instillation (referred to as ‘therapeutic approach’). Mice were treated with two different doses of terguride by intraperitoneal application twice daily until day 28. As an internal control, a treatment arm with daily administration of 50 mg/kg imatinib was included in the study.32 Treatment with imatinib was started immediately after instillation of bleomycin (referred as ‘preventive approach’). The detailed treatment scheme is shown in figure S4 in the online supplement. We investigated the effects of 5-HTR2A/B antagonism by terguride in vivo on key features of lung fibrosis such as impaired respiratory function, destroyed lung architecture and increased collagen deposition.
The assessment of lung compliance in vivo showed slight but significant improvements in lung function of terguride-treated but not imatinib-treated mice (figure 4A). We next assessed the degree of lung fibrosis by determination of the total hydroxyproline content (figure 4B) and fibrotic score (figure 4C). As shown in figure 4B, mice subjected to 5-HTR2A/B antagonism by terguride had significantly less total collagen than vehicle-treated controls (bleomycin+vehicle: 148±22%, bleomycin+terguride 0.4: 122±27%, bleomycin+terguride 1.2: 91±21%, n=10 each). These findings were corroborated by the fact that 5-HTR2A/B antagonism also led to a dose-dependent decrease in the fibrotic score (bleomycin+vehicle: 3.1±0.3, bleomycin+terguride 0.4: 1.9±0.6, bleomycin+terguride 1.2: 1.46±0.2 compared with saline-treated controls, n=10 each; figure 4C). The effects of terguride on collagen content as well as the fibrotic score were comparable to those of imatinib treatment (preventive approach). Finally, immunohistochemistry of vehicle- and terguride-treated mouse lungs confirmed a marked attenuation of fibrosis with less ECM deposition and restored lung architecture (figure 4D).
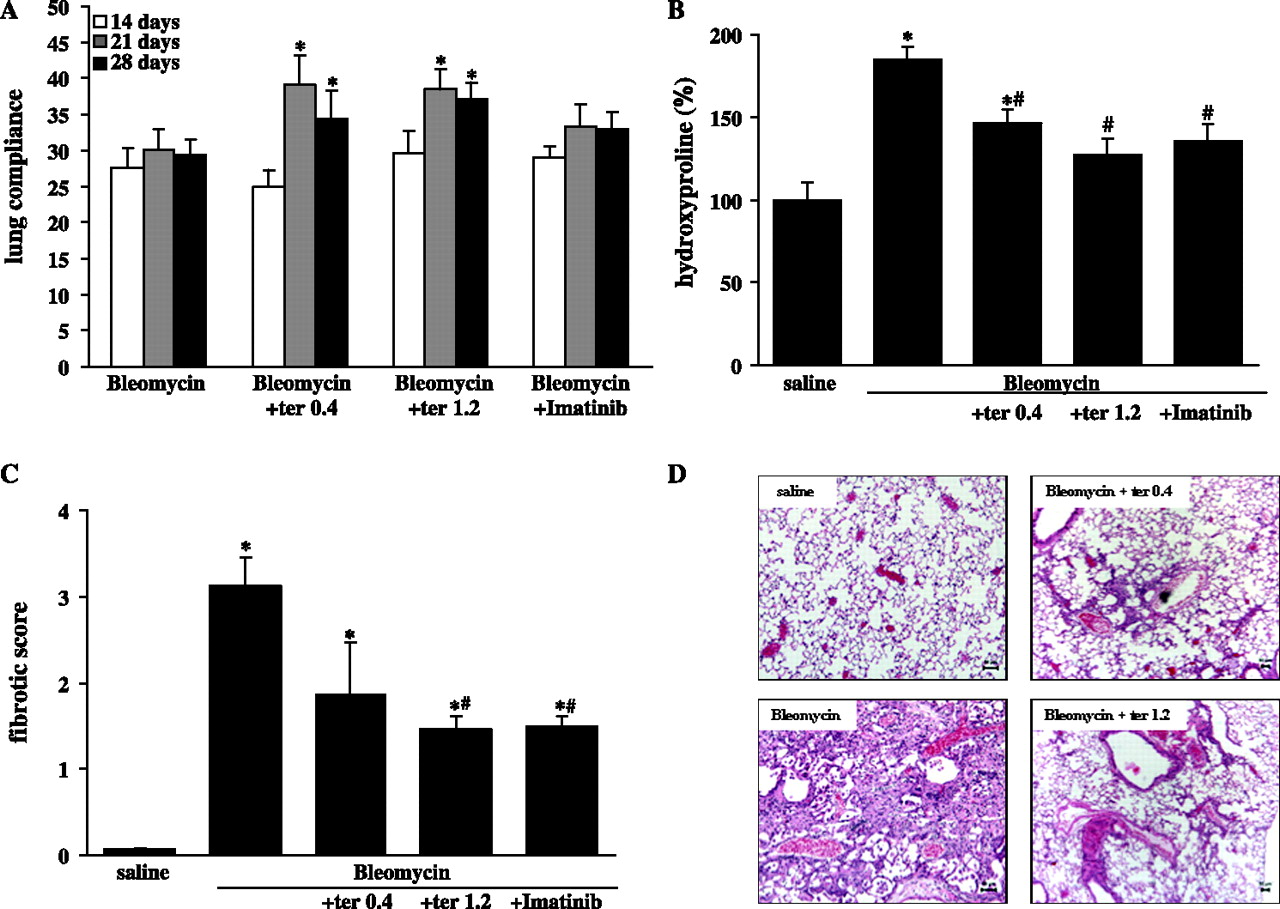
Lung function and collagen content after terguride treatment in experimental lung fibrosis. Mice were subjected to saline or bleomycin instillation and treated with terguride (ter). (A) Lung compliance measurements were obtained on days 14, 21 and 28. After 28 days the lungs were processed for (B) hydroxyproline measurement, (C) determination of the fibrotic score in lung sections stained with haematoxylin and eosin or (D) histological assessment using haematoxylin and eosin staining of lung sections. Scale bars indicate 50 μm.
Effects of 5-HTR2A/B antagonism on lung fibrosis in vitro
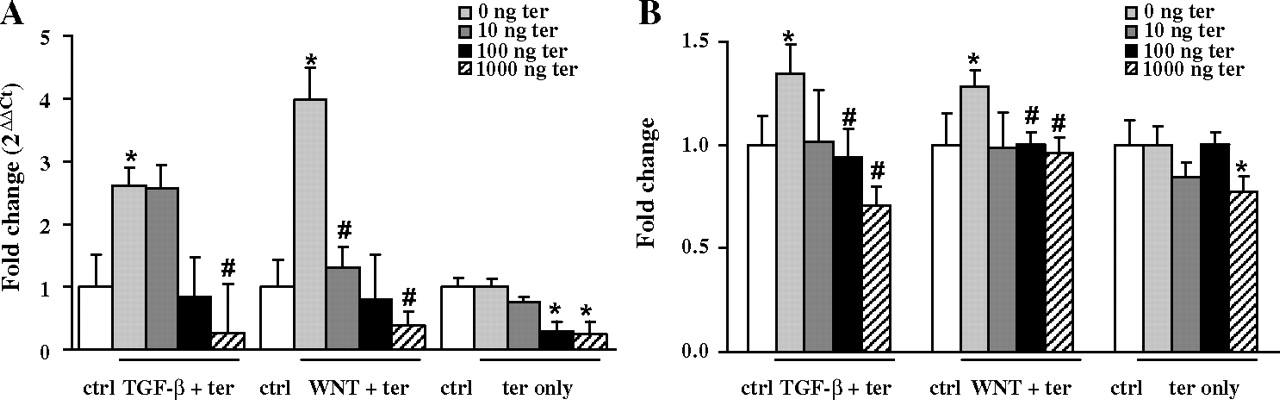
Fibroblasts are the key effector cells in lung fibrosis and are responsible for enhanced ECM deposition. We therefore assessed the effects of terguride on lung fibroblasts stimulated by the profibrotic mediators transforming growth factor (TGF)-β1 or WNT3a. Both, TGF-β1 and WNT3a treatment of human lung fibroblasts (HFL-1) led to a significant induction of the ECM components type I collagen α1 (Col1a1) as assessed by qRT-PCR (figure 5A). Moreover, these changes were confirmed at the total collagen level as assessed by Sircol assay (figure 5B). Most importantly, terguride treatment resulted in a significant decrease in TGF-β1-induced as well as WNT3a-induced collagen production at the mRNA (TGF-β1: 2.61±0.65, TGF-β1+terguride 1000: 0.26±0.77, WNT3a: 3.99±0.55, WNT3a+terguride 1000: 0.39±0.22; figure 5A) and protein level (TGF-β1: 1.34±0.29, TGF-β1+terguride 1000: 0.71±0.28, WNT3a: 1.28±0.16, WNT3a+terguride 1000: 0.96±0.12, figure 5B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of terguride (ter) treatment on transforming growth factor (TGF)‐β1- induced or WNT3a-induced type I collagen α1 expression in fibroblasts. (A) Using qRT-PCR analysis, the mRNA level of type I collagen α1 was assessed in human lung fibroblasts after stimulation with TGF-β1 (2 ng/ml) or WNT3a (100 ng/ml) for 12 h as indicated (n=4). (B) Human lung fibroblasts were stimulated with TGF-β1 (2 ng/ml) or WNT3a (100 ng/ml) for 24 h and the total collagen content was measured using the Sircol collagen assay (n=4).
Discussion
In this study we found altered expression of 5-HTR in IPF, suggesting that 5-HT signalling is altered in IPF. We analysed lung tissue samples from patients with IPF, NSIP and controls (transplant donors) and observed a marked upregulation of 5-HTR1A/B and 5-HTR2A/B but a dramatic downregulation of 5-HTT in IPF. Interestingly, in NSIP lung specimens 5-HTR2A was not differentially regulated compared with control subjects, which suggests this is a specific event in UIP.
We demonstrated that inhibition of 5-HTR2A and 5-HTR2B resulted in marked attenuation of bleomycin-induced lung fibrosis and thereby may be a suitable therapeutic approach in lung fibrosis. These findings are of special interest as IPF has a poor prognosis owing to limited responsiveness to currently available therapies.
5-HT and its receptors 5-HTR2A and 5-HTR2B have been implicated in fibrotic disorders such as retroperitoneal fibrosis, nephropathies, carcinoid heart disease and liver fibrosis.18–20 Cumulative evidence of the biological role of 5-HT in the lung has emerged, such as controlling vasoreactivity or bronchoreactivity.33 Under disease conditions such as asthma, increased levels of 5-HT correlate with the clinical status and pulmonary function,34 and several studies have indicated of the involvement of 5-HT in the aetiology of primary pulmonary hypertension.35 36 5-HT leads to vasoconstriction and vascular proliferation under hypoxic conditions,35 and enhanced 5-HTR2B expression seems to be centrally involved in the development of this disease.36
We quantified the mRNA expression levels of distinct 5-HT receptors and of 5-HTT in IPF, NSIP and control lung tissues. High levels of 5-HTR2A and 5-HTR2B were observed in the lung but 5-HTR2C was not detected. The expression of 5-HTR2A and 5-HTR2B was highly upregulated in IPF lung tissue and localised to key cell types involved in disease pathogenesis such as epithelial cells and fibroblasts. This is in line with previous findings that 5-HTR2A and 5-HTR2B are expressed in the human lung37 as well as in lung epithelial cell lines and fibroblasts.33 38 Most importantly, increased expression of 5-HTR2A was specific for IPF as no regulation of 5-HTR2A was observed in NSIP. In the NSIP lung specimen, classified as fibrotic NSIP, the highest expression level for 5-HTR1A and 5-HTR1B was observed. NSIP can be divided into ‘cellular’/‘non-fibrotic’ and ‘fibrotic’ phenotypes. In non-fibrotic NSIP mainly inflammatory cells are found whereas, in fibrotic NSIP, areas of thickened fibrotic septa with fewer inflammatory cells are predominant. Importantly, non-fibrotic NSIP has a significantly better prognosis and survival rate than fibrotic NSIP.39 40 Given the results of this study, future experiments may focus on 5-HTR1A and 5-HTR1B expression in non-fibrotic NSIP lung tissue specimens to elucidate whether this would be useful to distinguish subtypes of NSIP.
High levels of 5-HTT were found in the normal lung, with a dramatic downregulation in IPF. Decreased 5-HTT expression was also observed in bleomycin-induced lung fibrosis. 5-HTT is a key protein limiting systemic availability of 5-HT and is responsible for 5-HT uptake and subsequent inactivation of the amine passing through the lung. Mekontso-Dessap et al reported that mice deficient in 5-HTT develop cardiac fibrosis and valvulopathy, providing evidence for an inverse relationship between 5-HTT and fibrogenesis.41 Ablation of 5-HTT reduces the levels of 5-HT because platelet 5-HT is known to contribute 99% of 5-HT in the blood. Since 5-HTT is also the predominant pathway of 5-HT clearance by lung cells, an increase in the local 5-HT concentration in the lung with concomitant increases in binding to and signalling through 5-HT2 receptors may be the consequence.41 Downregulation of 5-HTT and upregulation of 5-HTR2A and 5-HTR2B may lead to increased 5-HT levels and enhanced receptor availability, respectively, and thus synergistic regulation of fibrogenesis. A detailed investigation of the underlying mechanisms remains elusive owing to limitations of investigative tools. In particular, the assessment of free and unbound 5-HT in nanomolar concentrations in blood is influenced by artificial activation of platelets and is therefore difficult to assess and interpret in mouse models.
We assessed the potential antifibrotic effects of 5-HTR2A/B antagonism using a therapeutic approach in the mouse model of bleomycin-induced lung fibrosis. We administered terguride, which is clinically approved for the treatment of ovulation disorders due to hyperprolactinaemia and hyperprolactaemic pituitary adenoma.27 Terguride belongs to the ergoline class of compounds which have been shown to have a high affinity to several 5-HT receptor isoforms. In contrast to most ergoline derivatives including ergotamine, pergoline, cabergoline and bromocriptine, terguride is a potent 5HTR2A and 5HTR2B receptor antagonist.24 Chronic treatment with ergoline derivatives such as pergolide and, to a lesser extent, cabergoline or with anorexic drugs such as fenfluramine has been associated with retroperitoneal, pleural and pericardial fibrosis as well as valvular heart disease.42 43 These drugs exhibit 5-HTR2B receptor agonist properties. In contrast, terguride acts as a non-surmountable antagonist on 5-HTR2B. Interestingly, Hauso et al recently showed that terguride treatment prevented 5-HT-induced heart valve disease and hypertrophy in rats.44
We have shown that administration of terguride at two different doses in a therapeutic approach had potent dose-dependent therapeutic effects in the mouse model of bleomycin-induced lung fibrosis. This model has been widely used for drug efficiency studies over the years, but most compounds have been tested in a preventive rather than a therapeutic approach. The preventive approach interferes mostly with the inflammatory and early fibrogenic response whereas the therapeutic approach acts on the progression of fibrosis, better reflecting the clinical situation in IPF.31 We showed that treatment with terguride in a therapeutic fashion led to a significant improvement in respiratory function, collagen deposition, lung architecture and survival. This suggests that terguride may have anti-remodelling properties. Previous evidence has indicated that 5-HT binding to 5-HTR2A exerts mitogenic properties via enhanced TGF-β1 expression. This mechanism seems to be regulated via protein kinase C and extracellular signal-regulated kinase.19 In addition, 5-HTR2B has been reported to interact with the platelet-derived growth factor receptor pathway to regulate cell cycle progression.45 Both receptors may provide possible pathways which lead to the reverse actions of 5-HTR2A/B antagonism by terguride. We have further corroborated the interaction between 5-HTR and TGF-β1 signalling, demonstrating that 5-HTR is involved in collagen production in human lung fibroblasts induced by TGF-β1.
Our findings are further validated by a recent study addressing the role of 5-HT signalling in experimental lung fibrosis. Fabre et al23 reported increased lung 5-HT levels and 5-HTR2A and 5-HTR2B expression during fibrogenesis in the bleomycin model of lung fibrosis, which we have confirmed in this study. Using a preventive approach and generic 5-HTR2A/B antagonists in the bleomycin model, they also demonstrated attenuation of lung fibrosis with the inhibitors ketanserin (a 5-HTR2A inhibitor) and SB215505 (a 5-HTR2B inhibitor).23 Our results are consistent with these findings, thus confirming the relevance of 5-HT signalling in lung fibrosis and its role as a therapeutic target. In addition, we have advanced the understanding of the role of 5-HT in the pathophysiology of IPF by (1) characterising and quantifying in detail the expression of 5-HT receptors in human IPF/UIP, NISP and control tissues; (2) demonstrating the therapeutic feasibility of terguride in the experimental model of lung fibrosis using therapeutically relevant conditions, thereby providing a rationale for translational research of a well-characterised and safe drug in pulmonary fibrosis; and (3) by investigating in detail the biological mechanisms involved in the therapeutic effect of terguride—for example, by demonstrating a significant role of 5-HTR2A/B in collagen turnover induced by the profibrotic mediators TGF-β1 and WNT3a.
In summary, we present strong evidence for an important role of 5-HT signalling via 5-HTR2A/B in lung fibrosis in vivo and in vitro, and the possibility of selectively and safely modulating this pathway in vivo which may provide a possible therapeutic approach for patients with IPF.
Acknowledgments
We are indebted to Simone Becker, Eva Bienek and Maria Magdalena Stein for excellent technical assistance, Walter Klepetko for providing lung tissues and all members of the Eickelberg Laboratory for stimulating discussions.
References
Supplementary materials
Web Only Data THORAX.2010.134353
Files in this Data Supplement:
Footnotes
Funding The authors are supported by the Helmholtz Association, the German Research Foundation (DFG) KliFo 118 and a career development award by the University of Giessen School of Medicine to MK.
Competing interests RR is a full-time employee of Ergonex Pharma.
Patient consent Obtained.
Ethics approval This study was conducted with the approval of the University of Giessen.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Editorial
- Airwaves